
Myostatin Knockout Limits Exercise-Induced Reduction in Bovine Erythrocyte Oxidative Stress by Enhancing the Efficiency of the Pentose Phosphate Pathway

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Animals
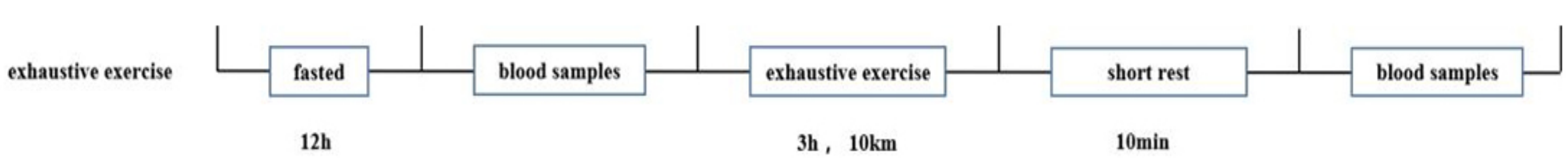
2.3. Experimental Design
2.4. Blood Sample Preparation
2.5. Antioxidant and Metabolite Detection Analysis
2.6. Fatty Acid (FA) Methyl Ester Analysis
2.7. Western Blotting
2.8. MSTN Protein Was Detected by ELISA
2.9. Immunofluorescence Staining
2.10. Calculations and Statistical Analyses
3. Results
3.1. Effect of Exhaustive Exercise on Bovine Hematological Parameters
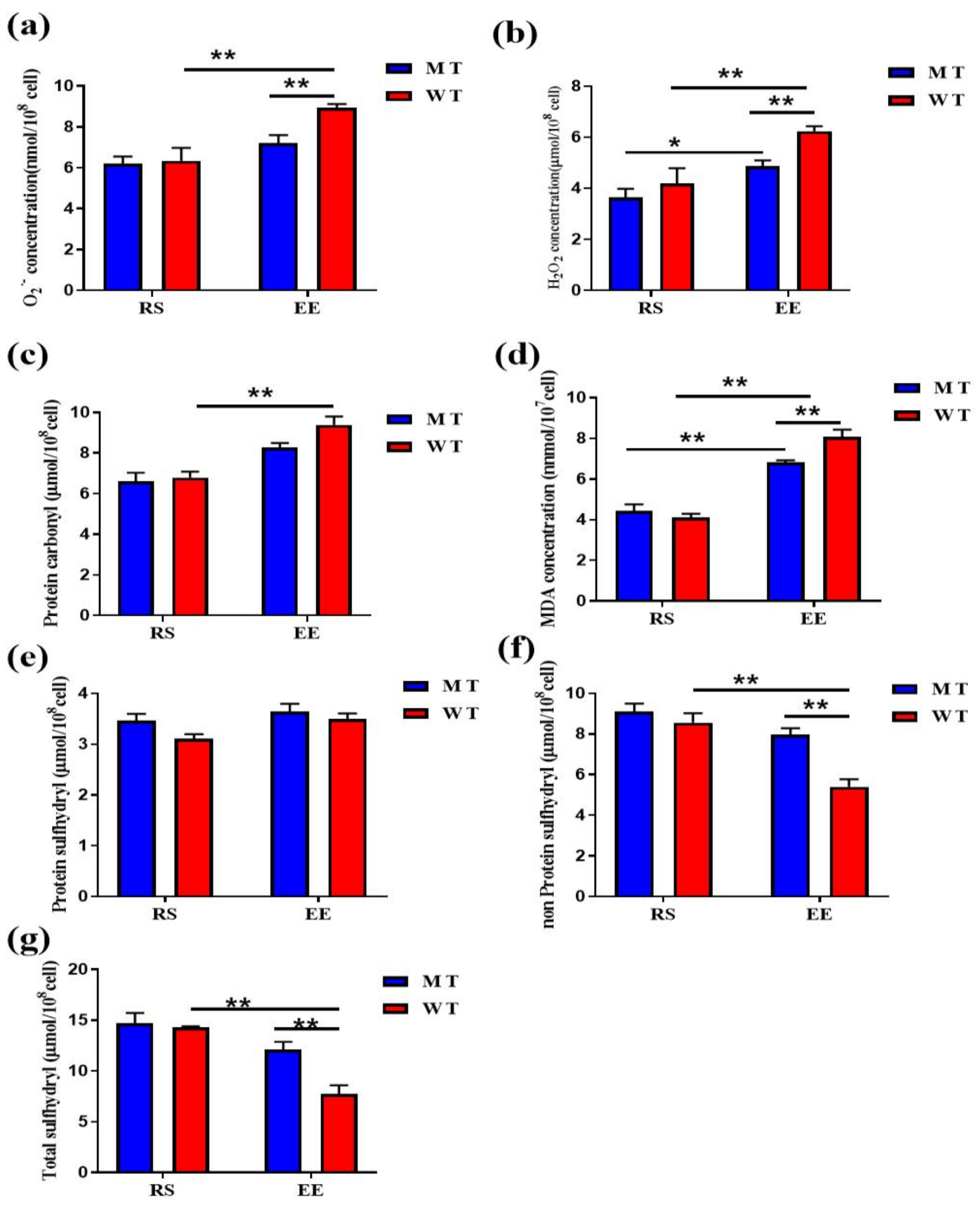
3.2. MT Erythrocytes Exhibit Less Oxidative Damage after Exhaustive Exercise
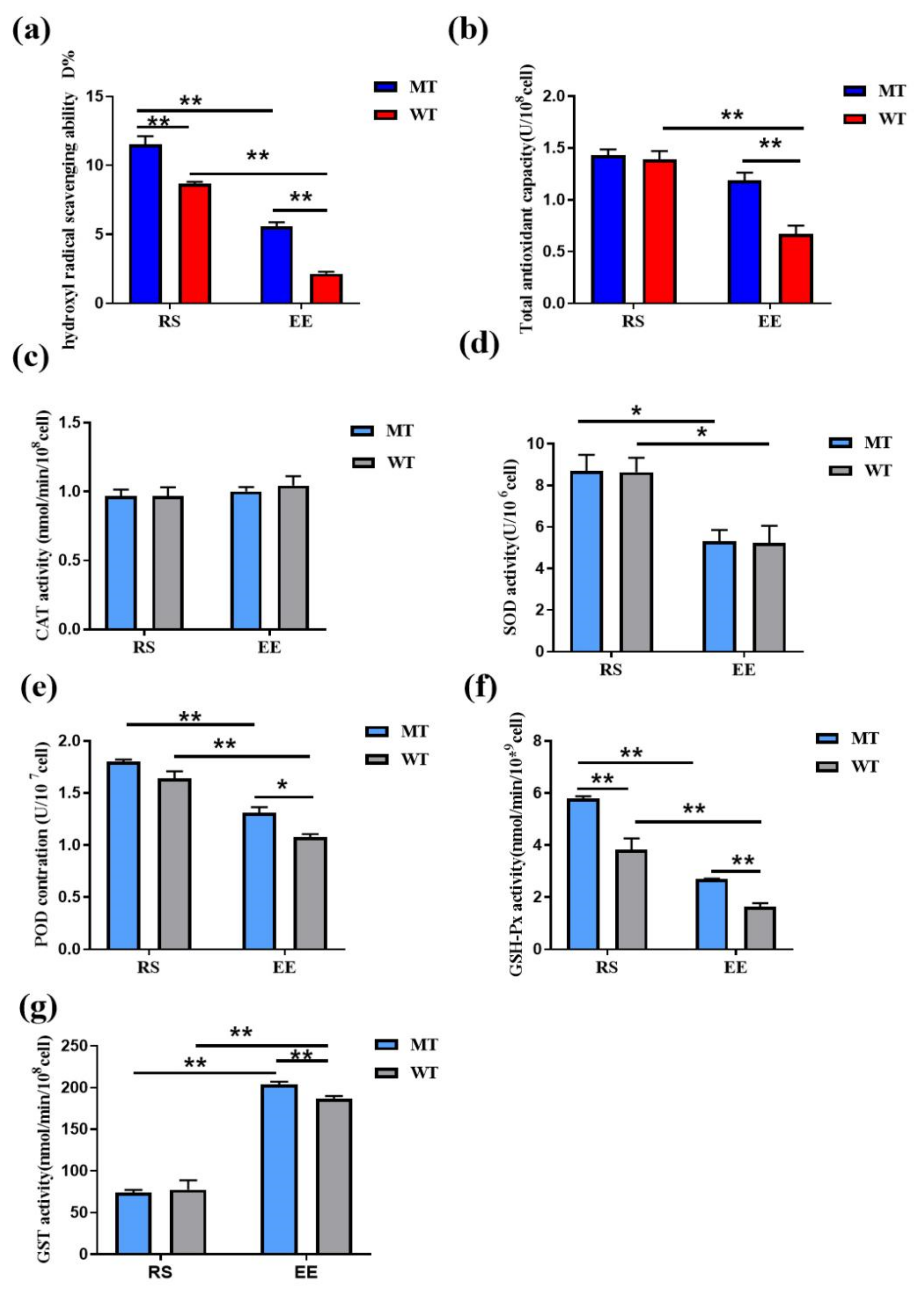
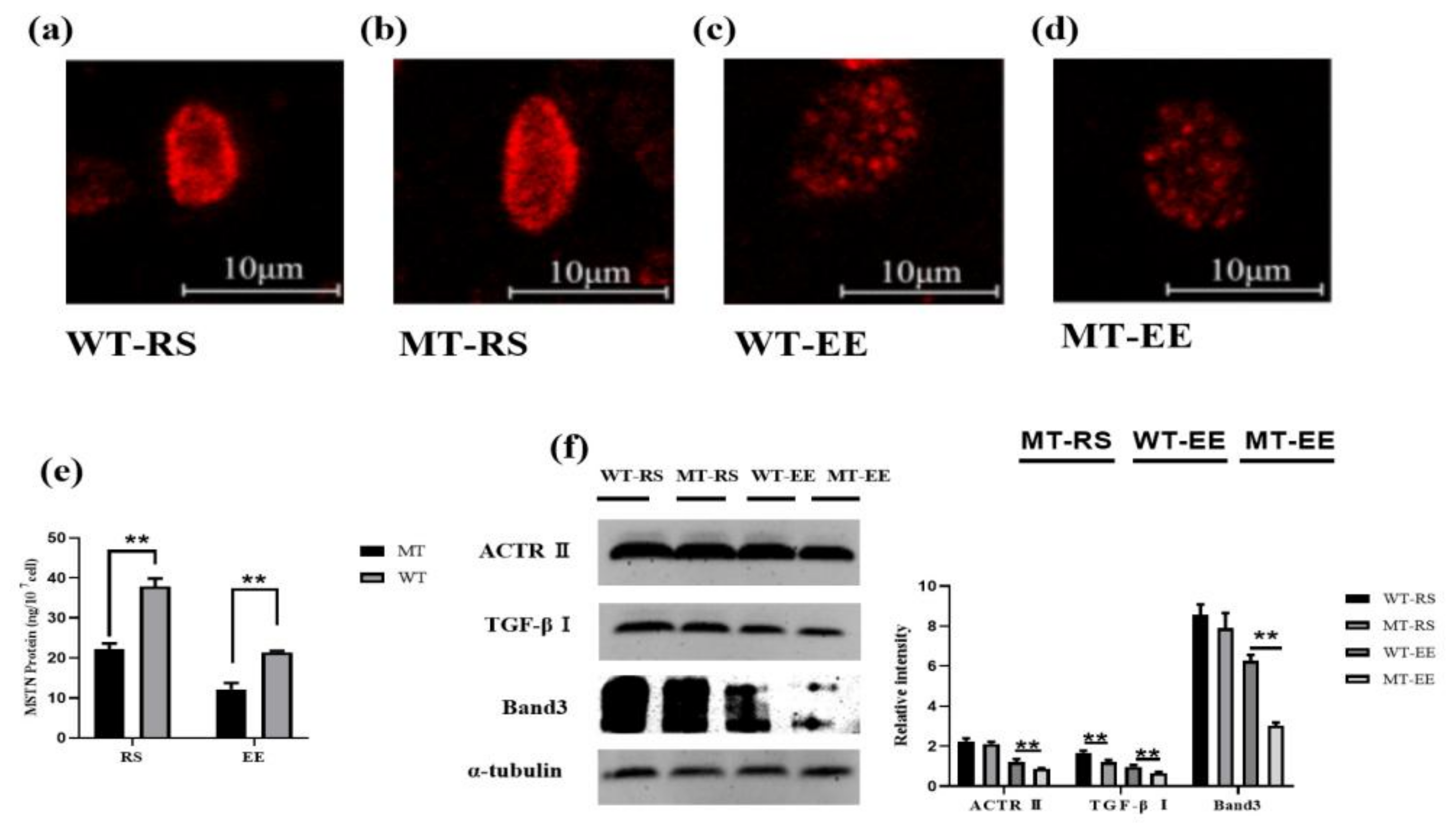
3.3. MSTN Knockout Increases the Erythrocyte Antioxidant Capacity
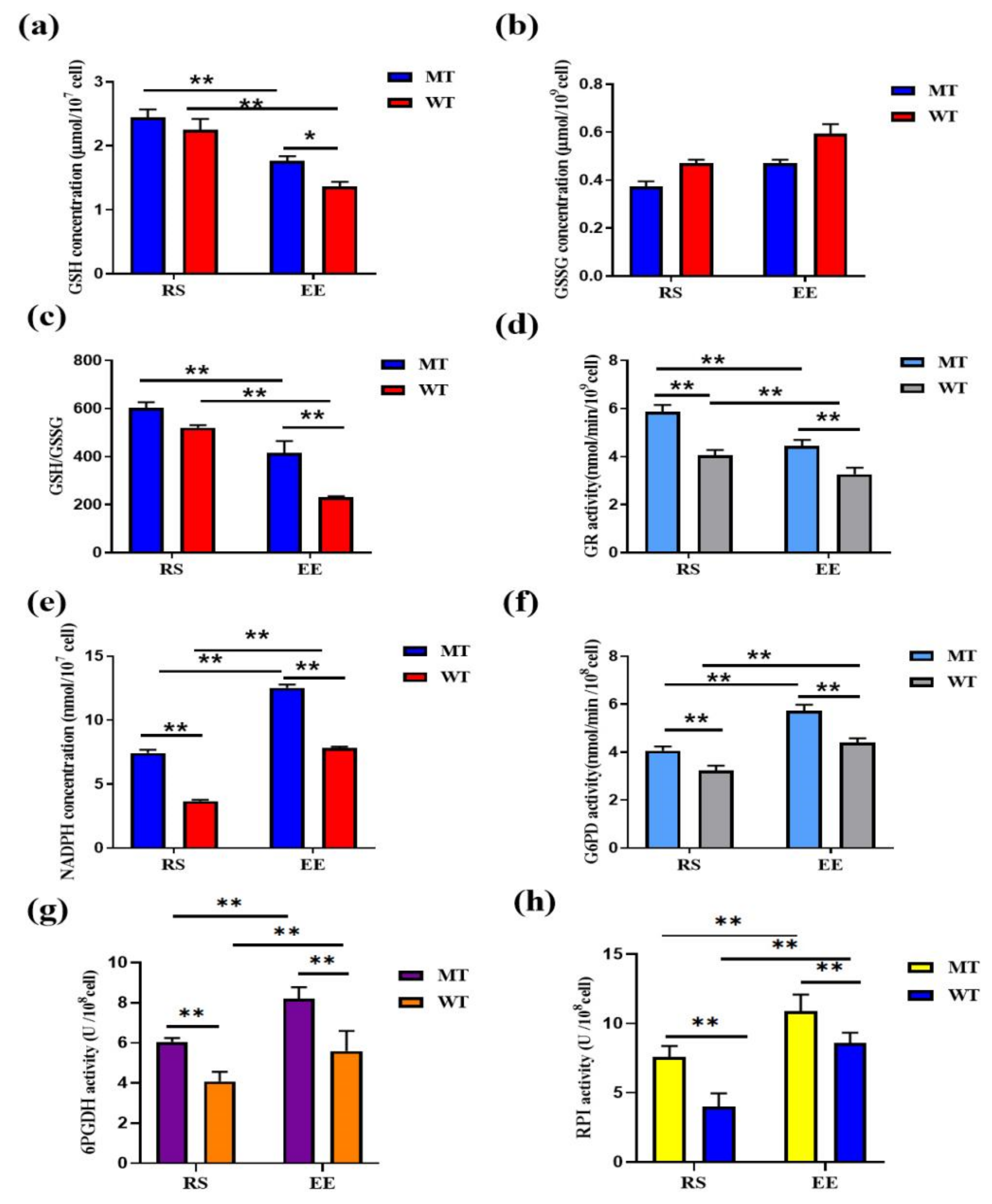
3.4. MSTN Knockout Increases Erythrocyte Antioxidant Capacity via GSH
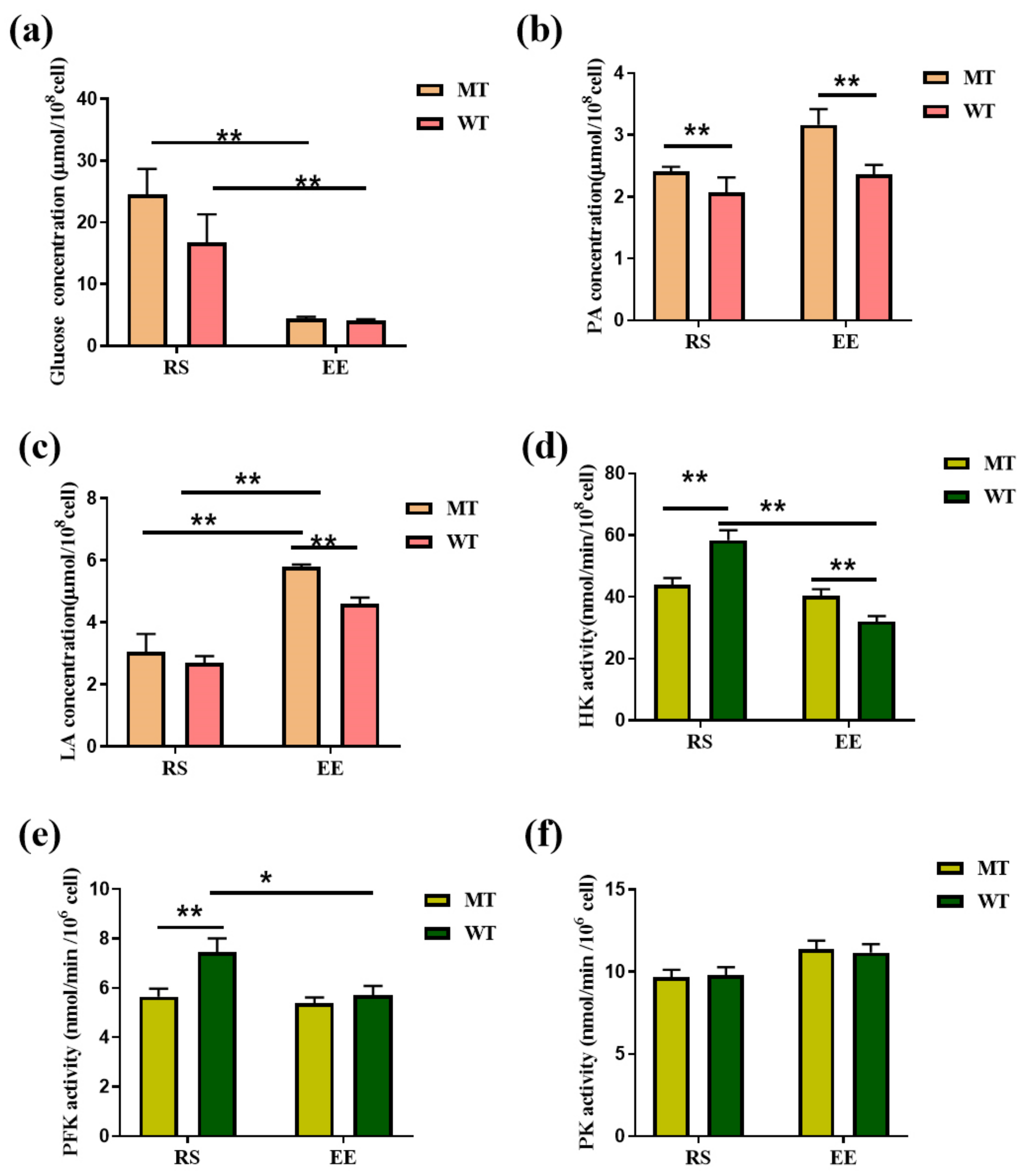
3.5. MSTN Knockout Affects Erythrocyte Glycolysis
3.6. MSTN Knockout Affects Erythrocyte Band 3
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jensen, F.B. The dual roles of red blood cells in tissue oxygen delivery: Oxygen carriers and regulators of local blood flow. J. Exp. Biol. 2009, 212, 3387–3393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Busija, D.W.; Gaspar, T.; Domoki, F.; Katakam, P.; Bari, F. Mitochondrial-mediated suppression of ROS production upon exposure of neurons to lethal stress: Mitochondrial targeted preconditioning. Adv. Drug Deliv. Rev. 2008, 60, 1471–1477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drummond, G.R.; Selemidis, S.; Griendling, K.K.; Sobey, C.G. Combating oxidative stress in vascular disease: NADPH oxidases as therapeutic targets. Nat. Rev. Drug Discov. 2011, 10, 453–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeo, J.H.; Lam, Y.W.; Fraser, S.T. Cellular dynamics of mammalian red blood cell production in the erythroblastic island niche. Biophys. Rev. 2019, 11, 873–894. [Google Scholar] [CrossRef]
- Van Wijk, R.; Van Solinge, W. The energy-less red blood cell is lost: Erythrocyte enzyme abnormalities of glycolysis. Blood 2005, 106, 4034–4042. [Google Scholar] [CrossRef]
- Johnson, R.M.; Goyette, G., Jr.; Ravindranath, Y.; Ho, Y.-S. Hemoglobin autoxidation and regulation of endogenous H2O2 levels in erythrocytes. Free Radic. Biol. Med. 2005, 39, 1407–1417. [Google Scholar] [CrossRef]
- Mohanty, J.G.; Nagababu, E.; Rifkind, J.M. Red blood cell oxidative stress impairs oxygen delivery and induces red blood cell aging. Front. Physiol. 2014, 5, 84. [Google Scholar] [CrossRef] [Green Version]
- Morabito, R.; Remigante, A.; Marino, A. Melatonin Protects Band 3 Protein in Human Erythrocytes against H2O2-Induced Oxidative Stress. Molecules 2019, 24, 2741. [Google Scholar] [CrossRef] [Green Version]
- Aoshiba, K.; Nakajima, Y.; Yasui, S.; Tamaoki, J.; Nagai, A. Red blood cells inhibit apoptosis of human neutrophils. Blood 1999, 93, 4006–4010. [Google Scholar] [CrossRef]
- Xiong, Y.; Xiong, Y.; Zhou, S.; Yu, Z.; Zhao, D.; Wang, Z.; Li, Y.; Yan, J.; Cai, Y.; Zhang, W. Inhibition of Glutathione Synthesis Induced by Exhaustive Running Exercise via the Decreased Influx Rate of L-Cysteine in Rat Erythrocytes. Cell. Physiol. Biochem. 2016, 40, 1410–1421. [Google Scholar] [CrossRef]
- Taverne, Y.J.H.J.; Bogers, A.J.J.C.; Duncker, D.J.; Merkus, D. Reactive Oxygen Species and the Cardiovascular System. Oxidative Med. Cell. Longev. 2013, 2013, 862423. [Google Scholar] [CrossRef] [PubMed]
- Tsantes, A.E.; Bonovas, S.; Travlou, A.; Sitaras, N.M. Redox Imbalance, Macrocytosis, and RBC Homeostasis. Antioxid. Redox Signal. 2006, 8, 1205–1216. [Google Scholar] [CrossRef] [PubMed]
- Corrado, D.; Zorzi, A. Sudden death in athletes. Int. J. Cardiol. 2017, 237, 67–70. [Google Scholar] [CrossRef] [PubMed]
- Maron, B.J.; Pelliccia, A. The Heart of Trained Athletes. Circulation 2006, 114, 1633–1644. [Google Scholar] [CrossRef] [Green Version]
- Popovic, L.; Mitic, N.R.; Radic, I.; Miric, D.; Kisic, B.; Krdzic, B.; Djokic, T. The effect of exhaustive exercise on oxidative stress generation and antioxidant defense in guinea pigs. Adv. Clin. Exp. Med. 2012, 21, 313–320. [Google Scholar]
- Yavari, A.; Javadi, M.; Mirmiran, P.; Bahadoran, Z. Exercise-Induced Oxidative Stress and Dietary antioxidants. Asian J. Sports Med. 2015, 6, e24898. [Google Scholar] [CrossRef] [Green Version]
- Xiong, Y.; Li, Y.; Xiong, Y.; Zhao, Y.; Tang, F.; Wang, X. Cluster of erythrocyte band 3: A potential molecular target of exhaustive exercise-induced dysfunction of erythrocyte deformability. Can. J. Physiol. Pharmacol. 2013, 91, 1127–1134. [Google Scholar] [CrossRef]
- Berzosa, C.; Gómez–Trullén, E.M.; Piedrafita, E.; Cebrián, I.; Martínez–Ballarín, E.; Miana–Mena, F.J.; Fuentes–Broto, L.; García, J.J. Erythrocyte membrane fluidity and indices of plasmatic oxidative damage after acute physical exercise in humans. Eur. J. Appl. Physiol. 2010, 111, 1127–1133. [Google Scholar] [CrossRef]
- Brzeszczynska, J.; Pieniazek, A.; Gwozdzinski, L.; Gwozdzinski, K.; Jegier, A. Structural alterations of erythrocyte membrane components induced by exhaustive exercise. Appl. Physiol. Nutr. Metab. 2008, 33, 1223–1231. [Google Scholar] [CrossRef]
- Aguiló, A.; Tauler, P.; Fuentespina, E.; Tur, J.A.; Córdova, A.; Pons, A. Antioxidant response to oxidative stress induced by exhaustive exercise. Physiol. Behav. 2005, 84, 1–7. [Google Scholar] [CrossRef]
- Veskoukis, A.S.; Nikolaidis, M.G.; Kyparos, A.; Kokkinos, D.; Nepka, C.; Barbanis, S.; Kouretas, D. Effects of xanthine oxidase inhibition on oxidative stress and swimming performance in rats. Appl. Physiol. Nutr. Metab. 2008, 33, 1140–1154. [Google Scholar] [CrossRef] [PubMed]
- Jain, M.; Cui, L.; Brenner, D.A.; Wang, B.; Handy, D.E.; Leopold, J.A.; Loscalzo, J.; Apstein, C.S.; Liao, R. Increased Myocardial Dysfunction After Ischemia-Reperfusion in Mice Lacking Glucose-6-Phosphate Dehydrogenase. Circulation 2004, 109, 898–903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Méndez, D.; Linares, M.; Diez, A.; Puyet, A.; Bautista, J.M. Stress response and cytoskeletal proteins involved in erythrocyte membrane remodeling upon Plasmodium falciparum invasion are differentially carbonylated in G6PD A− deficiency. Free Radic. Biol. Med. 2011, 50, 1305–1313. [Google Scholar] [CrossRef] [PubMed]
- Cheah, F.; Peskin, A.V.; Wong, F.; Ithnin, A.; Othman, A.; Winterbourn, C.C. Increased basal oxidation of peroxiredoxin 2 and limited peroxiredoxin recycling in glucose-6-phosphate dehydrogenase-deficient erythrocytes from newborn infants. FASEB J. 2014, 28, 3205–3210. [Google Scholar] [CrossRef]
- Aiello, D.; Patel, K.; Lasagna, E. Themyostatingene: An overview of mechanisms of action and its relevance to livestock animals. Anim. Genet. 2018, 49, 505–519. [Google Scholar] [CrossRef] [Green Version]
- Zhu, P.; Li, H.; Huang, G.; Cui, J.; Zhang, R.; Cui, K.; Yang, S.; Shi, D. Molecular Cloning, Identification, and Expression Patterns of Myostatin Gene in Water Buffalo (Bubalus Bubalis). Anim. Biotechnol. 2017, 29, 26–33. [Google Scholar] [CrossRef]
- Kong, X.; Yao, T.; Zhou, P.; Kazak, L.; Tenen, D.; Lyubetskaya, A.; Dawes, B.A.; Tsai, L.; Kahn, B.B.; Spiegelman, B.M.; et al. Brown Adipose Tissue Controls Skeletal Muscle Function via the Secretion of Myostatin. Cell Metab. 2018, 28, 631–643. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.-H.; Bauman, W.A.; Cardozo, C.P. Myostatin inhibits glucose uptake via suppression of insulin-dependent and -independent signaling pathways in myoblasts. Physiol. Rep. 2018, 6, e13837. [Google Scholar] [CrossRef]
- Camporez, J.-P.G.; Petersen, M.C.; Abudukadier, A.; Moreira, G.V.; Jurczak, M.J.; Friedman, G.; Haqq, C.M.; Petersen, K.F.; Shulman, G.I. Anti-myostatin antibody increases muscle mass and strength and improves insulin sensitivity in old mice. Proc. Natl. Acad. Sci. USA 2016, 113, 2212–2217. [Google Scholar] [CrossRef] [Green Version]
- Biesemann, N.; Mendler, L.; Wietelmann, A.; Hermann, S.; Schäfers, M.; Krüger, M.; Boettger, T.; Borchardt, T.; Braun, T. Myostatin Regulates Energy Homeostasis in the Heart and Prevents Heart Failure. Circ. Res. 2014, 115, 296–310. [Google Scholar] [CrossRef] [Green Version]
- Wu, D.; Gu, M.; Wei, Z.; Bai, C.; Su, G.; Liu, X.; Zhao, Y.; Yang, L.; Li, G. Myostatin Knockout Regulates Bile Acid Metabolism by Promoting Bile Acid Synthesis in Cattle. Animals 2022, 12, 205. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Bai, C.; We, Z.; Su, G.; Wu, Y.; Han, L.; Yang, L.; Liu, X.; Zhao, Y.; Song, E.; et al. Myostatin gene editing in yellow cattle. J. Inn. Mong. Univ. (Natl. Sci. Ed.) 2020, 51, 12–32. [Google Scholar] [CrossRef]
- Williams, C.A. Horse species symposium: The effect of oxidative stress during exercise in the horse. J. Anim. Sci. 2016, 94, 4067–4075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maśko, M.; Domino, M.; Jasiński, T.; Witkowska-Piłaszewicz, O. The Physical Activity-Dependent Hematological and Biochemical Changes in School Horses in Comparison to Blood Profiles in Endurance and Race Horses. Animals 2021, 11, 1128. [Google Scholar] [CrossRef] [PubMed]
- Bottegaro, N.B.; Gotić, J.; Šuran, J.; Brozić, D.; Klobučar, K.; Bojanić, K.; Vrbanac, Z. Effect of prolonged submaximal exercise on serum oxidative stress biomarkers (d-ROMs, MDA, BAP) and oxidative stress index in endurance horses. BMC Vet. Res. 2018, 14, 216. [Google Scholar] [CrossRef] [Green Version]
- Mostafiz, M.M.; Jhan, P.K.; Shim, J.-K.; Lee, K.Y. Methyl benzoate exhibits insecticidal and repellent activities against Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae). PLoS ONE 2018, 13, e0208552. [Google Scholar] [CrossRef]
- Baggio, B.; Priante, G.; Brunati, A.M.; Clari, G.; Bordin, L. Specific modulatory effect of arachidonic acid on human red blood cell oxalate transport: Clinical implications in calcium oxalate nephrolithiasis. J. Am. Soc. Nephrol. JASN 1999, 10 (Suppl. 14), S381–S384. [Google Scholar]
- Calder, P.C. Polyunsaturated fatty acids, inflammatory processes and inflammatory bowel diseases. Mol. Nutr. Food Res. 2008, 52, 885–897. [Google Scholar] [CrossRef]
- Ghashghaeinia, M.; Giustarini, D.; Koralkova, P.; Köberle, M.; Alzoubi, K.; Bissinger, R.; Hosseinzadeh, Z.; Dreischer, P.; Bernhardt, I.; Lang, F.; et al. Pharmacological targeting of glucose-6-phosphate dehydrogenase in human erythrocytes by Bay 11–7082, parthenolide and dimethyl fumarate. Sci. Rep. 2016, 6, 28754. [Google Scholar] [CrossRef] [Green Version]
- Van den Akker, E.; Satchwell, T.J.; Williamson, R.C.; Toye, A.M. Band 3 multiprotein complexes in the red cell membrane; of mice and men. Blood Cells, Mol. Dis. 2010, 45, 1–8. [Google Scholar] [CrossRef]
- Hsu, K. Exploring the Potential Roles of Band 3 and Aquaporin-1 in Blood CO2 Transport–Inspired by Comparative Studies of Glycophorin B-A-B Hybrid Protein GP.Mur. Front. Physiol. 2018, 9, 733. [Google Scholar] [CrossRef] [PubMed]
- Lewis, I.A.; Campanella, M.E.; Markley, J.L.; Low, P.S. Role of band 3 in regulating metabolic flux of red blood cells. Proc. Natl. Acad. Sci. USA 2009, 106, 18515–18520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimo, H.; Arjunan, S.N.V.; Machiyama, H.; Nishino, T.; Suematsu, M.; Fujita, H.; Tomita, M.; Takahashi, K. Particle Simulation of Oxidation Induced Band 3 Clustering in Human Erythrocytes. PLoS Comput. Biol. 2015, 11, e1004210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakraborty, I.; Mishra, R.; Gachhui, R.; Kar, M. Distortion of β-globin Chain of Hemoglobin Alters the Pathway of Erythrocytic Glucose Metabolism Through Band 3 Protein. Arch. Med Res. 2012, 43, 112–116. [Google Scholar] [CrossRef]
- Chu, H.; Low, P.S. Mapping of glycolytic enzyme-binding sites on human erythrocyte band 3. Biochem. J. 2006, 400, 143–151. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MT-RS | WT-RS | MT-EE | WT-EE | |
|---|---|---|---|---|
| RBC (10^12/L) | 7.72 ± 0.77 | 7.82 ± 0.43 | 7.87 ± 0.75 | 7.48 ± 0.46 |
| RDW (fL) | 10 ± 1.0 | 10.0 ± 1.04 | 9.67 ± 1.03 | 9.58 ± 1.08 |
| HCT (%) | 3403 ± 299 | 3388 ± 140 | 3584 ± 236 | 3506 ± 249 |
| MCV (fL) | 43.19 ± 2.2 | 45.7 ± 3.1 | 43.3 ± 2.2 | 45.2 ± 3.0 |
| MCH (Pg) | 14.71 ± 0.79 | 14.51 ± 0.93 | 14.78 ± 0.85 | 14.42 ± 1.04 |
| MCHC (g/L) | 31.78 ± 0.67 | 32.07 ± 0.27 | 31.5 ± 0.55 | 31.93 ± 0.47 |
| HGB (g/dl) | 111.2 ± 8.63 | 110.88 ± 6.79 | 109.46 ± 11.46 | 107.72 ± 10.59 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, L.; Bai, C.; Wang, X.; Wei, Z.; Gu, M.; Zhou, X.; Su, G.; Liu, X.; Yang, L.; Li, G. Myostatin Knockout Limits Exercise-Induced Reduction in Bovine Erythrocyte Oxidative Stress by Enhancing the Efficiency of the Pentose Phosphate Pathway. Animals 2022, 12, 927. https://doi.org/10.3390/ani12070927
Zhu L, Bai C, Wang X, Wei Z, Gu M, Zhou X, Su G, Liu X, Yang L, Li G. Myostatin Knockout Limits Exercise-Induced Reduction in Bovine Erythrocyte Oxidative Stress by Enhancing the Efficiency of the Pentose Phosphate Pathway. Animals. 2022; 12(7):927. https://doi.org/10.3390/ani12070927
Chicago/Turabian StyleZhu, Lin, Chunling Bai, Xueqiao Wang, Zhuying Wei, Mingjuan Gu, Xinyu Zhou, Guanghua Su, Xuefei Liu, Lei Yang, and Guangpeng Li. 2022. "Myostatin Knockout Limits Exercise-Induced Reduction in Bovine Erythrocyte Oxidative Stress by Enhancing the Efficiency of the Pentose Phosphate Pathway" Animals 12, no. 7: 927. https://doi.org/10.3390/ani12070927