
Biomarkers and De Novo Protein Design Can Improve Precise Amino Acid Nutrition in Broilers

 ,
,  , and
, and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Case Study 1: Use of SUN Content as a Rapid Biomarker of Amino Acid Imbalances
2.1. Background
2.2. Animals and Experimental Procedure
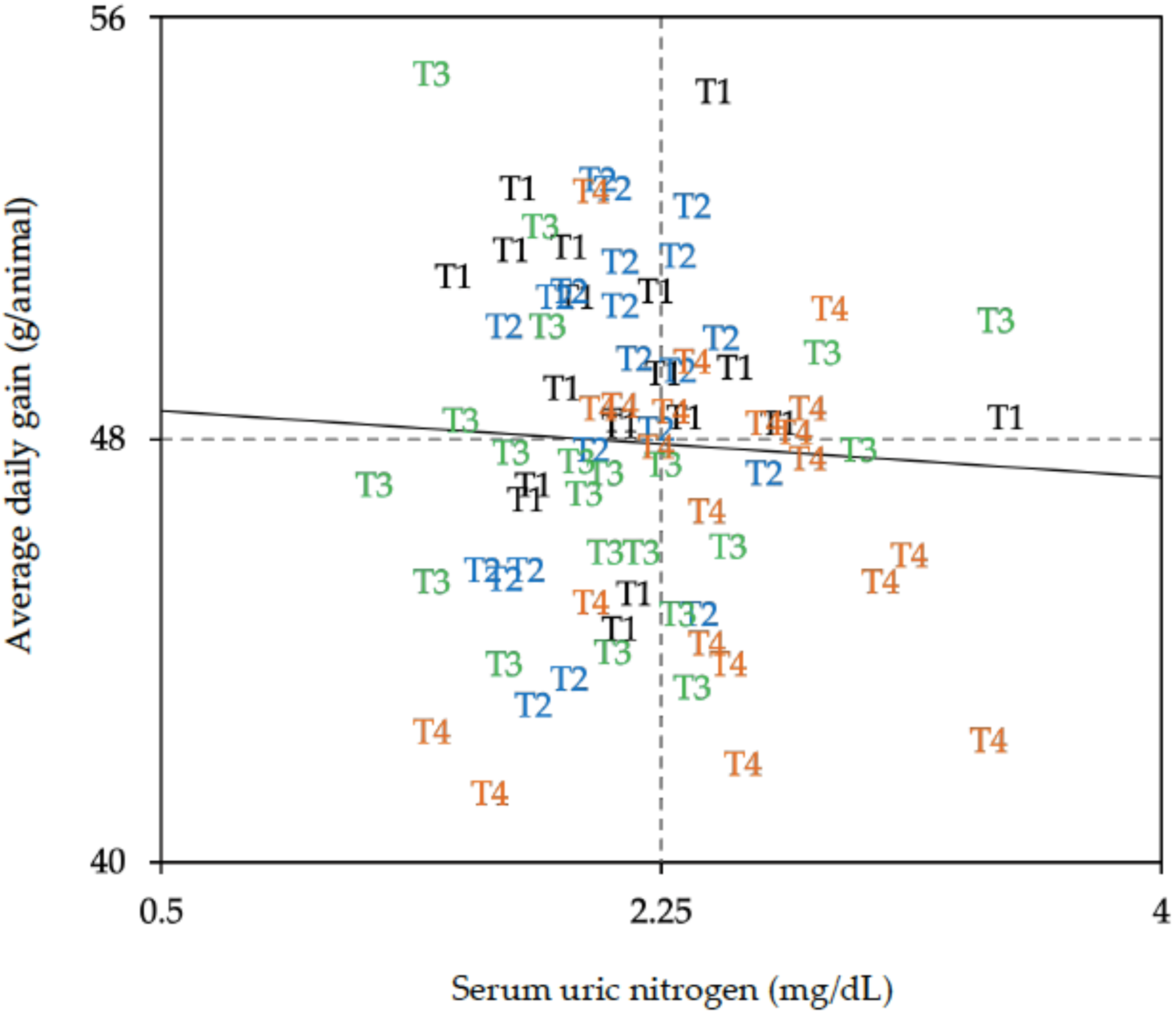
2.3. Results
2.4. Discussion
3. Case Study 2: De Novo Protein Design, an Example of an Ideal Protein for 21-Day-Old Broilers
3.1. Background
3.2. Experimental Procedure
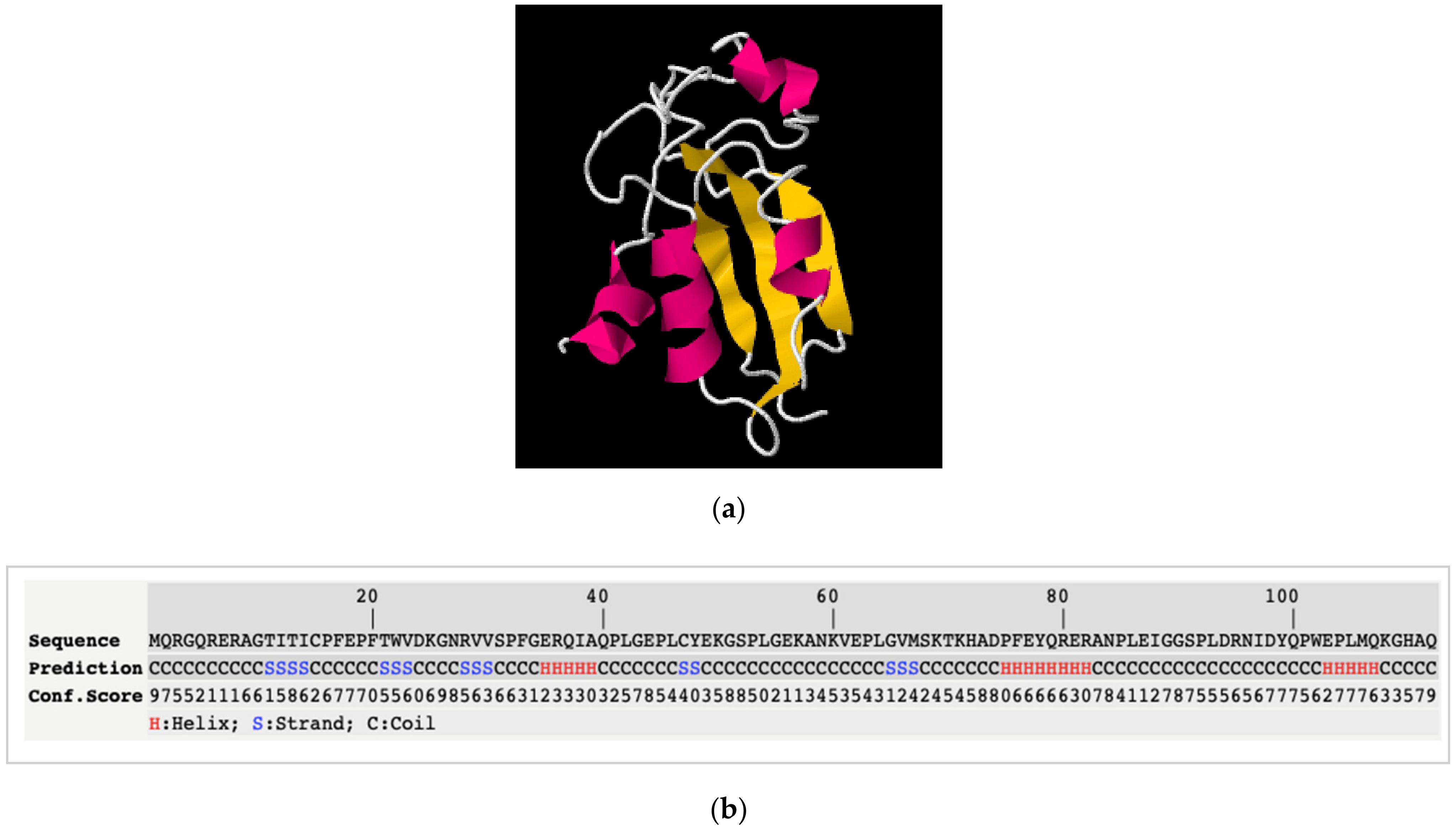
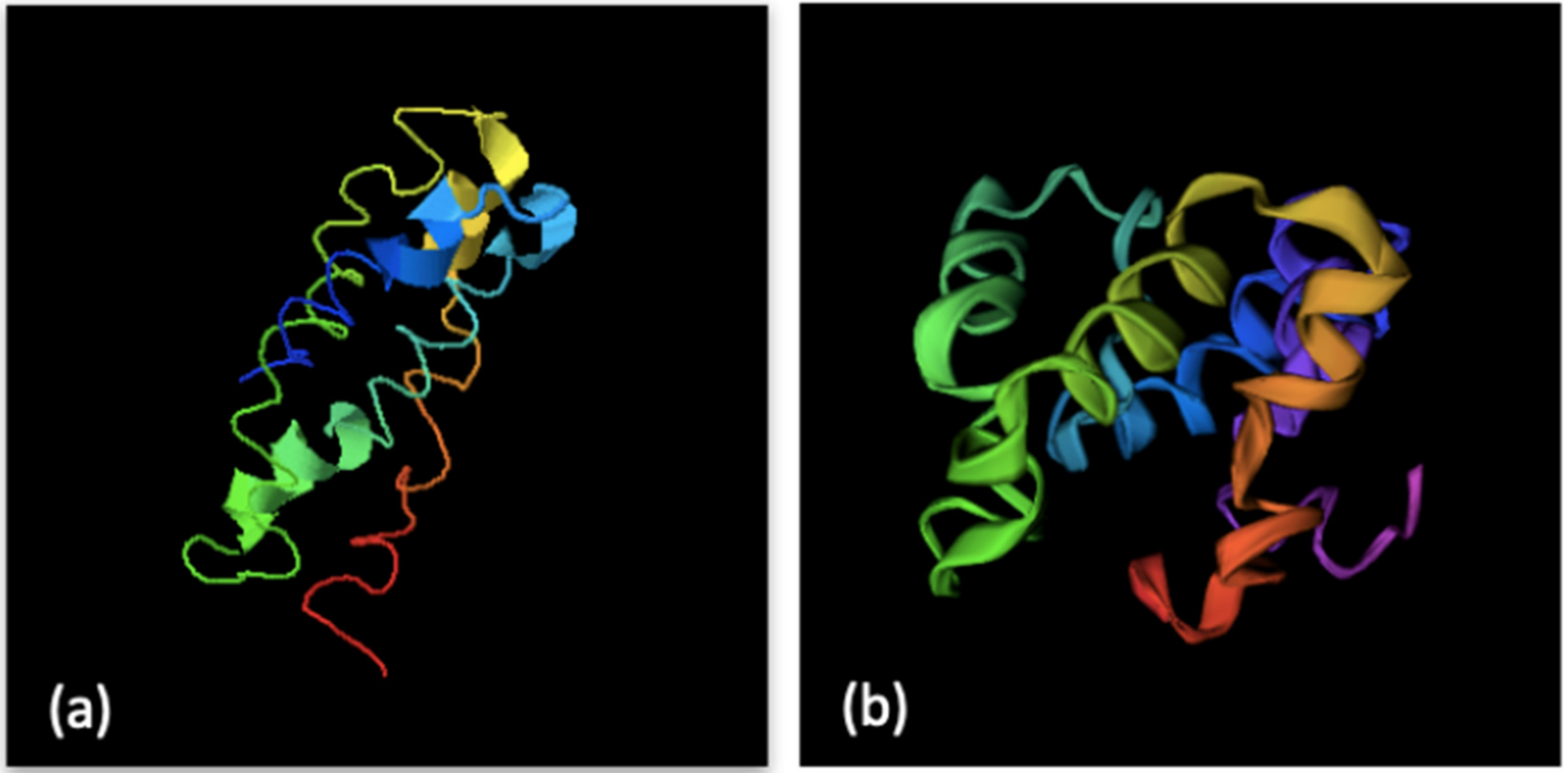
3.3. Results
3.4. Discussion
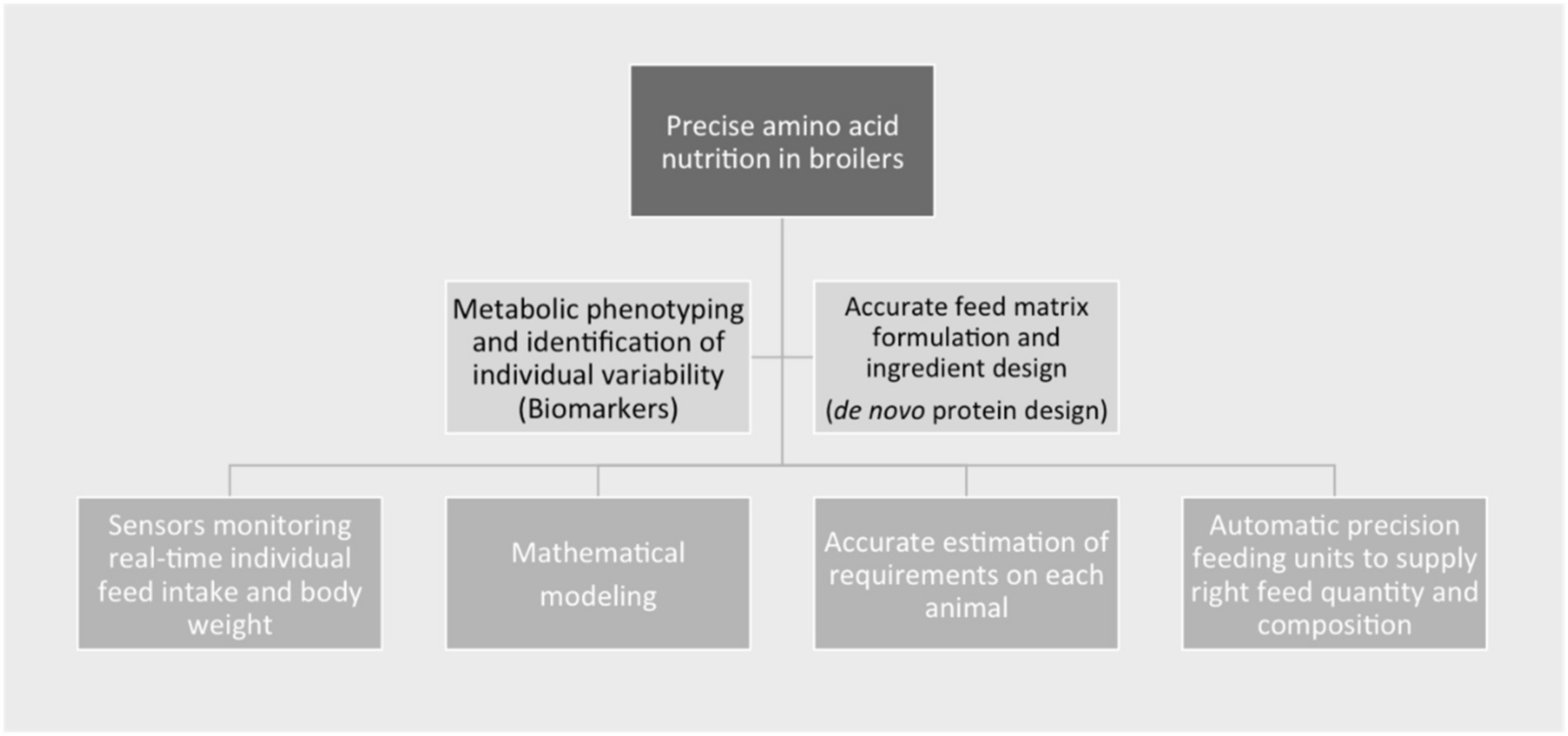
4. Future Precision Nutrition Needs to Improve Protein Nutrition in Broilers
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sibbald, I.R. A Bioassay for Available Amino Acids and True Metabolizable Energy in Feedstuffs. Poult. Sci. 1979, 58, 668–673. [Google Scholar] [CrossRef]
- Maharjan, P.; Liang, Y. Precision Livestock Farming: The Opportunities in Poultry Sector. J. Agric. Sci. Technol. A 2020, 10, 45–53. [Google Scholar] [CrossRef]
- Banhazi, T.; Lehr, H.; Black, J.; Crabtree, H.; Schofield, C.; Tscharke, M.; Berckmans, D. Precision Livestock Farming: An International Review of Scientific and Commercial Aspects. Int. J. Agric. Biol. Eng. 2012, 5, 1. [Google Scholar] [CrossRef]
- Moss, A.F.; Chrystal, P.V.; Cadogan, D.J.; Wilkinson, S.J.; Crowley, T.M.; Choct, M. Precision Feeding and Precision Nutri-tion: A Paradigm Shift in Broiler Feed Formulation? Anim. Biosci. 2021, 34, 354–362. [Google Scholar] [CrossRef] [PubMed]
- Warren, W.A.; Emmert, J.L. Efficacy of Phase-Feeding in Supporting Growth Performance of Broiler Chicks during the Star-ter and Finisher Phases. Poult. Sci. 2000, 79, 764–770. [Google Scholar] [CrossRef]
- Hauschild, L.; Bueno, C.F.D.; Remus, A.; Gobi, J.D.P.; Isola, R.D.G.; Sakomura, N.K. Multiphase Feeding Program for Broilers Can Replace Traditional System. Sci. Agric. 2015, 72, 210–214. [Google Scholar] [CrossRef]
- Currie, R.; Maclsaac, J.; Rathgeber, B. Increasing the Number of Phases Fed to Broiler Chickens by Blending Diets. Poult. Sci. 2006, 85, 52. [Google Scholar]
- Pomar, C.; Remus, A. Precision Pig Feeding: A Breakthrough toward Sustainability. Anim. Front. 2019, 9, 52–59. [Google Scholar] [CrossRef]
- Halachmi, I.; Guarino, M. Editorial: Precision Livestock Farming: A ‘per Animal’ Approach Using Advanced Monitoring Technologies. Animal 2016, 10, 1482–1483. [Google Scholar] [CrossRef]
- Liu, S.Y.; Macelline, S.P.; Chrystal, P.V.; Selle, P.H. Progress towards Reduced-Crude Protein Diets for Broiler Chickens and Sustainable Chicken-Meat Production. J. Anim. Sci. Biotechnol. 2021, 12, 20. [Google Scholar] [CrossRef]
- Emmert, J.L.; Baker, D.H. Use of the Ideal Protein Concept for Precision Formulation of Amino Acid Levels in Broiler Diets. J. Appl. Poult. Res. 1997, 6, 462–470. [Google Scholar] [CrossRef]
- Ferket, P.R.; van Heugten, E.; van Kempen, T.A.T.G.; Angel, R. Nutritional Strategies to Reduce Environmental Emissions from Nonruminants. J. Anim. Sci. 2002, 80, E168–E182. [Google Scholar] [CrossRef]
- Powers, W.; Angel, R. A Review of the Capacity for Nutritional Strategies to Address Environmental Challenges in Poultry Production. Poult. Sci. 2008, 87, 1929–1938. [Google Scholar] [CrossRef] [PubMed]
- Macfarlane, G.T.; Macfarlane, S. Bacteria, Colonic Fermentation, and Gastrointestinal Health. J. AOAC Int. 2012, 95, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, M.S.; Ijssennagger, N.; Kies, A.K.; van Mil, S.W.C. Protein Fermentation in the Gut; Implications for Intestinal Dys-function in Humans, Pigs, and Poultry. Am. J. Physiol. Gastrointest. Liver Physiol. 2018, 315, G159–G170. [Google Scholar] [CrossRef]
- Celi, P.; Cowieson, A.J.; Fru-Nji, F.; Steinert, R.E.; Kluenter, A.-M.; Verlhac, V. Gastrointestinal Functionality in Animal Nu-trition and Health: New Opportunities for Sustainable Animal Production. Anim. Feed Sci. Technol. 2017, 234, 88–100. [Google Scholar] [CrossRef]
- Bindari, Y.R.; Gerber, P.F. Centennial Review: Factors Affecting the Chicken Gastrointestinal Microbial Composition and Their Association with Gut Health and Productive Performance. Poult. Sci. 2022, 101, 101612. [Google Scholar] [CrossRef]
- McDonald, P.; Edwards, R.A.; Greenhalgh, J.F.D.; Morgan, C.A.; Sinclair, L.A.; Wilkinson, L.G. Animal Nutrition, 7th ed.; Pearson, Prentice Hall: Hoboken, NJ, USA, 2011. [Google Scholar]
- Steed, J.R.; Romero-Sanchez, H.; Han, Y.; Page, G.I.; Davis, A.J. Validation of NutriOpt Dietary Formulation Strategies on Broiler Growth and Economic Performance. J. Appl. Poult. Res. 2020, 29, 314–327. [Google Scholar] [CrossRef]
- Halas, V.; Dukhta, G. Growth Models and Their Application in Precision Feeding of Monogastric Farm Animals. Acta Fytotechn. Zootechn. 2020, 23, 258–264. [Google Scholar] [CrossRef]
- Coma, J.; Zimmerman, D.R.; Carrion, D. Lysine Requirement of the Lactating Sow Determined by Using Plasma Urea Nitro-gen as a Rapid Response Criterion. J. Anim. Sci. 1996, 74, 1056. [Google Scholar] [CrossRef]
- Nieto, R.; Barea, R.; Lara, L.; Palma-Granados, P.; Aguilera, J.F. Lysine Requirement Relative to Total Dietary Protein for Optimum Performance and Carcass Protein Deposition of Iberian Piglets. Anim. Feed Sci. Technol. 2015, 206, 48–56. [Google Scholar] [CrossRef]
- Roth-Maier, D.A.; Ott, H.; Roth, F.X.; Paulicks, B.R. Effects of the Level of Dietary Valine Supply on Amino Acids and Urea Concentration in Milk and Blood Plasma of Lactating Sows: Valine for Lactating Sows. J. Anim. Physiol. Anim. Nutr. 2004, 88, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Marín-García, P.J.; López-Luján, M.D.C.; Ródenas, L.; Martínez-Paredes, E.; Blas, E.; Pascual, J.J. Plasma Urea Nitrogen as an Indicator of Amino Acid Imbalance in Rabbit Diets. World Rabbit Sci. 2020, 28, 63. [Google Scholar] [CrossRef]
- Donsbough, A.L.; Powell, S.; Waguespack, A.; Bidner, T.D.; Southern, L.L. Uric Acid, Urea, and Ammonia Concentrations in Serum and Uric Acid Concentration in Excreta as Indicators of Amino Acid Utilization in Diets for Broilers. Poult. Sci. 2010, 89, 287–294. [Google Scholar] [CrossRef]
- Ravindran, V.; Bryden, W.L. Amino Acid Availability in Poultry—In Vitro and in Vivo Measurements. Aust. J. Agric. Res. 1999, 50, 889. [Google Scholar] [CrossRef]
- Vieira, S.L.; Lemme, A.; Goldenberg, D.B.; Brugalli, I. Responses of Growing Broilers to Diets with Increased Sulfur Amino Acids to Lysine Ratios at Two Dietary Protein Levels. Poult. Sci. 2004, 83, 1307–1313. [Google Scholar] [CrossRef]
- Han, Y.; Baker, D.H. Digestible Lysine Requirement of Male and Female Broiler Chicks During the Period Three to Six Weeks Posthatching. Poult. Sci. 1994, 73, 1739–1745. [Google Scholar] [CrossRef]
- Han, Y.; Baker, D.H. Effects of Sex, Heat Stress, Body Weight, and Genetic Strain on the Dietary Lysine Requirement of Broi-ler Chicks. Poult. Sci. 1993, 72, 701–708. [Google Scholar] [CrossRef]
- Han, Y.; Baker, D.H. Lysine Requirements of Fast- and Slow-Growing Broiler Chicks. Poult. Sci. 1991, 70, 2108–2114. [Google Scholar] [CrossRef]
- Samadi; Liebert, F. Modelling the Optimal Lysine to Threonine Ratio in Growing Chickens Depending on Age and Efficiency of Dietary Amino Acid Utilisation. Br. Poult. Sci. 2008, 49, 45–54. [Google Scholar] [CrossRef]
- Dozier, W.A.; Payne, R.L. Digestible Lysine Requirements of Female Broilers from 1 to 15 Days of Age. J. Appl. Poult. Res. 2012, 21, 348–357. [Google Scholar] [CrossRef]
- Agostini, P.S.; Santos, R.R.; Khan, D.R.; Siebert, D.; van der Aar, P. The Optimum Valine: Lysine Ratios on Performance and Carcass Traits of Male Broilers Based on Different Regression Approaches. Poult. Sci. 2019, 98, 1310–1320. [Google Scholar] [CrossRef] [PubMed]
- Zampiga, M.; Soglia, F.; Petracci, M.; Meluzzi, A.; Sirri, F. Effect of Different Arginine-to-Lysine Ratios in Broiler Chicken Diets on the Occurrence of Breast Myopathies and Meat Quality Attributes. Poult. Sci. 2019, 98, 2691–2697. [Google Scholar] [CrossRef] [PubMed]
- Belloir, P.; Méda, B.; Lambert, W.; Corrent, E.; Juin, H.; Lessire, M.; Tesseraud, S. Reducing the CP Content in Broiler Feeds: Impact on Animal Performance, Meat Quality and Nitrogen Utilization. Animal 2017, 11, 1881–1889. [Google Scholar] [CrossRef]
- Sigolo, S.; Deldar, E.; Seidavi, A.; Bouyeh, M.; Gallo, A.; Prandini, A. Effects of Dietary Surpluses of Methionine and Lysine on Growth Performance, Blood Serum Parameters, Immune Responses, and Carcass Traits of Broilers. J. Appl. Anim. Res. 2019, 47, 146–153. [Google Scholar] [CrossRef]
- Kriseldi, R.; Tillman, P.B.; Jiang, Z.; Dozier, W.A. Effects of Feeding Reduced Crude Protein Diets on Growth Performance, Nitrogen Excretion, and Plasma Uric Acid Concentration of Broiler Chicks during the Starter Period. Poult. Sci. 2018, 97, 1614–1626. [Google Scholar] [CrossRef]
- Berres, J.; Vieira, S.L.; Kidd, M.T.; Taschetto, D.; Freitas, D.M.; Barros, R.; Nogueira, E.T. Supplementing L-Valine and L-Isoleucine in Low-Protein Corn and Soybean Meal All-Vegetable Diets for Broilers. J. Appl. Poult. Res. 2010, 19, 373–379. [Google Scholar] [CrossRef]
- Yuan, J.; Karimi, A.; Zornes, S.; Goodgame, S.; Mussini, F.; Lu, C.; Waldroup, P.W. Evaluation of the Role of Glycine in Low-Protein Amino Acid-Supplemented Diets. J. Appl. Poult. Res. 2012, 21, 726–737. [Google Scholar] [CrossRef]
- Dilger, R.N.; Bryant-Angeloni, K.; Payne, R.L.; Lemme, A.; Parsons, C.M. Dietary Guanidino Acetic Acid Is an Efficacious Replacement for Arginine for Young Chicks. Poult. Sci. 2013, 92, 171–177. [Google Scholar] [CrossRef]
- Jahanian, R. Immunological Responses as Affected by Dietary Protein and Arginine Concentrations in Starting Broiler Chicks. Poult. Sci. 2009, 88, 1818–1824. [Google Scholar] [CrossRef]
- Mejia, L.; Zumwalt, C.D.; Tillman, P.B.; Shirley, R.B.; Corzo, A. Ratio Needs of Arginine Relative to Lysine of Male Broilers from 28 to 42 Days of Age during a Constant, Elevated Environmental Temperature Regimen. J. Appl. Poult. Res. 2012, 21, 305–310. [Google Scholar] [CrossRef]
- Attia, Y.A.; Bovera, F.; Wang, J.; Al-Harthi, M.A.; Kim, W.K. Multiple Amino Acid Supplementations to Low-Protein Diets: Effect on Performance, Carcass Yield, Meat Quality and Nitrogen Excretion of Finishing Broilers under Hot Climate Conditions. Animal 2020, 10, 973. [Google Scholar] [CrossRef] [PubMed]
- Vieira, S.L.; Angel, C.R. Optimizing Broiler Performance Using Different Amino Acid Density Diets: What Are the Limits? J. Appl. Poult. Res. 2012, 21, 149–155. [Google Scholar] [CrossRef]
- Carbonaro, M.; Maselli, P.; Nucara, A. Relationship between Digestibility and Secondary Structure of Raw and Thermally Treated Legume Proteins: A Fourier Transform Infrared (FT-IR) Spectroscopic Study. Amino Acids 2012, 43, 911–921. [Google Scholar] [CrossRef]
- Esteve, C. Towards Precision Poultry Feeding: Amino Acid Losses in the Body of Broiler Chickens Fed a Nitrogen-Free Diet. Master’s Thesis, Universitat Politècnica de València, Valencia, Spain, 2021. Available online: http://hdl.handle.net/10251/174720 (accessed on 28 February 2022).
- Wu, G. Dietary Requirements of Synthesizable Amino Acids by Animals: A Paradigm Shift in Protein Nutrition. J. Anim. Sci. Biotechnol. 2014, 5, 34. [Google Scholar] [CrossRef]
- National Research Council (NRC). Nutrient Requirements of Poultry, 9th ed.; National Academy Press: Washington, DC, USA, 1994. [Google Scholar]
- Santomá, G.; Mateos, G.G. Necesidades Nutricionales En Avicultura, Normas FEDNA, 2nd ed.; Fundación Española para el Desarrollo de la Nutrición Animal: Madrid, Spain, 2018. [Google Scholar]
- Veeroederbureau, C. Table Booklet Feeding of Poultry: Feeding Standards, Feeding Advices and Nutritional Values of Feed Ingredients; CVB Series; Federatie Nederlandse Diervoederketen: Zoetermeer, The Netherlands, 2018. [Google Scholar]
- Rostagno, H.S.; Texeira Albino, L.S.; Hannas, M.I.; Lopes Donzele, J.; Sakomura, N.; Perazzo, F.G.; Oliveira Brito, C. Tablas Brasileñas Para Aves y Cerdos, 4th ed.; Departamento de Zootecnia: Viçosa, Brazil, 2017. [Google Scholar]
- Grishaeva, T.M.; Bogdanov, Y.F. On the Origin of Synaptonemal Complex Proteins. Search for Related Proteins in Proteo-mes of Algae, Lower Fungi, Mosses, and Protozoa. Russ. J. Genet. Appl. Res. 2013, 3, 481–486. [Google Scholar] [CrossRef]
- Singh, M.; Kumar, V.; Sikka, K.; Thakur, R.; Harioudh, M.K.; Mishra, D.P.; Ghosh, J.K.; Siddiqi, M.I. Computational Design of Biologically Active Anticancer Peptides and Their Interactions with Heterogeneous POPC/POPS Lipid Membranes. J. Chem. Inf. Model. 2020, 60, 332–341. [Google Scholar] [CrossRef]
- Recoules, E.; Sabboh-Jourdan, H.; Narcy, A.; Lessire, M.; Harichaux, G.; Labas, V.; Duclos, M.J.; Réhault-Godbert, S. Exploring the in Vivo Digestion of Plant Proteins in Broiler Chickens. Poult. Sci. 2017, 96, 1735–1747. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, Y. Protein Structure and Function Prediction Using I-TASSER. Curr. Protoc. Bioinform. 2015, 52, 5–8. [Google Scholar] [CrossRef]
- Zhang, W.; Yang, J.; He, B.; Walker, S.E.; Zhang, H.; Govindarajoo, B.; Virtanen, J.; Xue, Z.; Shen, H.-B.; Zhang, Y. Integration of QUARK and I-TASSER for Ab Initio Protein Structure Prediction in CASP11: Ab Initio Structure Prediction in CASP11. Proteins 2016, 84, 76–86. [Google Scholar] [CrossRef]
- Lledó, C. De Novo Design of an Ideal Protein for Feeding Meat Chickens from 0 to 21 Days. Bachelor’s Thesis, Universitat Politècnica de València, Valencia, Spain, 2020. Available online: http://hdl.handle.net/10251/149623 (accessed on 28 February 2022).
- Esmail, S.H. Understanding Protein Requirements. 2016. Available online: https://www.poultryworld.net/Nutrition/Articles/2016/11/Understanding-protein-requirements-2914798W/ (accessed on 20 May 2020).
- Gabriel, I.; Quillien, L.; Cassecuelle, F.; Marget, P.; Juin, H.; Lessire, M.; Sève, B.; Duc, G.; Burstin, J. Variation in Seed Protein Digestion of Different Pea (Pisum Sativum L.) Genotypes by Cecectomized Broiler Chickens: 2. Relation between in Vivo Protein Digestibility and Pea Seed Characteristics, and Identification of Resistant Pea Polypeptides. Livest. Sci. 2008, 113, 262–273. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, Z.; Wang, R.; Sui, X.; Qi, B.; Han, F.; Li, Y.; Jiang, L. Secondary Structure and Subunit Composition of Soy Protein In Vitro Digested by Pepsin and Its Relation with Digestibility. Biomed. Res. Int. 2016, 2016, 5498639. [Google Scholar] [CrossRef] [PubMed]
- Bai, M.; Qin, G.; Sun, Z.; Long, G. Relationship between Molecular Structure Characteristics of Feed Proteins and Protein in Vitro Digestibility and Solubility. Asian Australas. J. Anim. Sci. 2015, 29, 1159–1165. [Google Scholar] [CrossRef]
- Boodhoo, K.V.K.; Flickinger, M.C.; Woodley, J.M.; Emanuelsson, E.A.C. Bioprocess Intensification: A Route to Efficient and Sustainable Biocatalytic Transformations for the Future. Chem. Eng. Process. Process Intensif. 2022, 172, 108793. [Google Scholar] [CrossRef]
- Monck, C.; Elani, Y.; Ceroni, F. Cell-Free Protein Synthesis: Biomedical Applications and Future Perspectives. Chem. Eng. Res. Des. 2022, 177, 653–658. [Google Scholar] [CrossRef]
- Banhazi, T.; Babinszky, L.; Halas, V.; Tscharke, M. Precision Livestock Farming: Precision Feeding Technologies and Sustainable Animal Production. Int. J. Agric. Biol. Eng. 2012, 5, 54–61. [Google Scholar] [CrossRef]
- Pomar, C.; van Milgen, J.; Remus, A. Precision Livestock Feeding, Principle and Practice. In Poultry and Pig Nutrition: Challenges of the 21st Century; Hendriks, W.H., Verstegen, M.W.A., Babinszky, L., Eds.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2019; ISBN 978-90-8686-333-4. [Google Scholar]
- Zuidhof, M.J.; Fedorak, M.V.; Ouellette, C.A.; Wenger, I.I. Precision Feeding: Innovative Management of Broiler Breeder Feed Intake and Flock Uniformity. Poult. Sci. 2017, 96, 2254–2263. [Google Scholar] [CrossRef]
- Li, G.; Zhao, Y.; Hailey, R.; Zhang, N.; Liang, Y.; Purswell, J.L. An Ultra-High Frequency Radio Frequency Identification System for Studying Individual Feeding and Drinking Behaviors of Group-Housed Broilers. Animal 2019, 13, 2060–2069. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Crude Protein (%) | Valine (%) | Arginine (%) | Valine to Lysine | Arginine to Lysine | |
|---|---|---|---|---|---|
| T1 | 20.00 | 0.896 | 1.228 | 0.815 | 1.116 |
| T2 | 18.13 | 0.873 | 1.136 | 0.806 | 1.049 |
| T3 | 18.07 | 0.770 | 1.136 | 0.711 | 1.049 |
| T4 | 17.88 | 0.874 | 1.004 | 0.807 | 0.927 |
| T1 | T2 | T3 | T4 | p-Value | |
|---|---|---|---|---|---|
| SUN (mg/dL) 1 | 1.96 ± 0.1 a | 1.89 ± 0.1 a | 1.90 ± 0.1 a | 2.26 ± 0.1 b | 0.001 |
| Final weight (g) | 2188 ± 18.4 c | 2143 ± 18.4 b | 2073 ± 18.4 a | 2055 ± 18.4 a | 0.028 |
| Average daily gain (g/d) | 109.3 ± 1.62 c | 105.7 ± 1.62 b | 100.8 ± 1.62 a | 101.7 ± 1.62 a | 0.043 |
| Amino Acid | Mw (g/mol) | Amino Acid/Lysine 1 | Molecules in the Sequence 2 |
|---|---|---|---|
| Alanine | 89.09 | 102 | 6 |
| Arginine | 174.2 | 105 | 7 |
| Asparagine | 132.1 | 56 | 4 |
| Aspartate | 133.1 | 66 | 4 |
| Cystein | 121.2 | 32 | 2 |
| Glutamate | 147.1 | 178 | 11 |
| Glutamine | 146.2 | 128 | 8 |
| Glycine | 75.1 | 176 | 11 |
| Histidine | 155.2 | 35 | 2 |
| Isoleucine | 131.2 | 67 | 4 |
| Leucine | 131.2 | 109 | 7 |
| Lysine | 146.2 | 100 | 6 |
| Methionine | 149.2 | 40 | 3 |
| Phenylalanine | 165.2 | 60 | 4 |
| Proline | 115.1 | 184 | 12 |
| Serine | 105.1 | 69 | 4 |
| Threonine | 119.1 | 67 | 4 |
| Tryptophan | 204.2 | 16 | 1 |
| Tyrosine | 181.2 | 45 | 3 |
| Valine | 117.2 | 77 | 5 |
| Total number of amino acids | 108 | ||
| Mw (g/mol) 3 | 12095.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cambra-López, M.; Marín-García, P.J.; Lledó, C.; Cerisuelo, A.; Pascual, J.J. Biomarkers and De Novo Protein Design Can Improve Precise Amino Acid Nutrition in Broilers. Animals 2022, 12, 935. https://doi.org/10.3390/ani12070935
Cambra-López M, Marín-García PJ, Lledó C, Cerisuelo A, Pascual JJ. Biomarkers and De Novo Protein Design Can Improve Precise Amino Acid Nutrition in Broilers. Animals. 2022; 12(7):935. https://doi.org/10.3390/ani12070935
Chicago/Turabian StyleCambra-López, María, Pablo Jesús Marín-García, Clara Lledó, Alba Cerisuelo, and Juan José Pascual. 2022. "Biomarkers and De Novo Protein Design Can Improve Precise Amino Acid Nutrition in Broilers" Animals 12, no. 7: 935. https://doi.org/10.3390/ani12070935