
Morphological Stasis in Time? A Triatoma brasiliensis brasiliensis Study Using Geometric Morphometrics in the Long Run

 , , ,
, , ,  , and
, and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Origin and Identification of T. b. brasiliensis Specimens
2.2. Wings Preparation
2.3. Data Collection
2.4. Geometric Morphometrics Analyses
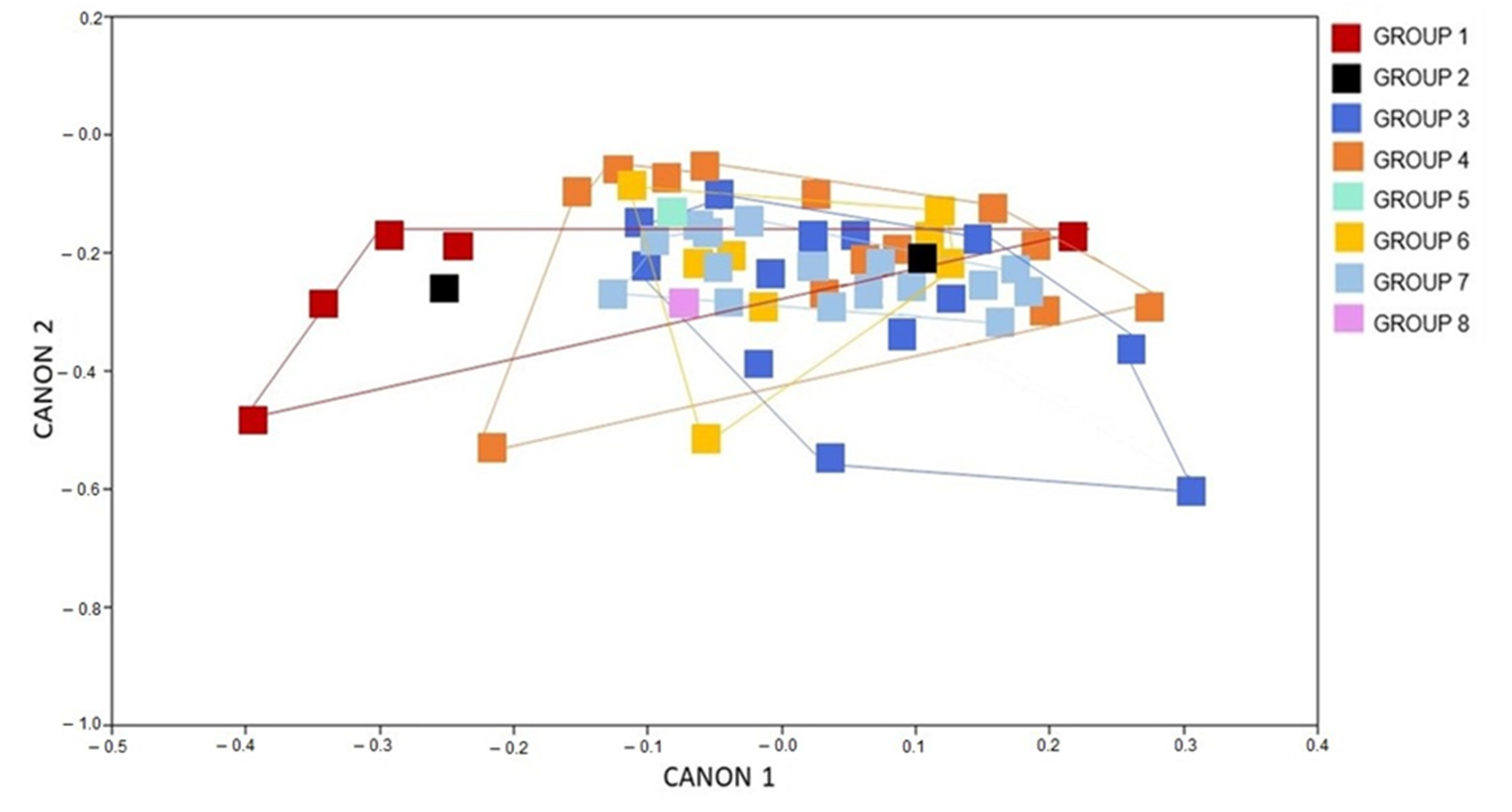
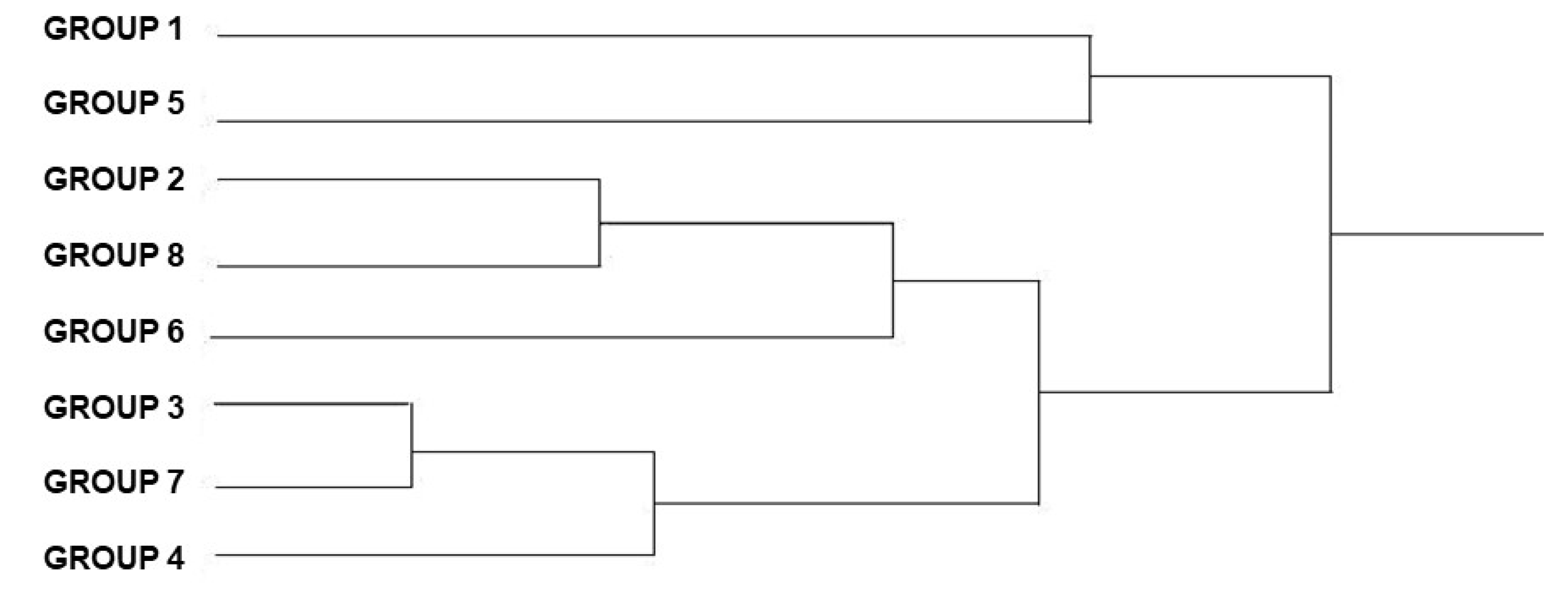
3. Results
3.1. Morphometric Analysis
3.1.1. Analysis of the Females Wings
3.1.2. Analysis of the Males Wings
3.1.3. Analysis of the Males and Females Wing Shape
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Silveira, A.C. Os novos desafios e perspectivas futuras do controle. Rev. Soc. Bras. Med. Trop. 2011, 44, 122–124. (In Portuguese) [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanaway, J.D.; Roth, G. The burden of Chagas disease: Estimates and challenges. Glob. Heart 2015, 10, 139–144. [Google Scholar] [CrossRef] [PubMed]
- WHO. World Health Organization. Chagas Disease (American Trypanosomiasis). 2021. Available online: https://www.who.int/news-room/facts-in-pictures/detail/chagas-disease (accessed on 11 January 2022).
- Costa, J.; Dale, C.; Galvão, C.; Almeida, C.E.; Dujardin, J.P. Do the new triatomine species pose new challenges or strategies for monitoring Chagas disease? An overview from 1979–2021. Mem. Inst. Oswaldo Cruz 2021, 116, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Alevi, K.C.C.; de Oliveira, J.; da Silva Rocha, D.; Galvão, C. Trends in Taxonomy of Chagas Disease Vectors (Hemiptera, Reduviidae, Triatominae): From Linnaean to Integrative Taxonomy. Pathogens 2021, 10, 1627. [Google Scholar] [CrossRef]
- Costa, J.; Correia, N.C.; Lima-Neiva, V.; Gonçalves, T.C.M.; Felix, M. Revalidation and redescription of Triatoma brasiliensis macromelasoma Galvão, 1956 and an identification key for the Triatoma brasiliensis complex (Hemiptera: Reduviidae: Triatominae). Mem. Inst. Oswaldo Cruz 2013, 108, 785–789. [Google Scholar] [CrossRef]
- Costa, J.; Dornak, L.L.; Almeida, C.E.; Peterson, A.T. Distributional potential of the Triatoma brasiliensis species complex at present and under scenarios of future climate conditions. Parasit Vectors 2014, 7, 238. [Google Scholar] [CrossRef]
- Costa, J.; Lima-Neiva, V.; Almeida, C.E.O. complexo Triatoma brasiliensis (Hemiptera, Reduviidae, Triatominae) como modelo de estudo: Uma abordagem multidisciplinar e ecoepidemiológica. Atual. Med. Trop. Vetores 2020, 6, 99–121. (In Portuguese) [Google Scholar] [CrossRef]
- Dale, C.; Almeida, C.E.; Mendonça, V.J.; Oliveira, J.; da Rosa, J.A.; Galvão, C.; Costa, J. An updated and illustrated dichotomous key for the Chagas disease vectors of Triatoma brasiliensis species complex and their epidemiologic importance. ZooKeys 2018, 805, 33–43. [Google Scholar] [CrossRef]
- Nascimento, D.; Novais, G. Clima do Cerrado: Dinâmica atmosférica e características, variabilidades e tipologias climáticas. Élisée Rev. Geografia UEG 2020, 9, e922021. (In Portuguese) [Google Scholar]
- Moreira, J.N. Caracterização da vegetação de Caatinga e da dieta de novilhos no Sertão de Pernambuco. Pesqui Agropecu 2006, 41, 1643–1651. (In Portuguese) [Google Scholar] [CrossRef] [Green Version]
- Costa, J.; Almeida, C.E.; Dotson, E.M.; Lins, A.; Vinhaes, M.; Silveira, A.C.; Beard, C.B. The epidemiologic importance of Triatoma brasiliensis as a Chagas disease vector in Brazil: A revision of domiciliary captures during 1993–1999. Mem. Inst. Oswaldo Cruz 2003, 98, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Silveira, A.C. Situação do controle da transmissão vetorial da doença de Chagas nas Américas. Cad. Saúde Pública 2000, 16, 35–42. (In Portuguese) [Google Scholar] [CrossRef] [PubMed]
- Almeida, C.E.; Faucher, L.; Lavina, M.; Costa, J.; Harry, M. Molecular Individual-Based Approach on Triatoma brasiliensis: Inferences on Triatomine Foci, Trypanosoma cruzi Natural Infection Prevalence, Parasite Diversity and Feeding Sources. PLoS Negl. Trop. Dis. 2016, 10, e0004447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lilioso, M.; Folly-Ramos, E.; Rocha, F.L.; Rabinovich, J.; Capdevielle-Dulac, C.; Harry, M. High Triatoma brasiliensis densities and Trypanosoma cruzi prevalence in domestic and peridomestic habitats in the state of Rio Grande do Norte, Brazil: The source for Chagas disease outbreaks? J. Trop. Med. Hyg. 2017, 96, 1456–1459. [Google Scholar] [CrossRef] [PubMed]
- Bezerra, C.M.; Belisário, C.J.; Pessoa, G.C.D.; Barezani, C.P.; Rosa, A.C.L.; Ferreira, F.C.; Ramos, A.N., Jr.; Gurtler, R.E.; Diotaiuti, L. Microsatellite variation revealed panmictic pattern for Triatoma brasiliensis (Triatominae: Reduviidae) in rural northeastern Brazil: The control measures implications. BMC Genom. 2020, 21, 92. [Google Scholar] [CrossRef]
- Lilioso, M.; Pires-Silva, D.; von Hertwig, M.F.F.; Oliveira, J.; da Rosa, J.A.; Vilela, R.V.; Folly-Ramos, E.; Almeida, C.E. Triatoma petrocchiae (Hemiptera, Reduviidae, Triatominae): A Chagas disease vector of T. brasiliensis species complex associated to reptiles. Infect. Genet. Evol. 2020, 82, 104307. [Google Scholar] [CrossRef]
- Lima-Neiva, V.; Toma, H.K.; Aguiar, L.M.A.; Lopes, C.M.; Dias, L.P.; Gonçalves, T.C.M.; Costa, J. The connection between Trypanosoma cruzi transmission cycles by Triatoma brasiliensis brasiliensis: A threat to human health in an area susceptible to desertification in the Seridó, Rio Grande do Norte, Brazil. PLoS Negl. Trop. Dis. 2021, 15, e0009919. [Google Scholar] [CrossRef]
- Costa, J.; Lorenzo, M. Biology, diversity and strategies for the monitoring and control of triatomines—Chagas disease vectors. Mem. Inst. Oswaldo Cruz 2009, 1, 46–51. [Google Scholar] [CrossRef] [Green Version]
- Silveira, A.C.; Dias, J.C.P. O controle da transmissão vetorial. Rev. Soc. Bras. Med. Trop. 2011, 44, 52–63. (In Portuguese) [Google Scholar] [CrossRef]
- Lima-Oliveira, T.M.; Fontes, F.V.H.M.; Lilioso, M.; Pires-Silva, D.; Teixeira, M.M.G.; Meza, J.G.B.; Harry, M.; Fileé, J.; Costa, J.; Valença-Barbosa, C.; et al. Molecular eco-epidemiology on the sympatric Chagas disease vectors Triatoma brasiliensis and Triatoma petrocchiae: Ecotopes, genetic variation, natural infection prevalence by trypanosomatids and parasite genotyping. Acta Trop. 2020, 201, 105188. [Google Scholar] [CrossRef]
- Kamimura, E.H.; Viana, M.C.; Lilioso, M.; Fontes, F.H.M.; Pires-Silva, D.; Valença-Barbosa, C.; Carbajal-de-la-Fuente, A.L.; Folly-Ramos, E.; Solferin, V.N.; Thyssen, P.J.; et al. Drivers of molecular and morphometric variation in Triatoma brasiliensis (Hemiptera: Triatominae): The resolution of geometric morphometrics for populational structuring on a microgeographical scale. Parasit Vectors 2020, 13, 455. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, J.; Marcet, P.L.; Takiya, D.M.; Mendonça, V.J.; Belintani, T.; Bargues, M.D.; Mateo, L.; Chagas, V.; Folly-Ramos, E.; Cordeiro-Estrela, P.; et al. Combined phylogenetic and morphometric information to delimit and unify the Triatoma brasiliensis species complex and the brasiliensis subcomplex. Acta Trop. 2017, 170, 140–148. [Google Scholar] [CrossRef] [PubMed]
- Belintani, T.; Oliveira, J.; Pinotti, H.; Silva, L.A.; Alevi, K.C.C.; Galvão, C.; Rosa, J.A. Phylogenetic and phenotypic relationships of the Triatoma sordida subcomplex (Hemiptera: Reduviidae: Triatominae). Acta Trop. 2020, 212, 105679. [Google Scholar] [CrossRef] [PubMed]
- Dujardin, J.P.; Bermudez, H.; Asine, C.; Schofield, C.J.; Tibayrenc, M. Metric differences between silvatic and domestic Triatoma infestans (Heteroptera- Reduviidae) in Bolivia. J. Med. Entomol. 1997, 34, 544–551. [Google Scholar] [CrossRef]
- Dujardin, J.P.; Chávez, T.; Moreno, J.M.; Machane, M.; Noireau, F.; Schofield, C.J. Comparison of isoenzyme electrophoresis and morphometric analysis for phylogenetic reconstruction of the Rhodniini (Hemiptera: Reduvidae: Triatominae). J. Med. Entomol. 1999, 36, 653–659. [Google Scholar] [CrossRef]
- Lunardi, R.R.; Benítez, H.A.; Câmara, T.P.; Gomes, L.P.; Arrais-Silva, W.W. Head shape variation in response to diet in Triatoma williami (Hemiptera, Reduviidae: Triatominae), a possible Chagas disease vector of legal Amazônia. Zoo. Anz. 2017, 267, 187–193. [Google Scholar] [CrossRef]
- Vilaseca, C.; Méndez, M.A.; Pinto, C.F.; Benítez, H.A. Assessment of patterns of geometric shape variation in Triatoma infestans Hemiptera: Reduviidae: A first report in populations from Bolivia. Insects 2020, 11, 274. [Google Scholar] [CrossRef]
- Vilaseca, C.; Méndez, M.A.; Pinto, C.F.; Lemic, D.; Benítez, H.A. Unraveling the Morphological Variation of Triatoma infestans in the Peridomestic Habitats of Chuquisaca Bolivia: A Geometric Morphometric Approach. Insects 2021, 12, 185. [Google Scholar] [CrossRef]
- Costa, J.; Peterson, A.T.; Dujardin, J.P. Morphological evidence suggests homoploid hybridization as a possible mode of speciation in the Triatominae (Hemiptera, Heteroptera, Reduviidae). Infect. Genet. Evol. 2009, 9, 263–270. [Google Scholar] [CrossRef]
- Lent, H.; Wygodzinsky, P.W. Revision of the Triatominae (Hemiptera, Reduviidae), and their significance as vectors of Chagas’ disease. Bull. Am. Mus. Nat. Hist. 1979, 163, 3. [Google Scholar]
- Rohlf, J. On Applications of Geometric Morphometrics to Studies of Ontogeny and Phylogeny. Syst. Biol. 1998, 47, 147–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rohlf, F.J. Tpsdig, Digitize Landmarks and Outlines, Version 2.17; Department of Ecology and Evolution: State University of New York at Stony Brook, NY, USA, 2013. [Google Scholar]
- Rohlf, F.J.; Slice, D. Extensions of the Procustes methods for the optimal superimposition of landmarks. Syst. Zool. 1990, 39, 40–59. [Google Scholar] [CrossRef] [Green Version]
- Dryden, I.L.; Mardia, K.V. Statistical Shape Analysis; John Wiley and Sons: Hoboken, NJ, USA, 1998; Volume 4. [Google Scholar]
- Rohlf, J. General Biology; The University of Chicago Press: Chicago, IL, USA, 2010; p. 85. [Google Scholar]
- Fruciano, C. Measurement error in geometric morphometrics. Dev. Genes Evol. 2016, 226, 139–158. [Google Scholar] [CrossRef] [PubMed]
- Moraes, D.A. A Morfometria Geométrica e a “Revolução na Morfometria”: Localizando mudanças na forma dos organismos. Bioletim 2003, 3, 14–20. (In Portuguese) [Google Scholar]
- Villalobos-Leiva, A.; Benítez, H.A. Morfometría Geométrica y sus Nuevas Aplicaciones en Ecología y Biología Evolutiva: Parte 2. Int. J. Morphol. 2020, 38, 1818–1836. (In Spanish) [Google Scholar] [CrossRef]
- Hair, J.F.; Tatham, R.L.; Anderson, R.E.; Black, W. Multivariate Data Analysis. Prentice Hall 1998, 5, 768. [Google Scholar]
- Bookstein, F.L. Morphometric Tools for Landmark Data: Geometry and Biology; Cambridge University Press: Cambridge, UK, 1991; p. 435. [Google Scholar]
- Hansen, T.F.; Houle, D. Evolvability, Stabilizing Selection, and the Problem of Stasis. In Phenotypic Integration: Studying the Ecology and Evolution of Complex Phenotypes; Oxford University Press: Oxford, UK, 2004; pp. 130–150. [Google Scholar]
- Houle, D.; Bolstad, G.H.; van der Linde, K.; Hansen, T.F. Mutation predicts 40 million years of fly wing evolution. Nature 2017, 548, 447–450. [Google Scholar] [CrossRef]
- Benitez, H.A.; Püschel, T.; Suazo, M. Drosophila wing integration and modularity: A multi-level approach to understand the history of morphological structures. Biology 2022, 11, 567. [Google Scholar] [CrossRef]
- Gumiel, M.S.; Catalá, F.; Noireau, A.; Rojas, D.A.; García, A.; Dujardin, J.P. Wing geometry in Triatoma infestans (Klug) and T. melanosoma Martinez, Olmedo & Carcavallo (Hemiptera: Reduviidae). Syst. Entomol. 2003, 28, 2–6. [Google Scholar] [CrossRef]
- Schachter-Broide, J.; Gürtler, R.E.; Kitron, U.; Dujardin, J.P. Temporal variations of wing size and shape of Triatoma infestans (Hemiptera: Reduviidae) populations from northwestern Argentina using geometric morphometry. J. Med. Entomol. 2009, 46, 994–1000. [Google Scholar] [CrossRef] [Green Version]
- Nattero, J.; Piccinali, R.V.; Lopes, C.M.; Hernández, M.L.; Abrahan, L.; Lobbia, P.A.; Rodríguez, C.S.; Carbajal-de-la-Fuente, A.L. Morphometric variability among the species of the Sordida subcomplex (Hemiptera: Reduviidae: Triatominae): Evidence for differentiation across the distribution range of Triatoma sordida. Parasit Vectors 2017, 10, 412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dujardin, J.P.; Costa, J.; Bustamante, D.; Jaramillo, N.; Catalá, S. Deciphering morphology in Triatominae: The evolutionary signals. Acta Trop. 2009, 110, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, F.A.; Donnelly, M.J.; Beard, C.B.; Costa, J. Nested clade and phylogeographic analyses of the Chagas disease vector Triatoma brasiliensis in Northeast Brazil. Mol. Phylogenet. Evol. 2004, 32, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Gurgel-Gonçalves, R.; Ferreira, J.B.C.; Rosa, A.F.; Bar, M.E.; Galvão, C. Geometric morphometrics and ecological niche modelling for delimitation of near-sibling triatomine species. Med. Vet. Entomol. 2011, 25, 84–93. [Google Scholar] [CrossRef]
- Paschoaletto, L.; Lima-Neiva, V.; Lopes, C.M.; Dale, C.; Mallet, J.R.S.; La Fuente, A.L.C.A.; Costa, J. 72 years of temporal analysis through geometric morphometrics detects phenotypic variation in populations of Triatoma infestans (Klug, 1834). Acta Biol. Par. 2020, 49, 1–17. [Google Scholar] [CrossRef]
- Forattini, O.P. Biogeography, origin, and distribution of triatominae domiciliarity in Brazil. Rev. Saúde Pública 2006, 40, 964–998. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Year | Collection | State | City | Males | Females | Total |
|---|---|---|---|---|---|---|---|
| Group 1 | 1912 | CTIOC | Piauí | Floriano | 5 | 1 | 6 |
| Group 2 | 1922 | CTIOC | Ceará | Quixada | 1 | 0 | 2 |
| 1929 | Rio Grande do Norte | Acary | 1 | 0 | |||
| Group 3 | 1940 | CTIOC | Ceará | Russas | 12 | 6 | 19 |
| 1941 | Pernambuco | Mangabeira | 1 | 0 | |||
| Group 4 | 1950 | CTIOC | Paraíba | Santa Lúcia | 12 | 7 | 22 |
| 1955 | Santa Lúcia | 1 | 2 | ||||
| Group 5 | 1977 | CTIOC | Piauí | Brejo Seco | 1 | 3 | 4 |
| Group 6 | 1996 | CEIOC CTIOC CTIOC | Rio Grande do Norte | Serra Negra do Norte, Caicó | 4 | 5 | 24 |
| 1998 | Piaui | Brejo Seco | 2 | 2 | |||
| 1999 | Piaui | Brejo Seco | 2 | 9 | |||
| Group 7 | 2002 | CEIOC | Paraíba | Livramento, São José da Lagoa da Tapada, São José das Piranhas, São Francisco da Prata | 16 | 15 | 31 |
| Group 8 | 2014 | CTIOC | Ceará | Santa Quitéria | 1 | 2 | 3 |
| TOTAL | 111 | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paschoaletto, L.; Dale, C.; Lima-Neiva, V.; Carbajal-de-la-Fuente, A.L.; de Oliveira, J.; Benítez, H.A.; Costa, J. Morphological Stasis in Time? A Triatoma brasiliensis brasiliensis Study Using Geometric Morphometrics in the Long Run. Animals 2022, 12, 1362. https://doi.org/10.3390/ani12111362
Paschoaletto L, Dale C, Lima-Neiva V, Carbajal-de-la-Fuente AL, de Oliveira J, Benítez HA, Costa J. Morphological Stasis in Time? A Triatoma brasiliensis brasiliensis Study Using Geometric Morphometrics in the Long Run. Animals. 2022; 12(11):1362. https://doi.org/10.3390/ani12111362
Chicago/Turabian StylePaschoaletto, Letícia, Carolina Dale, Vanessa Lima-Neiva, Ana Laura Carbajal-de-la-Fuente, Jader de Oliveira, Hugo A. Benítez, and Jane Costa. 2022. "Morphological Stasis in Time? A Triatoma brasiliensis brasiliensis Study Using Geometric Morphometrics in the Long Run" Animals 12, no. 11: 1362. https://doi.org/10.3390/ani12111362