
Proteomic Analysis of Intracellular and Membrane-Associated Fractions of Canine (Canis lupus familiaris) Epididymal Spermatozoa and Sperm Structure Separation

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Media
2.2. Animals
2.3. Cauda Epididymal Semen Collection
2.4. Spermatozoa Quality Assessment
2.5. Sample Preparation for the Sonication
2.6. Morphology Assessment of Epididymal Spermatozoa Using Spermac Stain
2.7. Total Protein Content Measurement
2.8. Polyacrylamide Gel Electrophoresis (SDS-PAGE)
2.9. Identification of Proteins by Mass Spectrometry
2.9.1. In-Gel Trypsin Digestion
2.9.2. NanoUPLC-Q-TOF/MS Analysis
2.10. Gene Ontology and Functional Annotation
2.11. Statistical Analysis
3. Results
3.1. Sperm Motility Assessment
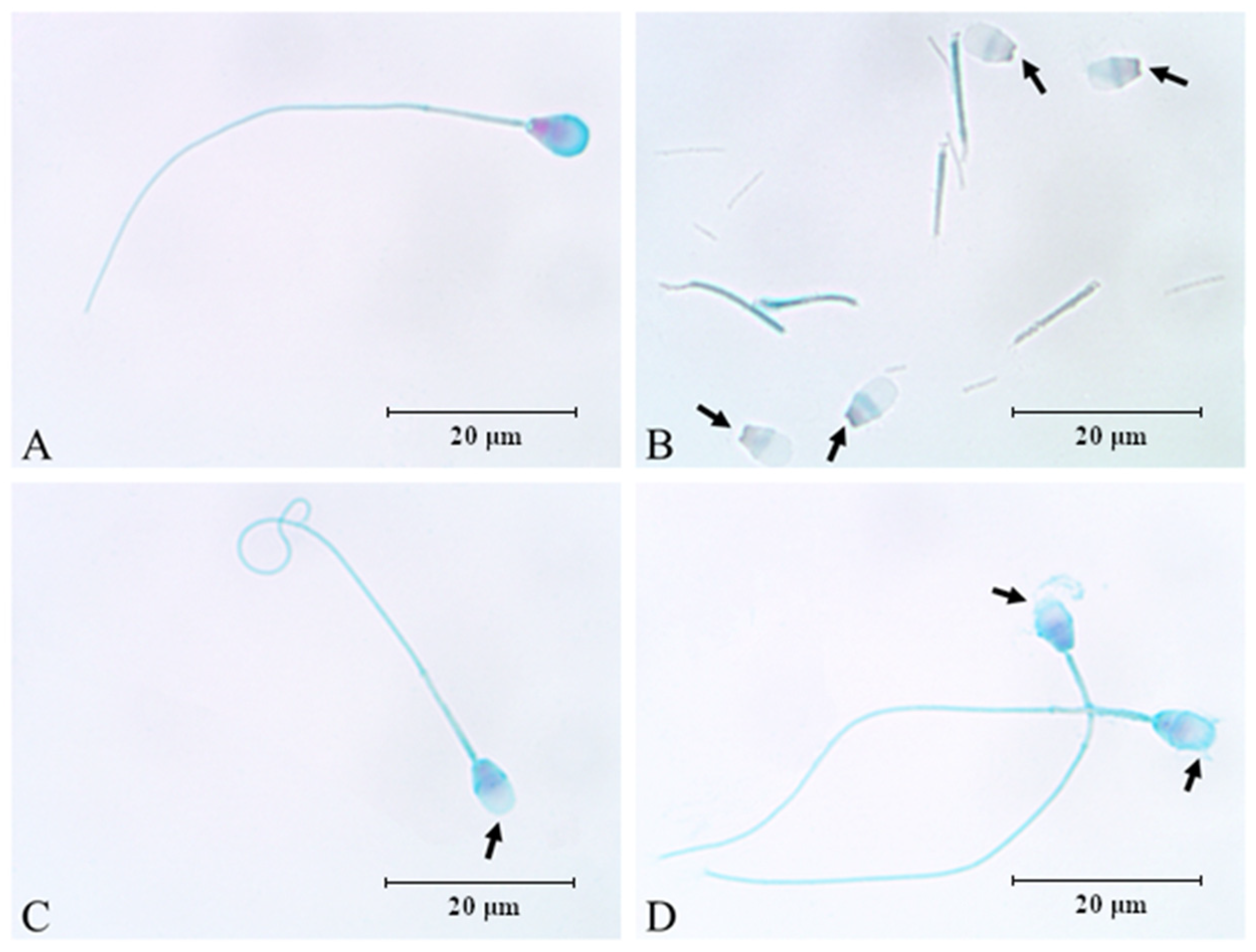
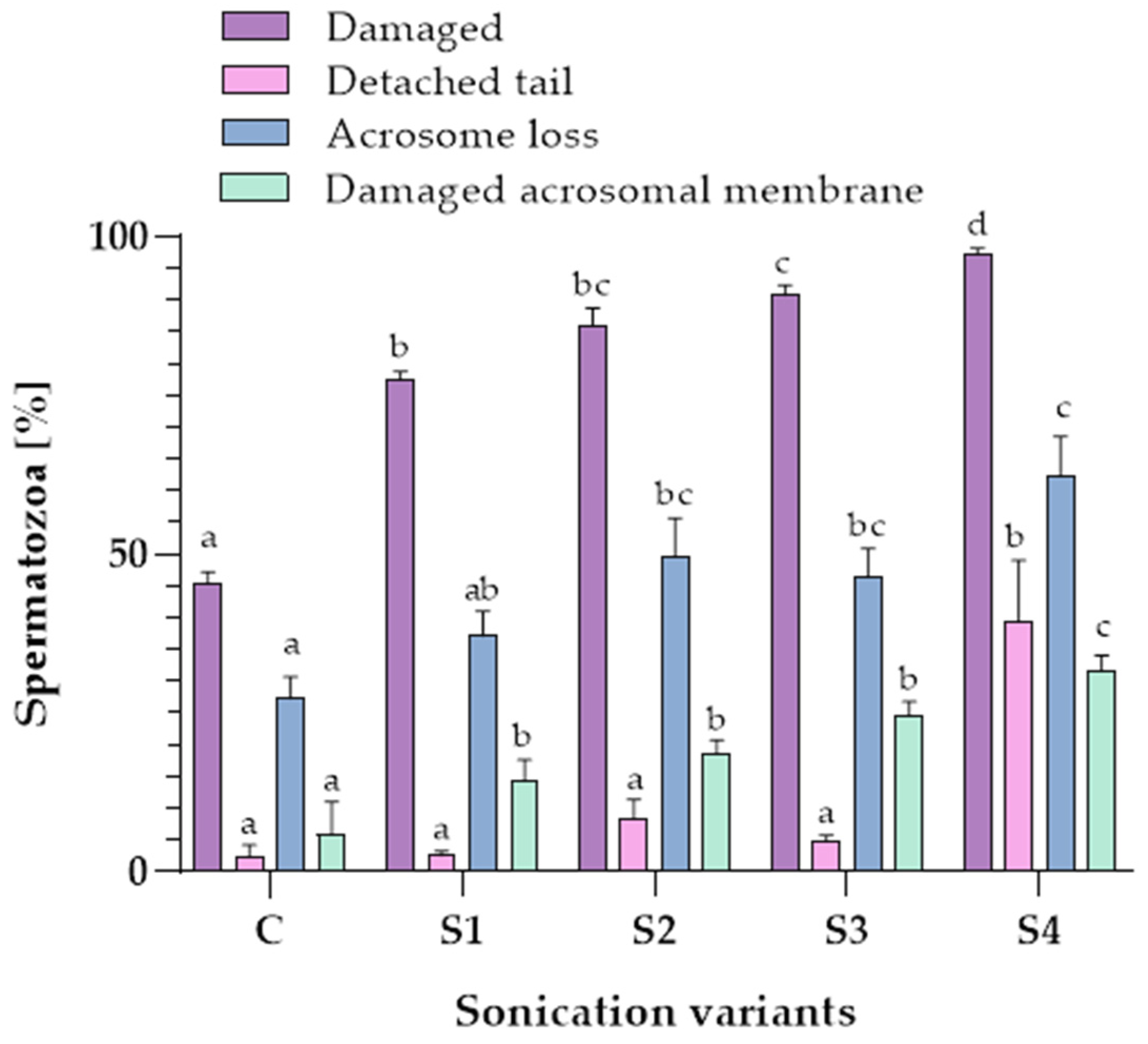
3.2. Influence of Sonication on Epididymal Spermatozoa Morphology
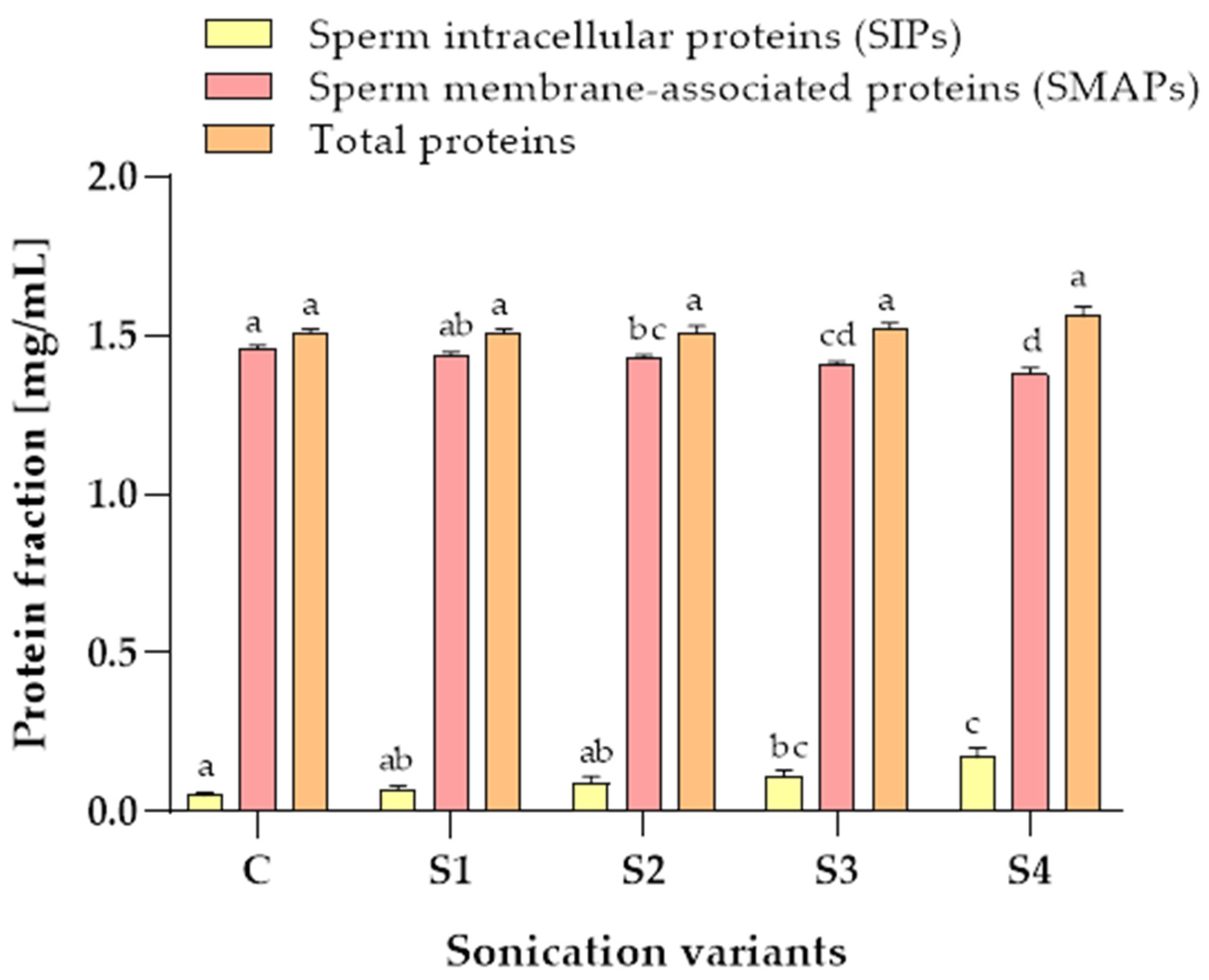
3.3. Protein Content in Samples after the Sonication
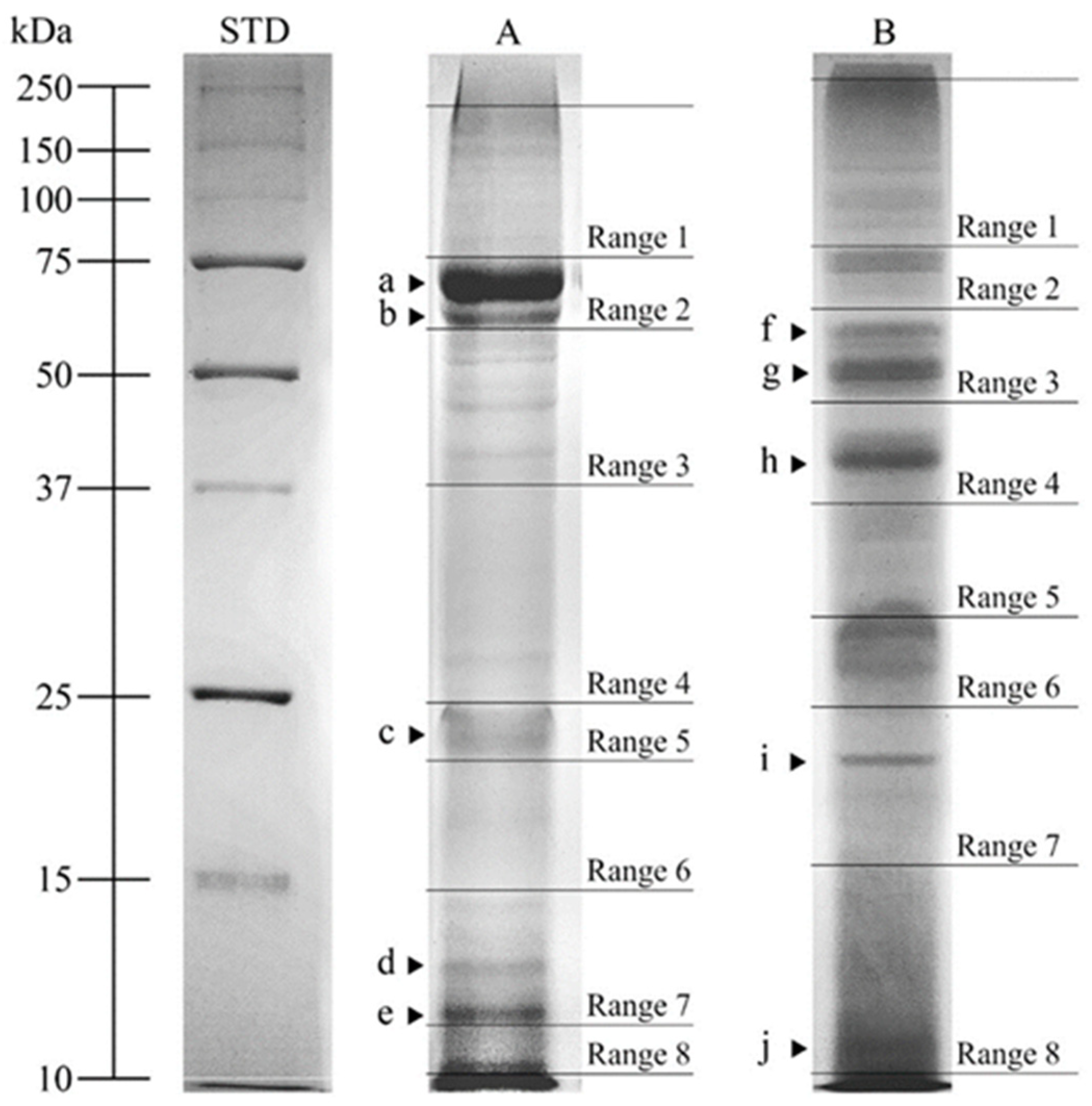
3.4. SDS-PAGE Analysis
3.5. Mass Spectrometry Analysis
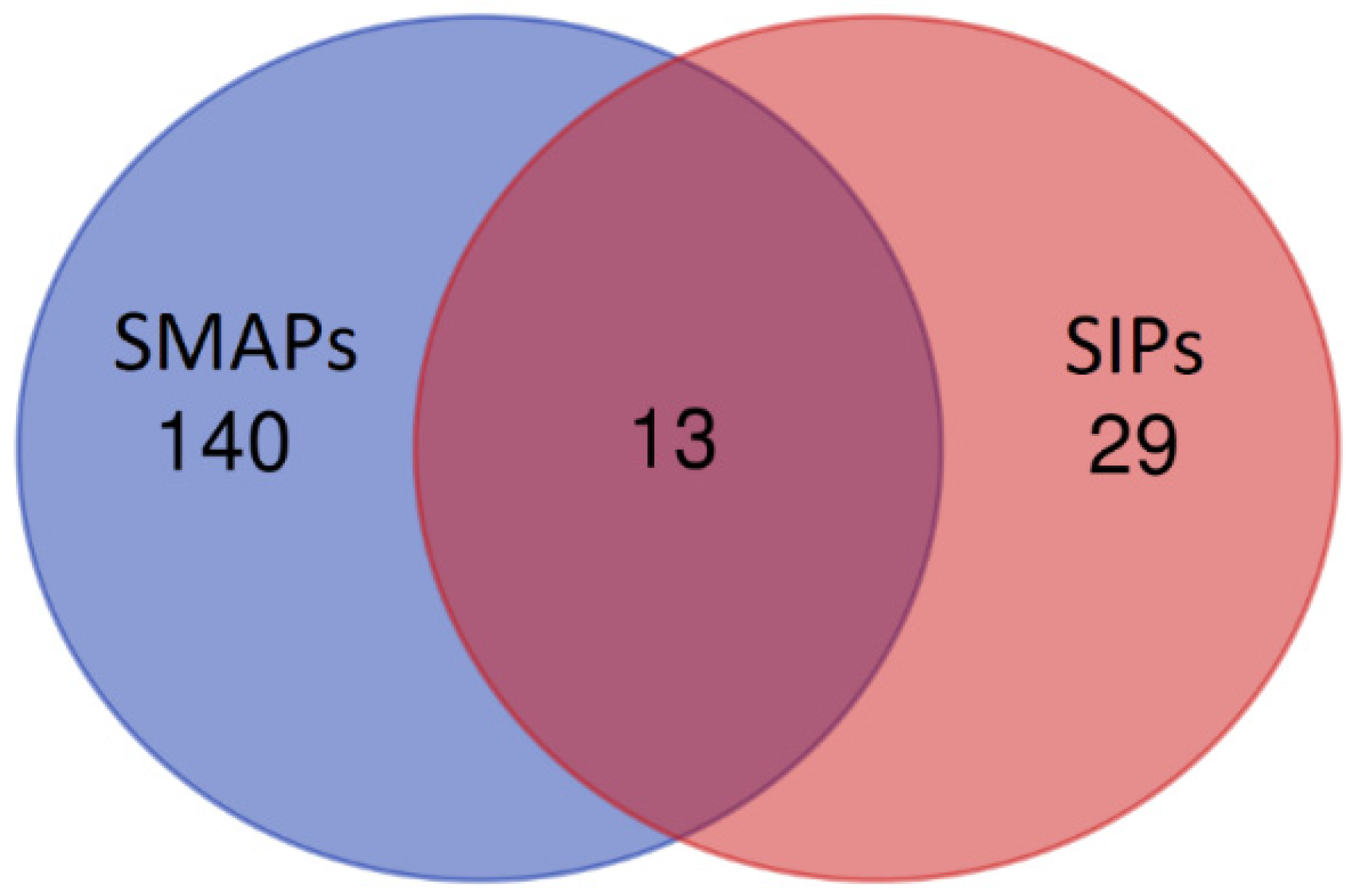
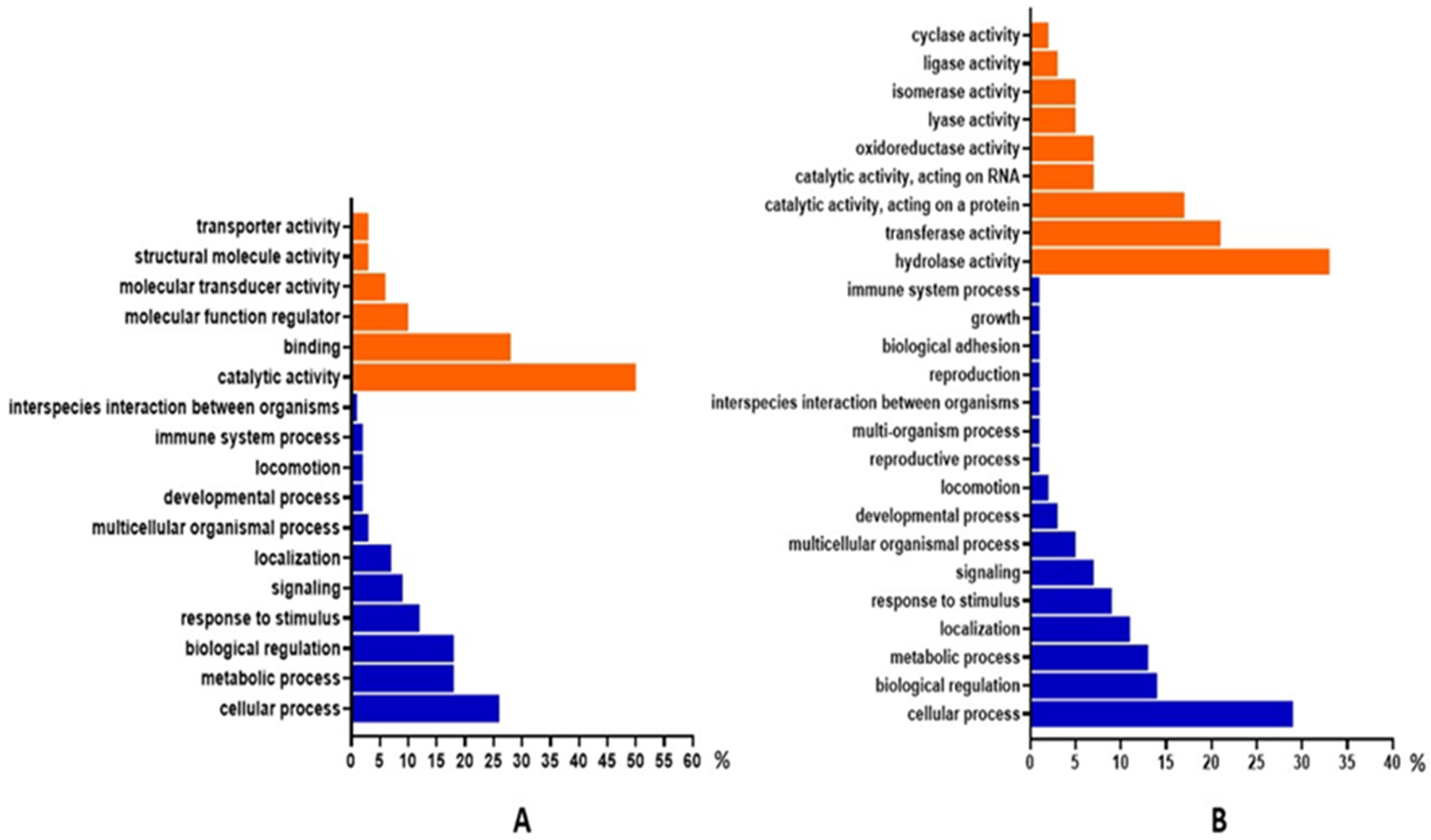
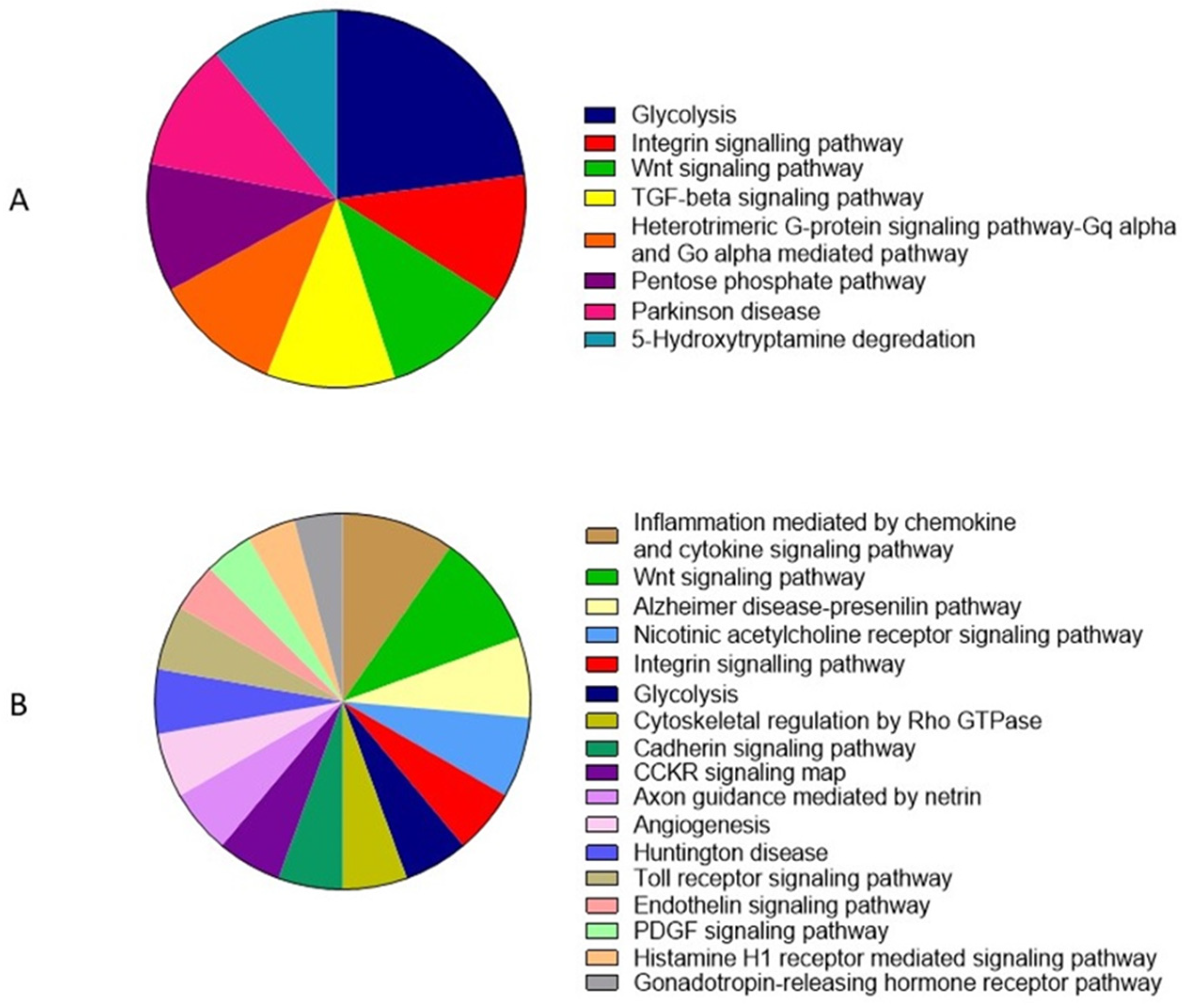
3.6. Gene Ontology and Functional Annotation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Luvoni, G.C.; Morselli, M.G. Canine epididymal spermatozoa: A hidden treasure with great potential. Reprod. Dom. Anim. 2016, 52, 197–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassan, H.A.; Domain, G.; Luvoni, G.C.; Chaaya, R.; Van Soom, A.; Wydooghe, E. Canine and Feline Epididymal Semen—A Plentiful Source of Gametes. Animals 2021, 11, 2961. [Google Scholar] [CrossRef] [PubMed]
- Varesi, S.; Varnocchi, V.; Faustini, M.; Luvoni, G.C. Morphological and acrosomal changes of canine spermatozoa during epididymal transit. Acta Vet. Scand. 2013, 55, 17. [Google Scholar] [CrossRef] [Green Version]
- Chaveiro, A.; Cerqueira, C.; Silva, J.; Franco, J.; Moreira da Silva, F. Evaluation of frozen thawed cauda epididymal sperms and in vitro fertilizing potential of bovine sperm collected from the cauda epididymal. Iran. J. Vet. Res. 2015, 16, 188–193. [Google Scholar] [PubMed]
- Kirchhoff, C. The dog as model to study human epididymal function at a molecular level. Mol. Hum. Reprod. 2002, 8, 695–701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Switonski, M. Dog as a model in studies on human hereditary diseases and their gene therapy. Reprod. Biol. 2014, 14, 44–50. [Google Scholar] [CrossRef]
- Toyoshima, M.; Sakata, M.; Ohnishi, K.; Tokumaru, Y.; Kato, Y.; Tokutsu, R.; Sakamoto, W.; Minagawa, J.; Matsuda, F.; Shimizu, H. Targeted proteome analysis of microalgae under high-light conditions by optimized protein extraction of photosynthetic organisms. J. Biosci. Bioeng. 2019, 127, 394–402. [Google Scholar] [CrossRef]
- Qin, P.; Han, T.; Yu, A.C.H.; Xu, L. Mechanistic understanding the bioeffects of ultrasound-driven microbubbles to enhance macromolecule delivery. J. Control. Release 2018, 272, 169–181. [Google Scholar] [CrossRef]
- Tateno, H.; Kimura, Y.; Yanagimachi, R. Sonication per se is not as deleterious to sperm chromosomes as previously inferred. Biol. Reprod. 2000, 63, 341–346. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, T.; Yoneyama, M.; Imanishi, M.; Takeuchi, M. Flow cytometric detection and analysis of tailless sperm caused by sonication or a chemical agent. J. Toxicol. Sci. 2000, 25, 41–48. [Google Scholar] [CrossRef]
- Somanath, P.R.; Gandhi, K.K. Isolation and partial characterisation of the plasma and outer acrosomal membranes of goat spermatozoa. Small Rumin. Res. 2004, 53, 67–74. [Google Scholar] [CrossRef]
- Amaral, A.; Castillo, J.; Estanyol, J.M.; Ballesca, J.L.; Ramalho-Santos, J.; Oliva, R. Human sperm tail proteome suggests new endogenous metabolic pathways. Mol. Cell. Proteom. 2013, 12, 330–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dias, G.M.; López, M.L.; Ferreira, A.T.S.; Chapeaurouge, D.A.; Rodrigues, A.; Perales, J.; Retamal, C.A. Thiol-disulfide proteins of stallion epididymal spermatozoa. Anim. Reprod. Sci. 2014, 145, 29–39. [Google Scholar] [CrossRef] [Green Version]
- Fanny, J.; Julien, S.; Francisco-Jose, F.G.; Sabiha, E.; Sophie, D.D.; Luc, B.; Hélène, B.; Nicolas, S.; Valérie, M. Gel electrophoresis of human sperm: A simple method for evaluating sperm protein quality. Basic Clin. Androl. 2018, 28, 10. [Google Scholar] [CrossRef]
- Guasti, P.N.; Souza, F.F.; Scott, C.; Papa, P.M.; Camargo, L.S.; Schmith, R.A.; Monteiro, G.A.; Hartwig, F.P.; Papa, F.O. Equine seminal plasma and sperm membrane: Functional proteomic assessment. Theriogenology 2020, 156, 70–81. [Google Scholar] [CrossRef]
- Baker, S.S.; Cardullo, R.A.; Thaler, C.D. Sonication of mouse sperm membranes reveals distinct protein domains. Biol. Reprod. 2002, 66, 57–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mann, T.; Lutwak-Mann, C. Secretory function of male accessory organs of reproduction in mammals. Physiol. Rev. 1951, 31, 27–55. [Google Scholar] [CrossRef]
- Mogielnicka-Brzozowska, M.; Kordan, W. Characteristics of selected seminal plasma proteins and their application in the improvement of the reproductive processes in mammals. Pol. J. Vet. Sci. 2011, 14, 489–499. [Google Scholar] [CrossRef]
- Panner Selvam, M.K.; Finelli, R.; Agarwal, A.; Henkel, R. Proteomics and metabolomics—Current and future perspectives in clinical andrology. Andrologia 2021, 53, e13711. [Google Scholar] [CrossRef]
- Gobello, C.; Castex, G.; Corrada, Y. Serum and seminal markers in the diagnosis of disorders of the genital tract of the dog: A mini-review. Theriogenology 2002, 57, 1285–1291. [Google Scholar] [CrossRef]
- Jobim, M.I.M.; Trein, C.; Zirkler, H.; Gregory, R.M.; Sieme, H.; Mattos, R.C. Two-dimensional polyacrylamide gel electrophoresis of equine seminal plasma proteins and their relation with semen freezability. Theriogenology 2011, 76, 765–771. [Google Scholar] [CrossRef] [PubMed]
- Schafer-Somi, S.; Palme, N. Seminal plasma characteristics and expression of ATP-binding cassette transporter A1 (ABCA1) in canine spermatozoa from ejaculates with good and bad freezability. Reprod. Dom. Anim. 2016, 21, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Aquino-Cortez, A.; Pinheiro, B.Q.; Lima, D.B.C.; Silva, H.V.R.; Mota-Filho, A.C.; Martins, J.A.M.; Rodriguez-Villamil, P.; Moura, A.A.; Silva, L.D.M. Proteomic characterization of canine seminal plasma. Theriogenology 2017, 95, 178–186. [Google Scholar] [CrossRef] [PubMed]
- Araujo, M.S.; Oliveira Henriques Paulo, O.L.D.; Paranzini, C.S.; Scott, C.; Codognoto, V.M.; de Paula Freitas Dell’Aqua, C.; Papa, F.O.; de Souza, F.F. Proteomic data of seminal plasma and spermatozoa of four purebred dogs. Data Brief 2020, 30, 105498. [Google Scholar] [CrossRef] [PubMed]
- Araujo, M.S.; Oliveira Henriques Paulo, O.L.D.; Scott, C.; Paranzini, C.S.; Codognoto, V.M.; de Paula Freitas Dell’Aqua, C.; Papa, F.O.; de Souza, F.F. Insights into the influence of canine breed on proteomics of the spermatozoa and seminal plasma. J. Proteom. 2022, 257, 104508. [Google Scholar] [CrossRef]
- Blash, S.; Melican, D.; Gavin, W. Cryopreservation of epididymal sperm obtained at necropsy from goats. Theriogenology 2000, 54, 899–905. [Google Scholar] [CrossRef]
- Mogielnicka-Brzozowska, M.; Prochowska, S.; Niżański, W.; Bromke, M.A.; Wiśniewski, J.; Olejnik, B.; Kuzborska, A.; Fraser, L.; Młynarz, P.; Kordan, W. Proteome of cat semen obtained after urethral catheterization. Theriogenology 2020, 141, 68–81. [Google Scholar] [CrossRef]
- Cebi, C.; Faundez, R.; Jurka, P.; Akcay, E.; Petrajtis-Golobow, M.; Ambarcioglu, P. Evaluation of the canine epididymal sperm morphology with two different staining methods, one fixative solution and Motile Sperm Organelle Morphology Examination (MSOME). Kafkas Univ. Vet. Fak. Derg. 2016, 22, 57–62. [Google Scholar] [CrossRef]
- Intasqui, P.; Agarwal, A.; Sharma, R.; Samanta, L.; Bertolla, R.P. Towards the identification of reliable sperm biomarkers for male infertility: A sperm proteomic approach. Andrologia 2018, 50, e12919. [Google Scholar] [CrossRef]
- World Health Organization. WHO Laboratory Manual for the Examination and Processing of Human Semen, 5th ed.; WHO Press: Geneva, Switzerland, 2010; ISBN 978-92-4-154778-9. [Google Scholar]
- Shevchenko, A.; Tomas, H.; Havli, J.; Olsen, J.V.; Mann, M. In-gel digestion for mass spectrometric characterization of proteins and proteomes. Nat. Protoc. 2006, 1, 2856–2860. [Google Scholar] [CrossRef]
- Yoon, S.J.; Rahman, M.S.; Kwon, W.S.; Ryu, D.Y.; Park, Y.J.; Pang, M.G. Proteomic identification of cryostress in epididymal spermatozoa. J. Anim. Sci. Biotechnol. 2016, 7, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labas, V.; Spina, L.; Belleannee, C.; Teixeira-Gomes, A.P.; Gargaros, A.; Dacheux, F.; Dacheux, J.L. Analysis of epididymal sperm maturation by MALDI profiling and top-down mass spectrometry. J. Proteom. 2015, 15, 226–243. [Google Scholar] [CrossRef] [PubMed]
- Chauvin, T.; Xie, F.; Liu, T.; Nicora, C.D.; Yang, F.; Camp, D.G.; Smith, R.D.; Roberts, K.P. A systematic analysis of a deep mouse epididymal sperm proteome. Biol. Reprod. 2012, 87, 141. [Google Scholar] [CrossRef] [PubMed]
- Björkgren, I.; Sipilä, P. The impact of epididymal proteins on sperm function. Reproduction 2019, 158, 155–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pearl, C. Immunolocalization of Lactoferrin in the Rodent Epididymis. Biol. Reprod. 2010, 83, 516. [Google Scholar] [CrossRef]
- Armstrong, J.S.; Rajasekaran, M.; Hellstrom, W.J.; Sikka, S.C. Antioxidant potential of human serum albumin: Role in the recovery of high quality human spermatozoa for assisted reproductive technology. J. Androl. 1998, 19, 412–419. [Google Scholar]
- Arroteia, K.F.; Barbieri, M.F.; Souza, G.H.M.F.; Tanaka, H.; Eberlin, M.N.; Hyslop, S.; Alvares, L.E.; Pereira, L.A.V.D. Albumin Is Synthesized in Epididymis and Aggregates in a High Molecular Mass Glycoprotein Complex Involved in Sperm-Egg Fertilization. PLoS ONE 2014, 9, e103566. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Rodgers, G.P. Olfactomedin 4 expression and functions in innate immunity, inflammation, and cancer. Cancer Metastasis Rev. 2016, 35, 201–212. [Google Scholar] [CrossRef]
- Kodithuwakku, S.P.; Pang, R.T.K.; Ng, E.H.Y.; Cheung, A.N.Y.; Horne, A.W.; Ho, P.-C.; Yeung, W.S.B.; Lee, K.-F. Wnt activation downregulates olfactomedin-1 in Fallopian tubal epithelial cells: A microenvironment predisposed to tubal ectopic pregnancy. Lab. Investig. 2012, 92, 256–264. [Google Scholar] [CrossRef]
- Li, H.; Liu, W.; Chen, W.; Zhu, J.; Deng, C.X.; Rodgers, G.P. Olfactomedin 4 deficiency promotes prostate neoplastic progression and is associated with upregulation of the hedgehog-signaling pathway. Sci. Rep. 2015, 5, 16974. [Google Scholar] [CrossRef] [Green Version]
- Sharma, R.; Agarwal, A.; Mohanty, G.; Hamada, A.J.; Gopalan, B.; Willard, B.; Yadav, S.; du Plessis, S. Proteomic analysis of human spermatozoa proteins with oxidative stress. Reprod. Biol. Endocrinol. 2013, 11, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, K.P.; Finn, G.; Lee, T.H.; Nicholson, L.K. Prolyl cis–trans isomerization as a molecular timer. Nat. Chem. Biol. 2007, 3, 619–629. [Google Scholar] [CrossRef] [PubMed]
- Nigro, P.; Pompilio, G.; Capogrossi, M.C. Cyclophilin A: A key player for human disease. Cell Death Dis. 2013, 4, e888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westfalewicz, B.; Dietrich, M.; Słowińska, M.; Judycka, S.; Ciereszko, A. Seasonal changes in the proteome of cryopreserved bull semen supernatant. Theriogenology 2019, 126, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Pini, T.; Parks, J.; Russ, J.; Dzieciatkowska, M.; Hansen, K.C.; Schoolcraft, W.B.; Katz-Jaffe, M. Obesity significantly alters the human sperm proteome, with potential implications for fertility. J. Assist. Reprod. Genet. 2020, 37, 777–787. [Google Scholar] [CrossRef]
- Carvalho, M.G.; Silva, K.M.; Aristizabal, V.H.V.; Ortiz, P.E.O.; Paranzini, C.S.; Melchert, A.; Amaro, J.L.; Souza, F.F. Effects of Obesity and Diabetes on Sperm Cell Proteomics in Rats. J. Proteome Res. 2021, 20, 2628–2642. [Google Scholar] [CrossRef]
- Xue, J.-C.; Goldberg, E. Identification of a Novel Testis-Specific Leucine-Rich Protein in Humans and Mice. Biol. Reprod. 2000, 62, 1278–1284. [Google Scholar] [CrossRef] [Green Version]
- Alvarez-Rodriguez, M.; Martinez, C.; Wright, D.; Barranco, I.; Roca, J.; Rodriguez-Martinez, H. The Transcriptome of Pig Spermatozoa, and Its Role in Fertility. Int. J. Mol. Sci. 2020, 21, 1572. [Google Scholar] [CrossRef] [Green Version]
- Fukamachi, K.; Matsuoka, Y.; Ohno, H.; Hamaguchi, T.; Tsuda, H. Neuronal leucine-rich repeat protein-3 amplifies MAPK activation by epidermal growth factor through a carboxyl-terminal region containing endocytosis motifs. J. Biol. Chem. 2002, 277, 43549–43552. [Google Scholar] [CrossRef] [Green Version]
- Foster, J.A.; Gerton, G.L. Autoantigen 1 of the guinea pig sperm acrosome is the homologue of mouse Tpx-1 and human TPX1 and is a member of the cysteine-rich secretory protein (CRISP) family. Mol. Reprod. Dev. 1996, 44, 221–229. [Google Scholar] [CrossRef]
- O’Bryan, M.K.; Loveland, K.L.; Herszfeld, D.; McFarlane, J.R.; Hearn, M.T.; de Kretser, D.M. Identification of a rat testis-specific gene encoding a potential rat outer dense fibre protein. Mol. Reprod. Dev. 1998, 50, 313–322. [Google Scholar] [CrossRef]
- Reddy, T.; Gibbs, G.M.; Merriner, D.J.; Kerr, J.B.; O’Bryan, M.K. Cysteine-rich secretory proteins are not exclusively expressed in the male reproductive tract. Dev. Dyn. 2008, 237, 3313–3323. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.; Kierzek, M.; O’Connor, A.E.; Brenker, C.; Merriner, D.J.; Okuda, H.; Volpert, M.; Gaikwad, A.; Bianco, D.; Potter, D.; et al. CRISP2 Is a Regulator of Multiple Aspects of Sperm Function and Male Fertility. Endocrinology 2019, 160, 915–924. [Google Scholar] [CrossRef] [PubMed]
- Maeda, T.; Sakashita, M.; Ohba, Y.; Nakanishi, Y. Molecular cloning of the rat Tpx-1 responsible for the interaction between spermatogenic and Sertoli cells. Biochem. Biophys. Res. Commun. 1998, 248, 140–146. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, V.; Schneiter, R. Pathogen-related yeast (PRY) proteins and members of the CAP superfamily are secreted sterol-binding proteins. Proc. Natl. Acad. Sci. USA 2012, 109, 16882–16887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibbs, G.M.; Scanlon, M.J.; Swarbrick, J.; Curtis, S.; Gallant, E.; Dulhunty, A.F.; O’Bryan, M.K. The cysteine-rich secretory protein domain of Tpx-1 is related to ion channel toxins and regulates ryanodine receptor Ca2+ signaling. J. Biol. Chem. 2006, 281, 4156–4163. [Google Scholar] [CrossRef] [Green Version]
- Guo, M.; Teng, M.; Niu, L.; Liu, Q.; Huang, Q.; Hao, Q. Crystal structure of the cysteine-rich secretory protein stecrisp reveals that the cysteine-rich domain has a K+ channel inhibitor-like fold. J. Biol. Chem. 2005, 280, 12405–12412. [Google Scholar] [CrossRef] [Green Version]
- Jamsai, D.; Reilly, A.; Smith, S.J.; Gibbs, G.M.; Baker, H.W.; McLachlan, R.I.; de Kretser, D.M.; O’Bryan, M.K. Polymorphisms in the human cysteine-rich secretory protein 2 (CRISP2) gene in Australian men. Hum. Reprod. 2008, 23, 2151–2159. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.H.; Zhou, Q.Z.; Lyu, X.M.; Zhu, T.; Chen, Z.J.; Chen, M.K.; Xia, H.; Wang, C.Y.; Qi, T.; Li, X.; et al. The expression of cysteine-rich secretory protein 2 (CRISP2) and its specific regulator miR-27b in the spermatozoa of patients with asthenozoospermia. Biol. Reprod. 2015, 92, 28. [Google Scholar] [CrossRef]
- Gottschalk, M.; Metzger, J.; Martinsson, G.; Sieme, H.; Distl, O. Genome-wide association study for semen quality traits in German Warmblood stallions. Anim. Reprod. Sci. 2016, 171, 81–86. [Google Scholar] [CrossRef]
- Pedersen, S.F.; Hoffmann, E.K.; Mills, J.W. The cytoskeleton and cell volume regulation. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2001, 130, 385–399. [Google Scholar] [CrossRef]
- Cabello-Agüeros, J.F.; Hernández-González, E.O.; Mújica, A. The role of F-actin cytoskeleton-associated gelsolin in the guinea pig capacitation and acrosome reaction. Cell Motil. Cytoskelet. 2003, 56, 94–108. [Google Scholar] [CrossRef] [PubMed]
- Breitbart, H.; Cohen, G.; Rubinstein, S. Role of actin cytoskeleton in mammalian sperm capacitation and the acrosome reaction. Reproduction 2005, 129, 263–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naresh, S. Effect of cooling (4 °C) and cryopreservation on cytoskeleton actin and protein tyrosine phosphorylation in buffalo spermatozoa. Cryobiology 2016, 72, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Sutovsky, P.; Hauser, R.; Sutovsky, M. Increased levels of sperm ubiquitin correlate with semen quality in men from an andrology laboratory clinic population. Hum. Reprod. 2004, 19, 628–638. [Google Scholar] [CrossRef] [PubMed]
- Baska, K.M.; Manandhar, G.; Feng, D.; Agca, Y.; Tengowski, M.W.; Sutovsky, M.; Yi, Y.J.; Sutovsky, P. Mechanism of extracellular ubiquitination in the mammalian epididymis. J. Cell. Physiol. 2008, 215, 684–696. [Google Scholar] [CrossRef]
- Vernocchi, V.; Morselli, M.G.; Varesi, S.; Nonnis, S.; Maffioli, E.; Negri, A.; Tedeschi, G.; Luvoni, G.C. Sperm ubiquitination in epididymal feline semen. Theriogenology 2014, 82, 636–642. [Google Scholar] [CrossRef]
- Takada, M.; Yonezawa, N.; Yoshizawa, M.; Noguchi, S.; Hatanaka, Y.; Nagai, T.; Kikuchi, K.; Aoki, H.; Nakano, M. pH-Sensitive Dissociation and Association of β-N-Acetylhexosaminidase from Boar Sperm Acrosome. Biol. Reprod. 1994, 50, 860–868. [Google Scholar] [CrossRef]
- Miller, D.J.; Gong, X.; Shur, B.D. Sperm require beta-N-acetylglucosaminidase to penetrate through the egg zona pellucida. Development 1993, 118, 1279–1289. [Google Scholar] [CrossRef]
- Wysocki, P.; Orzołek, A.; Strzezek, J.; Koziorowska-Gilun, M.; Zasiadczyk, Ł.; Kordan, W. The activity of N-acetyl-b-hexosaminidase in boar seminal plasma is linked with semen quality and its suitability for cryopreservation. Theriogenology 2015, 83, 1194–1202. [Google Scholar] [CrossRef]
- Baba, T.; Michikawa, Y.; Kashiwabara, S.; Arai, Y. Proacrosin activation in the presence of a 32-kDa protein from boar spermatozoa. Biochem. Biophys. Res. Commun. 1989, 160, 1026–1032. [Google Scholar] [CrossRef]
- Baba, T.; Niida, Y.; Michikawa, Y.; Kashiwabara, S.; Kodaira, K.; Takenaka, M.; Kohno, N.; Gerton, G.L.; Arai, Y. An acrosomal protein, sp32, in mammalian sperm is a binding protein specific for two proacrosins and an acrosin intermediate. J. Biol. Chem. 1994, 269, 10133–10140. [Google Scholar] [CrossRef]
- Ono, T.; Kurashige, T.; Harada, N.; Noguchi, Y.; Saika, T.; Niikawa, N.; Aoe, M.; Nakamura, S.; Higashi, T.; Hiraki, A.; et al. Identification of proacrosin binding protein sp32 precursor as a human cancer/testis antigen. Proc. Natl. Acad. Sci. USA 2001, 98, 3282–3287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitehurst, A.W.; Xie, Y.; Purinton, S.C.; Cappell, K.M.; Swanik, J.T.; Larson, B.; Girard, L.; Schorge, J.O.; White, M.A. Tumor antigen acrosin binding protein normalizes mitotic spindle function to promote cancer cell proliferation. Cancer Res. 2010, 70, 7652–7661. [Google Scholar] [CrossRef] [Green Version]
- Kanemori, Y.; Koga, Y.; Sudo, M.; Kang, W.; Kashiwabara, S.; Ikawa, M.; Hasuwa, H.; Nagashima, K.; Ishikawa, Y.; Ogonuki, N.; et al. Biogenesis of sperm acrosome is regulated by pre-mRNA alternative splicing of Acrbp in the mouse. Proc. Natl. Acad. Sci. USA 2016, 113, 3696–3705. [Google Scholar] [CrossRef] [Green Version]
- Waheed, M.M.; Gouda, E.M.; Khalifa, T.A. Impact of seminal plasma superoxide dismutase and glutathione peroxidase on cryopreserved buffalo spermatozoa. Anim. Reprod. Sci. 2013, 142, 126–130. [Google Scholar] [CrossRef]
- Neagu, V.R.; García, B.M.; Rodríguez, A.M.; Ferrusola, C.O.; Bolaños, J.M.G.; Fernández, L.G.; Tapia, J.A.; Peña, F.J. Determination of glutation peroxidase and superoxide dismutase activities in canine seminal plasma and its relation with sperm quality and lipid peroxidation post thaw. Theriogenology 2011, 75, 10–16. [Google Scholar] [CrossRef]
- Barranco, I.; Tvarijonaviciute, A.; Perez-Patiño, C.; Vicente-Carrillo, A.; Parrilla, I.; Ceron, J.J.; Martinez, E.A.; Rodriguez-Martinez, H.; Roca, J. Glutathione Peroxidase 5 Is Expressed by the Entire Pig Male Genital Tract and Once in the Seminal Plasma Contributes to Sperm Survival and In Vivo Fertility. PLoS ONE 2016, 11, e0162958. [Google Scholar] [CrossRef]
- Tilburg, M.; Sousa, S.; Lobo, M.D.P.; Monteiro-Azevedo, A.C.O.M.; Azevedo, R.A.; Araújo, A.A.; Moura, A.A. Mapping the major proteome of reproductive fluids and sperm membranes of rams: From the cauda epididymis to ejaculation. Theriogenology 2021, 159, 98–107. [Google Scholar] [CrossRef]
- Zhou, Y.; Shaw, N.; Li, Y.; Zhao, Y.; Zhang, R.; Liu, Z.J. Structure-function analysis of human l-prostaglandin D synthase bound with fatty acid molecules. FASEB J. 2010, 24, 4668–4677. [Google Scholar] [CrossRef] [Green Version]
- Froman, D.P.; Amann, R.P.; Riek, P.M.; Olar, T.T. Acrosin activity of canine spermatozoa as an index of cellular damage. J. Reprod. Fert. 1984, 70, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Pera, I.; Ivell, R.; Kirchhoff, C. Regional variation of gene expression in the dog epididymis as revealed by in-situ transcript hybridization. Int. J. Androl. 1994, 17, 324–330. [Google Scholar] [CrossRef] [PubMed]
- Naureckiene, S.; Sleat, D.E.; Lackland, H.; Fensom, A.; Vanier, M.T.; Wattiaux, R.; Jadot, M.; Lobel, P. Identification of HE1 as the second gene of Niemann-Pick C disease. Science 2000, 290, 2298–2301. [Google Scholar] [CrossRef]
- Osterhoff, C.; Kirchhoff, C.; Krull, N.; Ivell, R. Molecular cloning and characterization of a novel human sperm antigen (HE2) specifically expressed in the proximal epididymis. Biol. Reprod. 1994, 50, 516–525. [Google Scholar] [CrossRef] [Green Version]
- Ru, Y.F.; Xue, H.M.; Ni, Z.M.; Xia, D.; Zhou, Y.C.; Zhang, Y.L. An epididymis-specific carboxyl esterase CES5A is required for sperm capacitation and male fertility in the rat. Asian J. Androl. 2015, 17, 292–297. [Google Scholar] [CrossRef]
- Zhang, L.; Hu, Z.; Zhu, C.; Liu, Q.; Zhou, Y.; Zhang, Y. Identification and characterization of an epididymis-specific gene, Ces7. Acta Biochim. Biophys. Sin. 2009, 41, 809–815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, L.; Yang, Z.; Wang, L.-J.; Yan, H.-Y.; Zhang, Y.; Han, X.-Y.; Yao, Y.-P.; Dang, X.; Zhang, Y.-H.; Guo, X.-M.; et al. Expression of the Ces5a gene in the rat testis. Zhonghua Nan Ke Xue 2019, 25, 867–873. [Google Scholar]
- Salier, J.P. Chromosomal location, exon/intron organization and evolution of lipocalin genes. Biochim. Biophys. Acta 2000, 1482, 25–34. [Google Scholar] [CrossRef]
- Suzuki, K.; Yu, X.; Chaurand, P.; Araki, Y.; Lareyre, J.-J.; Caprioli, R.M.; Orgebin-Crist, M.-C.; Matusik, R.J. Epididymis-specific lipocalin promoters. Asian J. Androl. 2007, 9, 515–521. [Google Scholar] [CrossRef]
- Thimon, V.; Koukoui, O.; Calvo, E.; Sullivan, R. Region-specific gene expression profiling along the human epididymis. Mol. Hum. Reprod. 2007, 13, 691–704. [Google Scholar] [CrossRef]
- Wu, C.; Wang, C.; Zhai, B.; Zhao, Y.; Zhao, Z.; Yuan, Z.; Fu, X.; Zhang, M. Study on the region-specific expression of epididymis mRNA in the rams. PLoS ONE 2021, 16, e0245933. [Google Scholar] [CrossRef] [PubMed]
- Ding, G.; Wang, J.; Feng, C.; Jiang, H.; Xu, J.; Ding, Q. Lipocalin 2 over-expression facilitates progress of castration-resistant prostate cancer via improving androgen receptor transcriptional activity. Oncotarget 2016, 7, 64309–64317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindblad-Toh, K.; Wade, C.M.; Mikkelsen, T.S.; Karlsson, E.K.; Jaffe, D.B.; Kamal, M.; Clamp, M.; Chang, J.L.; Kulbokas, E.J., III; Zody, M.C.; et al. Genome sequence, comparative analysis and haplotype structure of the domestic dog. Nature 2005, 438, 803–819. [Google Scholar] [CrossRef] [PubMed]
- Hereng, T.H.; Elgstoen, K.B.P.; Cederkvist, F.H.; Eide, L.; Jahnsen, T.; Skalhegg, B.S.; Rosendal, K.R. Exogenous pyruvate accelerates glycolysis and promotes capacitation in human spermatozoa. Hum. Reprod. 2011, 26, 3249–3263. [Google Scholar] [CrossRef] [Green Version]
- Merc, V.; Frolikova, M.; Komrskova, K. Role of Integrins in Sperm Activation and Fertilization. Int. J. Mol. Sci. 2021, 22, 11809. [Google Scholar] [CrossRef]
- Loveland, K.L.; Klein, B.; Pueschl, D.; Indumathy, S.; Bergmann, M.; Loveland, B.E.; Hedger, M.P.; Schuppe, H.-C. Cytokines in Male Fertility and Reproductive Pathologies: Immunoregulation and Beyond. Front. Endocrinol. 2017, 8, 307. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, M.; Cai, F.; Song, W. Biological function of Presenilin and its role in AD pathogenesis. Transl. Neurodegener. 2013, 2, 15. [Google Scholar] [CrossRef] [Green Version]
- Fardilha, M.; Vieira, S.I.; Barros, A.; Sousa, M.; Da Cruz e Silva, O.A.B.; Da Cruz e Silva, E.F. Differential distribution of Alzheimer’s amyloid precursor protein family variants in human sperm. Ann. N. Y. Acad. Sci. 2007, 1096, 196–206. [Google Scholar] [CrossRef]
- Silva, J.V.; Yoon, S.; Domingues, S.; Guimarães, S.; Goltsev, A.V.; da Cruz e Silva, E.F.; Mendes, J.F.F.; da Cruz e Silva, O.A.B.; Fardilha, M. Amyloid precursor protein interaction network in human testis: Sentinel proteins for male reproduction. BMC Bioinform. 2015, 16, 12. [Google Scholar] [CrossRef] [Green Version]
- Komiya, Y.; Habas, R. Wnt signal transduction pathways. Organogenesis 2008, 4, 68–75. [Google Scholar] [CrossRef] [Green Version]
- Cheng, J.-M.; Tang, J.-X.; Li, J.; Wang, Y.-Q.; Wang, X.-X.; Zhang, Y.; Chen, S.-R.; Liu, Y.-X. Role of WNT signaling in epididymal sperm maturation. J. Assist. Reprod. Genet. 2018, 35, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Koch, S.; Acebron, S.P.; Herbst, J.; Hatiboglu, G.; Niehrs, C. Post-transcriptional Wnt signaling governs epididymal sperm maturation. Cell 2015, 163, 1225–1236. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DAPs | MW (kDa) | Optical Density | |
|---|---|---|---|
| SIPs | SMAPs | ||
| a | 71.8 | 0.36 ± 0.012 a | 0.21 ± 0.006 b |
| b | 65.0 | 0.32 ± 0.013 a | 0.19 ± 0.005 b |
| c | 20.5 | 0.29 ± 0.008 a | 0.17 ± 0.007 b |
| d | 12.8 | 0.28 ± 0.006 a | 0.17 ± 0.005 b |
| e | 11.9 | 0.30 ± 0.007 a | 0.18 ± 0.005 b |
| f | 57.5 | 0.22 ± 0.009 a | 0.28 ± 0.006 b |
| g | 50.0 | 0.18 ± 0.006 a | 0.32 ± 0.008 b |
| h | 38.6 | 0.17 ± 0.006 a | 0.29 ± 0.014 b |
| i | 18.0 | 0.18 ± 0.007 a | 0.29 ± 0.006 b |
| j | 10.9 | 0.20 ± 0.004 a | 0.31 ± 0.013 b |
| DAPs | Protein Name | Swiss-Prot Accession Number | Gene Symbol | Sequence Coverage (%) | Molecular Weight (kDa) | Peptide Counts (Unique) | Peptide Counts (All) | Protein Score |
|---|---|---|---|---|---|---|---|---|
| SIPs | ||||||||
| a | Lactotransferrin | F1PR54 | LTF | 34.6 | 77.3 | 21 | 72 | 440 |
| Carboxylesterase 5A | Q6AW47 | CES5A | 15.1 | 63.6 | 6 | 44 | 226 | |
| b | Albumin | P49822 | ALB | 9.5 | 68.6 | 4 | 25 | 337 |
| Olfactomedin 4 | F1PB68 | OLFM4 | 4.6 | 54.4 | 1 | 6 | 236 | |
| c | Prostaglandin-H2 D-isomerase | Q9XS65 | PTGDS | 28.3 | 21.1 | 5 | 25 | 212 |
| Glutathione peroxidase | F1PJ71 | GPX5 | 16.7 | 25.3 | 3 | 20 | 282 | |
| d | Lipocalin_cytosolic_FA-bd domain-containing protein | E2R6E0 | LCNL1 | 3.7 | 32.6 | 1 | 7 | 229 |
| Peptidyl-prolyl cis-trans isomerase | F1PLV2 | CSNK1G1 | 5.4 | 26.6 | 1 | 1 | 147 | |
| e | NPC intracellular cholesterol transporter 2 | Q28895 | NPC2 | 12.1 | 16.1 | 1 | 33 | 247 |
| Leucine rich repeat neuronal protein 3 | F1PYL2 | LRRN3 | 1.6 | 79.6 | 1 | 2 | 135 | |
| SMAPs | ||||||||
| f | Epididymal secretory protein E1 | F1PAR9 | NPC2 | 33.3 | 20.2 | 4 | 29 | 470 |
| Lactotransferrin | F1PR54 | LTF | 17.4 | 77.3 | 12 | 22 | 335 | |
| g | Cysteine-rich secretory protein 2 | A0A5F4CCD | CRISP2 | 13.5 | 35.4 | 2 | 5 | 302 |
| WAP domain-containing protein | E2RCT1 | N/A * WAPdcp ** | 14.7 | 13.0 | 1 | 3 | 303 | |
| h | Actin, cytoplasmic 1 | O18840 | ACTB | 5.3 | 41.7 | 4 | 41 | 378 |
| Ubiquitin-60S ribosomal protein L40 | P63050 | UBA52 | 7.0 | 14.7 | 1 | 4 | 262 | |
| i | Epididymal secretory protein E1 | F1PAR9 | NPC2 | 25.4 | 20.2 | 3 | 29 | 591 |
| Beta-N-acetylhexosaminidase | F1Q1M8 | HEXB | 6.7 | 38.1 | 1 | 4 | 276 | |
| j | Epididymal secretory protein E1 | F1PAR9 | NPC2 | 32.3 | 20.2 | 3 | 21 | 540 |
| Acrosin-binding protein | E2RNS8 | ACRBP | 1.9 | 61.3 | 1 | 3 | 231 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zmudzinska, A.; Bromke, M.A.; Strzezek, R.; Zielinska, M.; Olejnik, B.; Mogielnicka-Brzozowska, M. Proteomic Analysis of Intracellular and Membrane-Associated Fractions of Canine (Canis lupus familiaris) Epididymal Spermatozoa and Sperm Structure Separation. Animals 2022, 12, 772. https://doi.org/10.3390/ani12060772
Zmudzinska A, Bromke MA, Strzezek R, Zielinska M, Olejnik B, Mogielnicka-Brzozowska M. Proteomic Analysis of Intracellular and Membrane-Associated Fractions of Canine (Canis lupus familiaris) Epididymal Spermatozoa and Sperm Structure Separation. Animals. 2022; 12(6):772. https://doi.org/10.3390/ani12060772
Chicago/Turabian StyleZmudzinska, Anna, Mariusz A. Bromke, Rafal Strzezek, Magdalena Zielinska, Beata Olejnik, and Marzena Mogielnicka-Brzozowska. 2022. "Proteomic Analysis of Intracellular and Membrane-Associated Fractions of Canine (Canis lupus familiaris) Epididymal Spermatozoa and Sperm Structure Separation" Animals 12, no. 6: 772. https://doi.org/10.3390/ani12060772