
Insularity and Aridity as Drivers of Mandibular Disparity in Thylamys elegans (Waterhouse, 1839) from Populations of the Atacama Desert, Chile

 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
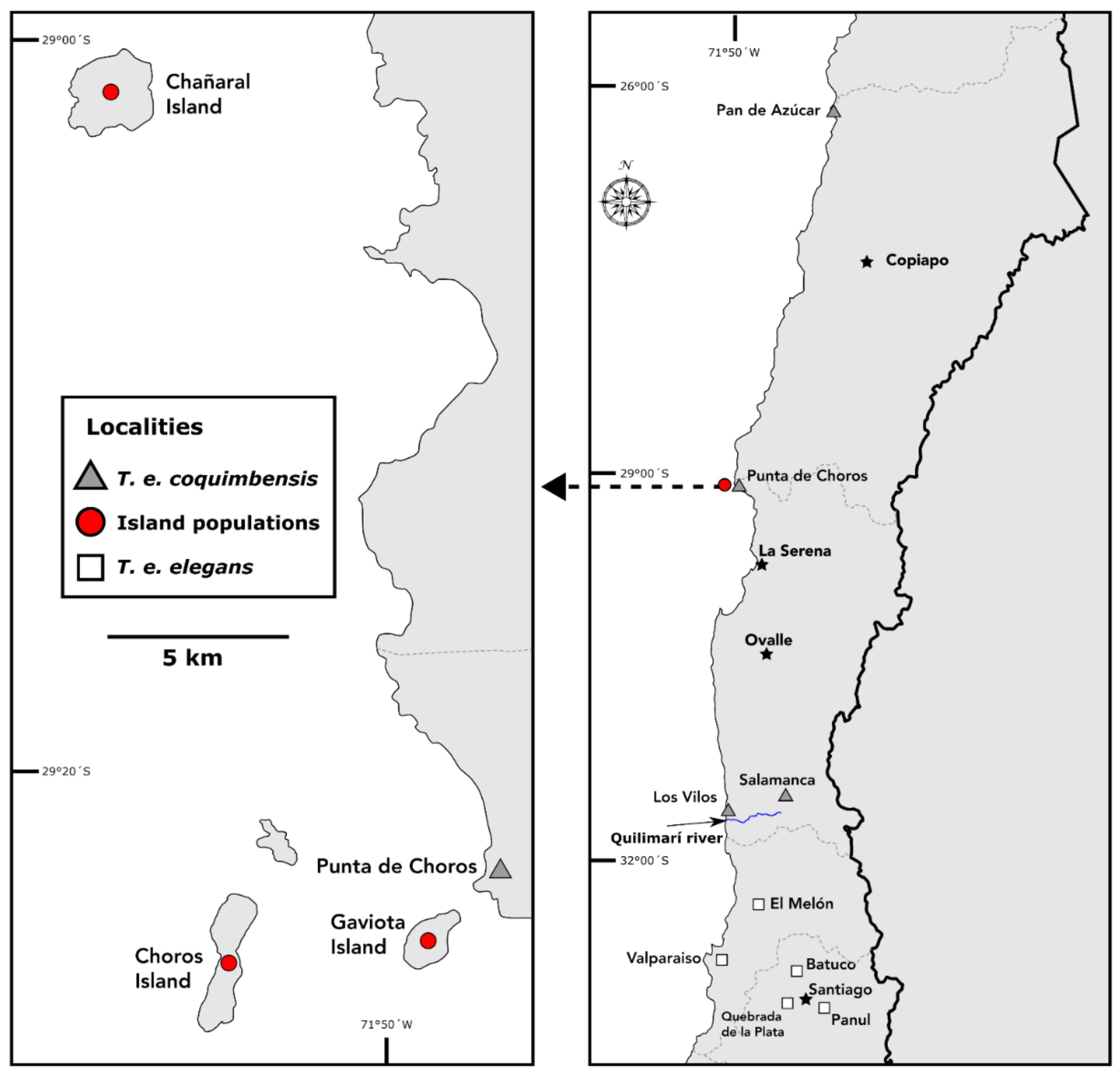
2. Materials and Methods
2.1. Thylamys elegans Subspecies
2.2. Linear Morphometrics
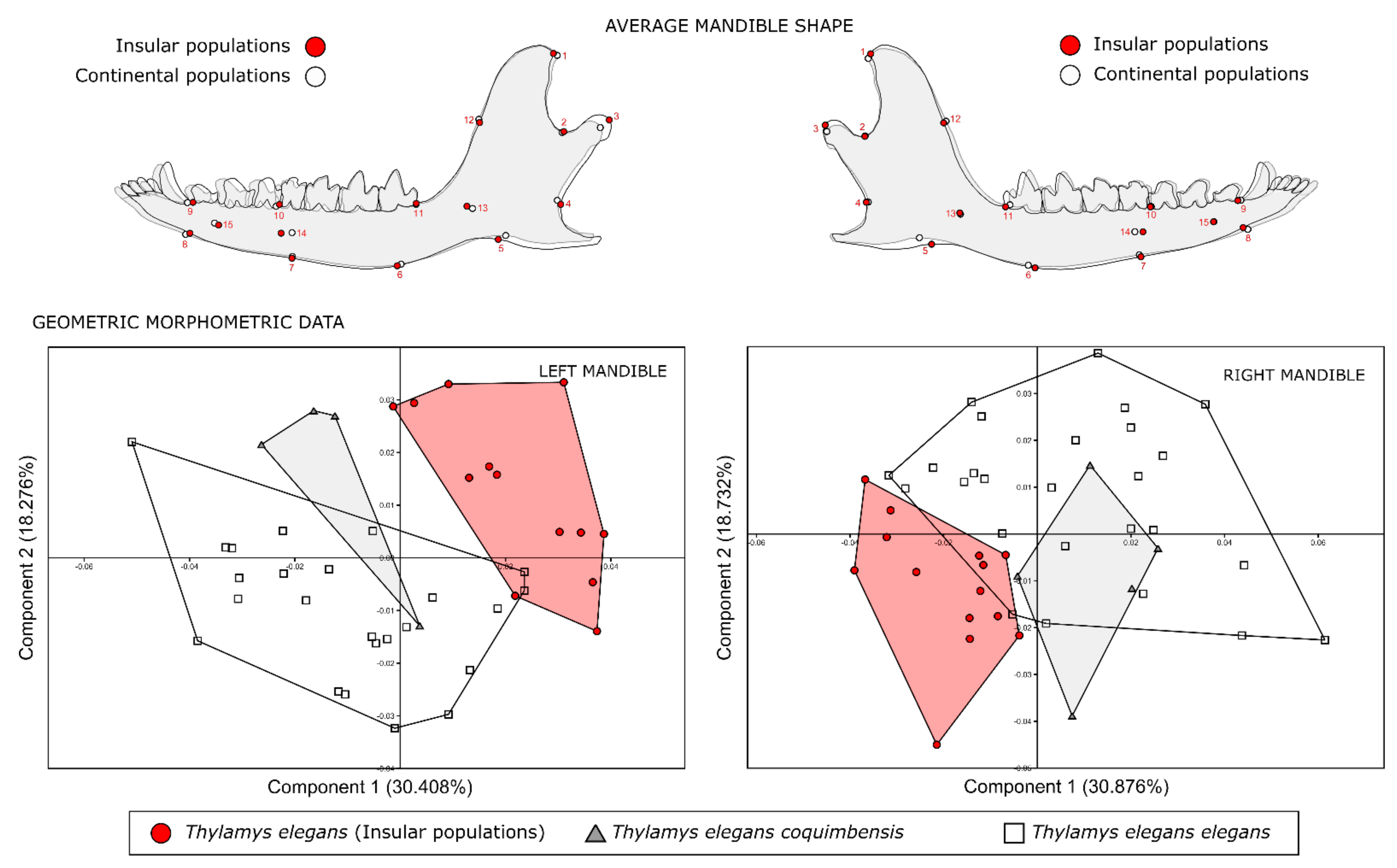
2.3. Geometric Morphometrics
2.4. Biomechanical Model
2.5. Comparisons between Groups
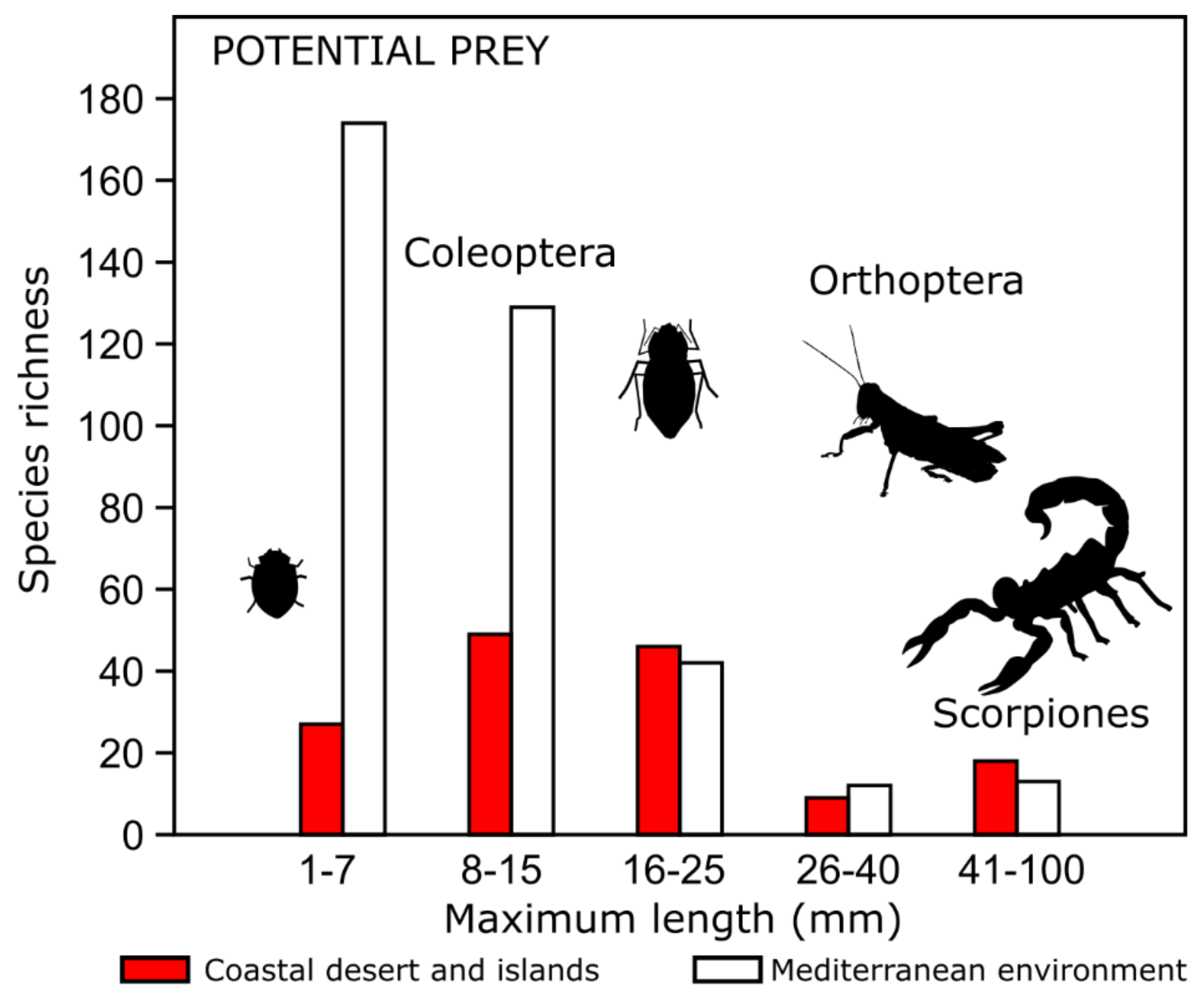
2.6. Characterization of Potential Prey: Arthropods, Fruits and Seeds
2.7. Relation between Environment and Mandible Shape and Size
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Grant, P.R.; Grant, B.R. Unpredictable evolution in a 30-year study of Darwin’s finches. Science 2002, 296, 707–711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Losos, J.B.; Glor, R.E.; Kolbe, J.J.; Nicholson, K. Adaptation, speciation, and convergence: A hierarchical analysis of adaptive radiation in Caribbean Anolis lizards1. Ann. Mo. Bot. Gard. 2006, 93, 24–33. [Google Scholar] [CrossRef] [Green Version]
- Monteiro, L.R.; Nogueira, M.R. Adaptive radiations, ecological specialization, and the evolutionary integration of complex morphological structures. Evolution 2010, 64, 724–743. [Google Scholar] [CrossRef] [PubMed]
- Berry, R. Small mammal differentiation on islands. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 1996, 351, 753–764. [Google Scholar]
- Millien, V. Morphological evolution is accelerated among island mammals. PLoS Biol. 2006, 4, e321. [Google Scholar] [CrossRef]
- Whittaker, R.J.; Fernández-Palacios, J.M.; Matthews, T.J.; Borregaard, M.K.; Triantis, K.A. Island biogeography: Taking the long view of nature’s laboratories. Science 2017, 357, eaam8326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foster, J.B. Evolution of mammals on islands. Nature 1964, 202, 234–235. [Google Scholar] [CrossRef]
- Lomolino, M.V. Body size of mammals on islands: The island rule reexamined. Am. Nat. 1985, 125, 310–316. [Google Scholar] [CrossRef]
- Lomolino, M.V. Body size evolution in insular vertebrates: Generality of the island rule. J. Biogeogr. 2005, 32, 1683–1699. [Google Scholar] [CrossRef]
- Valladares-Gómez, A.; Huenumilla-Linares, M.; Rodríguez-Serrano, E.; Hernández, C.E.; Palma, R.E. Morphological variation in two sigmodontine rodents along the mainland and the Fuegian archipelago in Chilean southern Patagonia. Rev. Chil. Hist. Nat. 2020, 93. [Google Scholar] [CrossRef]
- Ponce, J.F.; Rabassa, J.; Coronato, A.; Borromei, A.M. Palaeogeographical evolution of the Atlantic coast of Pampa and Patagonia from the last glacial maximum to the Middle Holocene. Biol. J. Linn. Soc. 2011, 103, 363–379. [Google Scholar] [CrossRef] [Green Version]
- Davies, B.J.; Darvill, C.M.; Lovell, H.; Bendle, J.M.; Dowdeswell, J.A.; Fabel, D.; García, J.-L.; Geiger, A.; Glasser, N.F.; Gheorghiu, D.M. The evolution of the Patagonian Ice Sheet from 35 ka to the present day (PATICE). Earth-Sci. Rev. 2020, 204, 103152. [Google Scholar] [CrossRef]
- Lambeck, K.; Rouby, H.; Purcell, A.; Sun, Y.; Sambridge, M. Sea level and global ice volumes from the Last Glacial Maximum to the Holocene. Proc. Natl. Acad. Sci. USA 2014, 111, 15296–15303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finstad, K.M.; Pfeiffer, M.; McNicol, G.; Tuite, M.; Williford, K.; Amundson, R. A late Quaternary paleoenvironmental record in sand dunes of the northern Atacama Desert, Chile. Quat. Res. 2018, 90, 127–138. [Google Scholar] [CrossRef]
- Cornette, R.; Herrel, A.; Cosson, J.-F.; Poitevin, F.; Baylac, M. Rapid morpho-functional changes among insular populations of the greater white-toothed shrew. Biol. J. Linn. Soc. 2012, 107, 322–331. [Google Scholar] [CrossRef] [Green Version]
- Pergams, O.R.; Byrn, D.; Lee, K.L.; Jackson, R. Rapid morphological change in black rats (Rattus rattus) after an island introduction. PeerJ 2015, 3, e812. [Google Scholar] [CrossRef] [Green Version]
- Vega, R.; Mcdevitt, A.D.; Kryštufek, B.; Searle, J.B. Ecogeographical patterns of morphological variation in pygmy shrews Sorex minutus (Soricomorpha: Soricinae) within a phylogeographical and continental-and-island framework. Biol. J. Linn. Soc. 2016, 119, 799–815. [Google Scholar] [CrossRef] [Green Version]
- Fabre, P.H.; Herrel, A.; Fitriana, Y.; Meslin, L.; Hautier, L. Masticatory muscle architecture in a water-rat from Australasia (Murinae, Hydromys) and its implication for the evolution of carnivory in rodents. J. Anat. 2017, 231, 380–397. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Chardi, A.; García-Pando, M.; Pujol-Buxó, E.; Sans-Fuentes, M.A.; López-Fuster, M.J.; Muñoz-Muñoz, F. Insularity induces changes on body and mandible morphology in a Mediterranean population of the greater white-toothed shrew Crocidura russula (Hermann, 1780). Contrib. Zool. 2018, 87, 275–286. [Google Scholar] [CrossRef] [Green Version]
- Benítez, H.A.; Pizarro-Araya, J.; Bravi, R.; Sanzana, M.-J.; Alfaro, F.M. Morphological variation on isolated populations of Praocis (Praocis) spinolai. J. Insect Sci. 2014, 14, 11. [Google Scholar] [CrossRef]
- Gardner, A.L. Mammals of South America, Volume 1: Marsupials, Xenarthrans, Shrews, and Bats; University of Chicago Press: Chicago, IL, USA, 2008. [Google Scholar]
- Voss, R.S.; Jansa, S.A. Opossums: An Adaptive Radiation of New World Marsupials; Johns Hopkins University Press: Baltimore, MD, USA, 2021. [Google Scholar]
- Voss, R.S.; Jansa, S.A. Phylogenetic relationships and classification of didelphid marsupials, an extant radiation of New World metatherian mammals. Bull. Am. Mus. Nat. Hist. 2009, 2009, 1–177. [Google Scholar] [CrossRef]
- Palma, R.E.; Boric-Bargetto, D.; Jayat, J.P.; Flores, D.A.; Zeballos, H.; Pacheco, V.; Cancino, R.A.; Alfaro, F.D.; Rodríguez-Serrano, E.; Pardiñas, U.F. Molecular phylogenetics of mouse opossums: New findings on the phylogeny of Thylamys (Didelphimorphia, Didelphidae). Zool. Scr. 2014, 43, 217–234. [Google Scholar] [CrossRef]
- Pedreros, A.H.M.; Valenzuela, J.Y. Mamíferos de Chile; CEA Ediciones: Santiago, Chile, 2009. [Google Scholar]
- Sabat, P.; Bozinovic, F.; Zambrano, F. Role of dietary substrates on intestinal disaccharidases, digestibility, and energetics in the insectivorous mouse-opossum (Thylamys elegans). J. Mammal. 1995, 76, 603–611. [Google Scholar] [CrossRef]
- Mann Fischer, G. Los Pequeños Mamíferos de Chile: Marsupiales, Quirópteros, Edentados y Roedores; Gayana n°40 Universidad de Concepción: Concepción, Chile, 1978. [Google Scholar]
- Mella, J.; Simonetti, J.A.; Spotorno, A.; Contreras, L. Mamíferos de Chile. In Diversidad y conservación de los Mamíferos Neotropicales; Conabio-UNAM: Mexico City, Mexico, 2002. [Google Scholar]
- Boric-Bargetto, D.; Zuniga-Reinoso, A.; Cancino, R.A.; Gonzalez-Acuna, D.; Rodriguez-Serrano, E.; Palma, R.E.; Hernández, C.E. Phylogenetic evaluation of taxonomic definition of didelphid mouse opossum of the genus Thylamys from valleys of Coquimbo region, Chile. Zootaxa 2016, 4105, 339–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boric-Bargetto, D.; Zúñiga-Reinoso, Á.; Inostroza-Michel, O.; Rodríguez-Serrano, E.; González-Acuña, D.; Palma, R.E.; Hernández, C.E. A comprehensive overview of the genetic diversity in Thylamys elegans (Didelphimorphia: Didelphidae): Establishing the phylogeographic determinants. Revista Chilena de Historia Natural 2021, 94, 5. [Google Scholar] [CrossRef]
- Luna-Jorquera, G.; Fernández, C.E.; Rivadeneira, M.M. Determinants of the diversity of plants, birds and mammals of coastal islands of the Humboldt current systems: Implications for conservation. Biodivers. Conserv. 2012, 21, 13–32. [Google Scholar] [CrossRef]
- Cruz-Jofré, F.; Vilina, Y.A. Ecología trófica de Athene cunicularia (Aves: Strigidae) en un sistema insular del norte de Chile:¿ posible respuesta funcional y numérica frente a Pelecanoides garnotii (Aves: Pelecanoididae)? Gayana (Concepción) 2014, 78, 31–40. [Google Scholar] [CrossRef] [Green Version]
- Gajardo, R. La Vegetación Natural de Chile: Clasificación y Distribución Geográfica; Editorial Universitaria: Santiago, Chile, 1994. [Google Scholar]
- Arancio, G.; Jara, P. Flora de la Reserva Nacional Pingüino de Humboldt; Editorial Universidad de La Serena: La Serena, Chile, 2007. [Google Scholar]
- Arancio, G.; Munoz, M.; Squeo, F. Descripción de algunas especies con problemas de conservación en la IV Región de Coquimbo. In Libro Rojo de la Flora Nativa de la Región de Coquimbo y de los Sitios Prioritarios para su Conservación; Squeo, F.A., Arancio, G., Gutiérrez, J.R., Eds.; Ediciones de la Universidad de La Serena: La Serena, Chile, 2001. [Google Scholar]
- Aguilella, A.; Amigo, J.; Mota-Poveda, J.; Gómez-Mercado, T. Transectos de vegetación en el centro-sur de Chile. In Vegetación y Cambios Climáticos; University of Almería: Almería, Spain, 2001; pp. 87–101. [Google Scholar]
- Prevosti, F.J.; Turazzini, G.F.; Ercoli, M.D.; Hingst-Zaher, E. Mandible shape in marsupial and placental carnivorous mammals: A morphological comparative study using geometric morphometrics. Zool. J. Linn. Soc. 2012, 164, 836–855. [Google Scholar] [CrossRef] [Green Version]
- Echarri, S.; Ercoli, M.D.; Chemisquy, M.A.; Turazzini, G.; Prevosti, F.J. Mandible morphology and diet of the South American extinct metatherian predators (Mammalia, Metatheria, Sparassodonta). Earth Environ. Sci. Trans. R. Soc. Edinb. 2015, 106, 277–288. [Google Scholar] [CrossRef] [Green Version]
- Lemos, B.; Cerqueira, R. Morphological differentiation in the white-eared opossum group (Didelphidae: Didelphis). J. Mammal. 2002, 83, 354–369. [Google Scholar] [CrossRef] [Green Version]
- Fonseca, R.; Astúa, D. Geographic variation in Caluromys derbianus and Caluromys lanatus (Didelphimorphia: Didelphidae). Zoologia 2015, 32, 109–122. [Google Scholar] [CrossRef] [Green Version]
- Damasceno, E.M.; Astúa, D. Geographic variation in cranial morphology of the Water Opossum Chironectes minimus (Didelphimorphia, Didelphidae). Mamm. Biol. 2016, 81, 380–392. [Google Scholar] [CrossRef] [Green Version]
- Michaux, J.; Chevret, P.; Renaud, S. Morphological diversity of Old World rats and mice (Rodentia, Muridae) mandible in relation with phylogeny and adaptation. J. Zool. Syst. Evol. Res. 2007, 45, 263–279. [Google Scholar] [CrossRef]
- De Esteban-Trivigno, S. Ecomorphology of extinct Xenarthrans: Analysis of the mandible using geometric morphometrics methods. Ameghiniana 2011, 48, 381–398. [Google Scholar] [CrossRef]
- Meloro, C.; O’Higgins, P. Ecological Adaptations of Mandibular Form in Fissiped Carnivora. J. Mamm. Evol. 2011, 18, 185–200. [Google Scholar] [CrossRef]
- Vieira, E.M.; De Moraes, D.A. Carnivory and insectivory in Neotropical marsupials. In Predators with Pouches: The Biology of Carnivorous Marsupials; CSIRO Publishing: Clayton, Australia, 2003; pp. 271–284. [Google Scholar]
- De Moraes, D.A.; Hingst-Zaher, E.; Marcus, L.; Cerqueira, R. A geometric morphometric analysis of cranial and mandibular shape variation of didelphid marsupials. Hystrix Ital. J. Mammal. 2000, 11. [Google Scholar]
- Astúa, D. Cranial sexual dimorphism in New World marsupials and a test of Rensch’s rule in Didelphidae. J. Mammal. 2010, 91, 1011–1024. [Google Scholar] [CrossRef]
- Cerqueira, R.; Lemos, B. Morphometric differentiation between Neotropical black-eared opossums, Didelphis marsupialis and D. aurita (Didelphimorphia, Didelphidae). Mammalia 2000, 64, 319–327. [Google Scholar] [CrossRef]
- Martínez-Lanfranco, J.A.; Flores, D.; Jayat, J.P.; D’Elía, G. A new species of lutrine opossum, genus Lutreolina Thomas (Didelphidae), from the South American Yungas. J. Mammal. 2014, 95, 225–240. [Google Scholar] [CrossRef] [Green Version]
- Rohlf, F.J. TPSdig, version 2.17; Stony Brook University: New York, NY, USA, 2013. [Google Scholar]
- Rohlf, F.J.; Slice, D. Extensions of the Procustes methods for the optimal superimposition of landmarks. Syst. Zool. 1990, 39, 40–59. [Google Scholar] [CrossRef] [Green Version]
- Young, R.L.; Haselkorn, T.S.; Badyaev, A.V. Functional equivalence of morphologies enables morphological and ecological diversity. Evol. Int. J. Org. Evol. 2007, 61, 2480–2492. [Google Scholar] [CrossRef]
- Jolliffe, I.T. Principal Component Analysis, 2nd ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Benítez, H.A.; Püschel, T.A. Modelando la Varianza de la Forma: Morfometría Geométrica Aplicaciones en Biología Evolutiva. Int. J. Morphol. 2014, 32, 998–1008. [Google Scholar] [CrossRef] [Green Version]
- Klingenberg, C.P. MorphoJ: An integrated software package for geometric morphometrics. Mol. Ecol. Resour. 2011, 11, 353–357. [Google Scholar] [CrossRef] [PubMed]
- Villalobos-Leiva, A.; Benítez, H.A. Morfometría Geométrica y sus Nuevas Aplicaciones en Ecología y Biología Evolutiva. Parte 2. Int. J. Morphol. 2020, 38, 1818–1836. [Google Scholar] [CrossRef]
- Monteiro, L.R. Multivariate regression models and geometric morphometrics: The search for causal factors in the analysis of shape. Syst. Biol. 1999, 48, 192–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontologia Electronica 2001, 4, 9. [Google Scholar]
- Yu, S.; Katz, O.; Fang, W.; Li, D.; Sang, W.; Liu, C. Shift of fleshy fruited species along elevation: Temperature, canopy coverage, phylogeny and origin. Sci. Rep. 2017, 7, 40417. [Google Scholar] [CrossRef] [Green Version]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Palma, R.E. Thylamys elegans. Mamm. Species 1997, 572, 1–4. [Google Scholar] [CrossRef]
- MacArthur, R.H.; Wilson, E.O. An equilibrium theory of insular zoogeography. Evolution 1963, 17, 373–387. [Google Scholar] [CrossRef]
- Silva, S.I. Posiciones tróficas de pequeños mamíferos en Chile: Una revisión. Revista Chilena de Historia Natural 2005, 78, 589–599. [Google Scholar] [CrossRef] [Green Version]
- Bozinovic, F.; Méndez, M.A. Role of dietary fatty acids on energetics and torpor in the Chilean mouse-opossum Thylamys elegans. Comp. Biochem. Physiol. Part A Physiol. 1997, 116, 101–104. [Google Scholar] [CrossRef]
- Hauenstein, E.; Muñoz-Pedreros, A.; Yánez, J.; Sánchez, P.; Möller, P.; Guiñez, B.; Gil, C. Flora y vegetación de la Reserva Nacional Lago Peñuelas, Reserva de la Biósfera, Región de Valparaíso, Chile. Bosque 2009, 30, 159–179. [Google Scholar] [CrossRef] [Green Version]
- Jaksic, F.M. Spatiotemporal variation patterns of plants and animals in San Carlos de Apoquindo, central Chile. Rev. Chil. Hist. Nat. 2001, 74, 477–502. [Google Scholar] [CrossRef] [Green Version]
- Pizarro-Araya, J.; Ojanguren-Affilastro, A.A.; López-Cortés, F.; Agusto, P.; Briones, R.; Cepeda-Pizarro, J. Diversidad y composición estacional de la escorpiofauna (Arachnida: Scorpiones) del archipiélago Los Choros (Región de Coquimbo, Chile). Gayana (Concepción) 2014, 78, 46–56. [Google Scholar] [CrossRef] [Green Version]
- Valdivia, D.E.; Pizarro-Araya, J.; Cepeda-Pizarro, J.; Ojanguren-Affilastro, A.A. Diversidad taxonómica y denso-actividad de solífugos (Arachnida: Solifugae) asociados a un ecosistema desértico costero del centro norte de Chile. Revista de la Sociedad Entomológica Argentina 2008, 67, 1–10. [Google Scholar]
- Valdivia, D.E.; Pizarro-Araya, J.; Briones, R.; Ojanguren-Affilastro, A.A.; Cepeda-Pizarro, J. Species composition and abundance of solpugids (Arachnida: Solifugae) in ecotopes of the transitional coastal desert of Chile. Revista Mexicana de Biodiversidad 2011, 82, 1234–1242. [Google Scholar] [CrossRef] [Green Version]
- Flores, G.E.; Pizarro-Araya, J. Systematic revision of the South American genus Praocis Eschscholtz, 1829 (Coleoptera: Tenebrionidae). Part 1: Introduction and subgenus Praocis s. str. Zootaxa 2012, 3336, 1–35. [Google Scholar] [CrossRef]
- Alfaro, F.M.; Pizarro-Araya, J.; Flores, G.E. Epigean tenebrionids (Coleoptera: Tenebrionidae) from the Choros Archipelago (Coquimbo Region, Chile). Entomol. News 2009, 120, 125–130. [Google Scholar] [CrossRef]
- Alfaro, F.M.; Pizarro-Araya, J.; Flores, G.E. Composición y estructura del ensamble de tenebriónidos epigeos (Coleoptera: Tenebrionidae) de ecosistemas continentales e insulares del desierto costero transicional de Chile. Revista Mexicana de Biodiversidad 2016, 87, 1283–1291. [Google Scholar] [CrossRef] [Green Version]
- Alfaro, F.M.; Pizarro-Araya, J.; Letelier, L.; Cepeda-Pizarro, J. Distribución geográfica de los ortópteros (Insecta: Orthoptera) presentes en las provincias biogeográficas de Atacama y Coquimbo (Chile). In Revista de Geografía Norte Grande; 2013; Volume 56, pp. 235–250. [Google Scholar]
- Morales-García, N.M.; Gill, P.G.; Janis, C.M.; Rayfield, E.J. Jaw shape and mechanical advantage are indicative of diet in Mesozoic mammals. Commun. Biol. 2021, 4, 242. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, Y.; Lambeck, K.; De Deckker, P.; Johnston, P.; Fifield, L.K. Timing of the Last Glacial Maximum from observed sea-level minima. Nature 2000, 406, 713–716. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Coastal Desert | Mediterranean Region | |||
|---|---|---|---|---|
| Type of Fruit | No. Species | % | No. Species | % |
| Fleshy | 4 | 10 | 29 | 31 |
| Not Fleshy | 36 | 90 | 67 | 69 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arriagada, J.I.; Benítez, H.A.; Toro, F.; Suazo, M.J.; Abarca, P.; Canto, J.; Vilina, Y.A.; Cruz-Jofré, F. Insularity and Aridity as Drivers of Mandibular Disparity in Thylamys elegans (Waterhouse, 1839) from Populations of the Atacama Desert, Chile. Animals 2022, 12, 1179. https://doi.org/10.3390/ani12091179
Arriagada JI, Benítez HA, Toro F, Suazo MJ, Abarca P, Canto J, Vilina YA, Cruz-Jofré F. Insularity and Aridity as Drivers of Mandibular Disparity in Thylamys elegans (Waterhouse, 1839) from Populations of the Atacama Desert, Chile. Animals. 2022; 12(9):1179. https://doi.org/10.3390/ani12091179
Chicago/Turabian StyleArriagada, José I., Hugo A. Benítez, Frederick Toro, Manuel J. Suazo, Paulette Abarca, Jhoann Canto, Yerko A. Vilina, and Franco Cruz-Jofré. 2022. "Insularity and Aridity as Drivers of Mandibular Disparity in Thylamys elegans (Waterhouse, 1839) from Populations of the Atacama Desert, Chile" Animals 12, no. 9: 1179. https://doi.org/10.3390/ani12091179