
Positional Behavior and Substrate Use in Wild Tibetan Macaques

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Methods
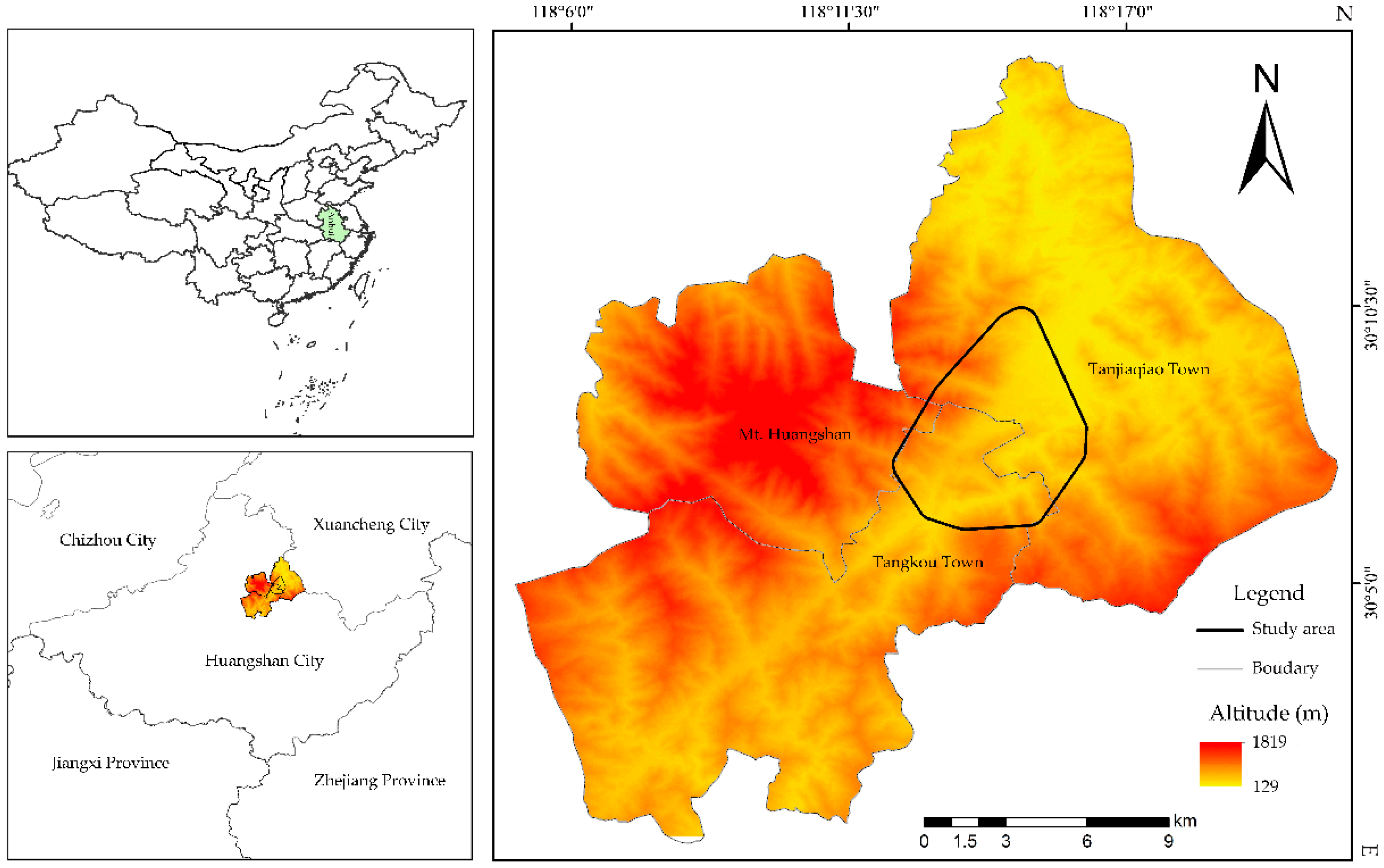
2.1. Study Site
2.2. Study Groups
2.3. Behavioral Data Collection
2.4. Data Analysis
2.5. Ethics Statement
3. Results
3.1. Description of Positional Behavior and Substrate Use
3.2. Age- and Sex-Based Differences in Positional Behavior
3.3. Age- and Sex-Based Differences in Substrate Use
4. Discussion
4.1. Positional Behavior of Tibetan Macaques
4.2. Effect of Body Mass on Intraspecific Positional Behavior and Substrate Use
4.3. Ontogeny of Positional Behavior in Tibetan Macaque
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Prost, J.H. A definitional system for the classification of primate locomotion. Am. Anthropol. 1965, 67, 1198–1214. [Google Scholar] [CrossRef]
- Napier, J.R.; Napier, P.H. A Handbook of Living Primates; Morphology, Ecology and Behaviour of Nonhuman Primates; Academic Press: London, UK, 1967. [Google Scholar]
- Subramaniam, V. Chemical Composition and Digestibility of Natural and Domestic Food of the Lar Gibbon (Hylobates lar) in Malaysia. Ph.D. Thesis, Universiti Pertanian Malaysia, Selangor, Malaysia, 1981. [Google Scholar]
- Gebo, D.L.; Chapman, C.A. Positional behavior in five sympatric Old World monkeys. Am. J. Phys. Anthropol. 1995, 97, 49–76. [Google Scholar] [CrossRef] [PubMed]
- Walker, S.E. The Evolution of Positional Behavior in the Saki-Uakaris (Pithecia, Chiropotes, and Cacajao); Plenum Press: New York, NY, USA, 1996. [Google Scholar]
- Lawler, R.R.; Stamps, C. The relationship between tail use and positional behavior in Alouatta palliata. Primates 2002, 43, 147–152. [Google Scholar] [CrossRef]
- Mittermeier, R.A. Locomotion and Posture in Ateles geoffroyi and Ateles paniscus. Folia Primatol. 1978, 30, 161–193. [Google Scholar] [CrossRef]
- Zhou, Q.H.; Luo, B.; Wei, F.; Huang, C. Habitat use and locomotion of the Francois’ langur (Trachypithecus francoisi) in limestone habitats of Nonggang, China. Integr. Zool. 2013, 8, 346–355. [Google Scholar] [CrossRef] [PubMed]
- Wright, K.A.; Biondi, L.; Visalberghi, E.; Ma, Z.; Izar, P.; Fragaszy, D. Positional behavior and substrate use in wild adult bearded capuchin monkeys (Sapajus libidinosus). Am. J. Primatol. 2019, 81, e23067. [Google Scholar] [CrossRef]
- Fleagle, J.G.; Mittermeier, R.A. Locomotor behavior, body size, and comparative ecology of seven Surinam monkeys. Am. J. Phys. Anthropol. 1980, 52, 301–314. [Google Scholar] [CrossRef]
- Rodman, P.S. Structural differentiation of microhabitats of sympatric macaca fascicularis and M. nemestrina in East Kalimantan, Indonesia. Int. J. Primatol. 1991, 12, 357–375. [Google Scholar] [CrossRef]
- Chatani, K. Positional behavior of free-ranging Japanese macaques (Macaca fuscata). Primates 2003, 44, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.H.; Huang, C.M.; Wei, H.; Tang, H.X.; Krzton, A.L.; Ma, G.Z.; Zhou, Q.H. Factors influencing positional behavior and habitat use of sympatric macaques in the limestone habitat of Nonggang, China. Int. J. Primatol. 2015, 36, 95–112. [Google Scholar] [CrossRef]
- Hershkovitz, P. Notes on tertiary platyrrhine monkeys and description of a new genus from the late Miocene of Colombia. Folia Primatol. 1970, 12, 1–37. [Google Scholar] [CrossRef] [PubMed]
- Cartmill, M.; Milton, K. The lorisiform wrist joint and the evolution of "brachiating" adaptations in the hominoidea. Am. J. Phys. Anthropol. 1977, 47, 249–272. [Google Scholar] [CrossRef] [PubMed]
- Garber, P.A. Locomotor behavior and feeding ecology of the panamanian tamarin (Saguinus oedipus geoffroyi, callitrichidae, primates). Int. J. Primatol. 1980, 1, 185–201. [Google Scholar] [CrossRef]
- Fleagle, J.G. Primate Adaptation and Evolution, 3rd ed.; Academic Press: New York, NY, USA, 2013. [Google Scholar]
- McGraw, W.S. Comparative locomotion and habitat use of six monkeys in the Tai Forest, Ivory Coast. Am. J. Phys. Anthropol. 1998, 105, 493–510. [Google Scholar] [CrossRef]
- Workman, C.; Covert, H.H. Learning the ropes: The ontogeny of locomotion in red-shanked douc (Pygathrix nemaeus), Delacour’s (Trachypithecus delacouri), and Hatinh Langurs (Trachypithecus hatinhensis) I. Positional behavior. Am. J. Phys. Anthropol. 2005, 128, 371–380. [Google Scholar] [CrossRef]
- Bezanson, M.; Morbeck, M.E. Future adults or old children? Integrating life history frameworks for understanding primate positional patterns. In Building Babies: Primate Development in Proximate and Ultimate Perspective; Clancy, K., Hinde, K., Rutherford, J., Eds.; Springer Book Series: New York, USA, 2013; pp. 435–458. [Google Scholar]
- Dunham, N.T. Ontogeny of positional behavior and support use among Colobus angolensis palliatus of the Diani Forest, Kenya. Primates 2015, 56, 183–192. [Google Scholar] [CrossRef]
- Young, J.W.; Shapiro, L.J. Developments in development: What have we learned from primate locomotor ontogeny? Am. J. Phys. Anthropol. 2018, 165, 37–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hurov, J.R. Rethinking primate locomotion: What can we learn from development? J. Motor. Behav. 1991, 23, 211. [Google Scholar] [CrossRef]
- Turnquist, J.E.; Wells, J.P. Ontogeny of locomotion in rhesus macaques (Macaca mulatta): I. Early postnatal ontogeny of the musculoskeletal system. J. Hum. Evol. 1994, 26, 487–499. [Google Scholar] [CrossRef]
- Ishida, H.; Kumakura, H.; Kondo, S. Primate Bipedalism and Quadrupedalism: Comparative Electromyography; University of Tokyo: Tokyo, Japan, 1985. [Google Scholar]
- Bezanson, M. The Ontogeny of Prehensile-Tail Use in Cebus capucinus and Alouatta palliata. Am. J. Primatol. 2012, 74, 770–782. [Google Scholar] [CrossRef]
- Druelle, F.; Aerts, P.; Berillon, G. Effect of body mass distribution on the ontogeny of positional behaviors in non-human primates: Longitudinal follow-up of infant captive olive baboons (Papio anubis). Am. J. Primatol. 2016, 78, 1201–1221. [Google Scholar] [CrossRef] [PubMed]
- Young, J.W. Ontogeny of muscle mechanical advantage in capuchin monkeys (Cebus albifrons and Cebus apella). J. Zool. 2005, 267, 351–362. [Google Scholar] [CrossRef]
- Sarringhaus, L.A.; MacLatchy, L.M.; Mitani, J.C. Locomotor and postural development of wild chimpanzees. J. Hum. Evol. 2014, 66, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Doran, D.M. The ontogeny of chimpanzee and pygmy chimpanzee locomotor behavior: A case study of paedomorphism and its behavioral correlates. J. Hum. Evol. 1992, 23, 139–157. [Google Scholar] [CrossRef]
- Zhu, W.W.; Garber, P.A.; Bezanson, M.; Qi, X.G.; Li, B.G. Age- and sex-based patterns of positional behavior and substrate utilization in the golden snub-nosed monkey (Rhinopithecus roxellana). Am. J. Primatol. 2015, 77, 98–108. [Google Scholar] [CrossRef] [PubMed]
- Li, J.H.; Sun, L.X.; Kappeler, P.M. The Behavioral Ecology of the Tibetan Macaque; Springer International Publishing: Cham, Switzerland, 2020. [Google Scholar]
- Jiang, X.L.; Wang, Q.S. Taxonomy and distribution of Tibetan macaque (Macaca thibetana). Zool. Res. 1996, 17, 361–369. (In Chinese) [Google Scholar]
- Xiong, C.P. Ecological studies of the stump-tailed macaque. Acta Theriol. Sin. 1984, 4, 1–9. (In Chinese) [Google Scholar]
- Zhao, Q.K.; Deng, Z.Y. Macaca thibetana at Mt. Emei, china: I. a cross-sectional study of growth and development. Am. J. Primatol. 1988, 16, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Li, W.B.; Yang, P.P.; Li, B.W.; Liu, C.; Sun, L.X.; Li, J.H. Habitat characteristics or protected area size: What is more important for the composition and diversity of mammals in nonprotected areas? Ecol. Evol. 2021, 11, 7250–7263. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.H.; Pan, R.L. A comparative study on the grow th of long bones in M. Tibetana and M. mulata. Zool. Res. 1989, 10, 11–18. (In Chinese) [Google Scholar]
- Altmann, J. Observational study of behavior: Sampling methods. Behaviour 1974, 49, 227–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mekonnen, A.; Fashing, P.J.; Sargis, E.J.; Venkataraman, V.V.; Stenseth, N.C. Flexibility in positional behavior, strata use, and substrate utilization among Bale monkeys (Chlorocebus djamdjamensis) in response to habitat fragmentation and degradation. Am. J. Primatol. 2018, 80, e22760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunt, K.D.; Cant, J.G.H.; Gebo, D.L.; Rose, M.D.; Walker, S.E.; Youlatos, D. Standardized descriptions of primate locomotor and postural modes. Primates 1996, 37, 363–387. [Google Scholar] [CrossRef]
- Fan, P.; Scott, M.B.; Fei, H.; Ma, C. Locomotion behavior of cao vit gibbon (Nomascus nasutus) living in karst forest in Bangliang Nature Reserve, Guangxi, China. Integr. Zool. 2013, 8, 356–364. [Google Scholar] [CrossRef]
- Fei, H.L.; Ma, C.Y.; Bartlett, T.Q.; Dai, R.; Xiao, W.; Fan, P.F. Feeding postures of cao vit gibbons (Nomascus nasutus) living in a low-canopy karst forest. Int. J. Primatol. 2015, 36, 1036–1054. [Google Scholar] [CrossRef]
- Isbell, L.A.; Pruetz, J.D.; Lewis, M.; Young, T.P. Locomotor activity differences between sympatric patas monkeys (Erythrocebus patas) and vervet monkeys (Cercopithecus aethiops): Implications for the evolution of long hindlimb length in Homo. Am. J. Phys. Anthropol. 1998, 105, 199–207. [Google Scholar] [CrossRef]
- Motsch, P.; Le Flohic, G.; Dilger, C.; Delahaye, A.; Chateau-Smith, C.; Couette, S. Degree of Terrestrial Activity of the Elusive Sun-Tailed Monkey (Cercopithecus solatus) in Gabon: Comparative Study of Behavior and Postcranial Morphometric Data. Am. J. Primatol. 2015, 77, 1060–1074. [Google Scholar] [CrossRef]
- Hanna, J.B.; Schmitt, D. Interpreting the Role of Climbing in Primate Locomotor Evolution: Are the Biomechanics of Climbing Influenced by Habitual Substrate Use and Anatomy? Int. J. Primatol. 2011, 32, 430–444. [Google Scholar] [CrossRef]
- Cant, J.G. Positional behavior of long-tailed macaques (Macaca fascicularis) in northern Sumatra. Am. J. Phys. Anthropol. 1988, 76, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Pan, R.L.; Wang, H. Long bone and skeletal allometry in Rhinopithecus. Zool. Res. 1989, 10, 23–30. (In Chinese) [Google Scholar]
- Garber, P.A. Primate locomotor bahvior and ecology. In Primates in Perspective; Campbell, C., Fuentes, A., MacKinnon, K.C., Panger, M., Bearder, S.K., Eds.; Oxford University Press: New York, NY, USA, 2007. [Google Scholar]
- Rawlins, R.G. Locomotor ontogeny in Macaca mulatta: I. Behavioral strategies and tactics. Am. J. Phys. Anthropol. 1976, 44, 201. [Google Scholar]
- Prates, H.M.; Bicca-Marques, J.C. Age-sex analysis of activity budget, diet, and positional behavior in Alouatta caraya in an orchard forest. Int. J. Primatol. 2008, 29, 703–715. [Google Scholar] [CrossRef]
- Wang, Q.S. Mamals in Anhui Province; Anhui Science and Technology Press: Hefei, China, 1990. (In Chinese) [Google Scholar]
- Youlatos, D. Seasonal variation in the positional behavior of red howling monkeys (Alouatta seniculus). Primates 1998, 39, 449–457. [Google Scholar] [CrossRef]
- Dunbar, D.C. Locomotor behavior of rhesus macaques (Macaca mulatta) on Cayo Santiago. P. R. Health Sci. J. 1989, 8, 79–85. [Google Scholar] [PubMed]
- Gregoire, B.-A.; Anthony, H.; Emmanuelle, P. Ontogeny of locomotion in mouse lemurs: Implications for primate evolution. J. Hum. Evol. 2020, 142, 102732. [Google Scholar] [CrossRef]
- Lawler, R.R. Sifaka positional behavior: Ontogenetic and quantitative genetic approaches. Am. J. Phys. Anthropol. 2006, 131, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Wang, X.; Garber, P.A.; Sun, B.-H.; Sun, L.; Xia, D.-P.; Li, J.-H. Sex-specific variation of social play in wild immature Tibetan macaques, Macaca thibetana. Animals 2021, 11, 805. [Google Scholar] [CrossRef] [PubMed]
- Wunderlich, R.E.; Lawler, R.R.; Williams, A.E. Field and Experimental Approaches to the Study of Locomotor Ontogeny in Propithecus verreauxi. In Primate Locomotion; Developments in Primatology: Progress and Prospects; Springer: New York, NY, USA, 2011; pp. 135–154. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Terms | Definitions * |
|---|---|
| Activity | |
| Feeding | Searching for, chewing, and swallowing food |
| Traveling | Changing the position of the body in space by walking, running, leaping, climbing, etc., and not obtaining food within 5 s |
| Resting | Maintaining a stationary position without movement |
| Social grooming | One individual uses their fingers or palms to separate and smooth the hairs of another individual and occasionally picks up certain small particles from the separated hairs or exposed skin and puts them in their mouth to chew |
| Other | Play and unusual behaviors, including sexual behavior or aggression |
| Locomotor mode | |
| Quadrupedal Walking | Movement in a particular gait along a substrate with an inclination of less than 50° |
| Climbing | Quadrupedal movement on a large inclination (substrate angle > 50°) or unstable substrate |
| Leaping | Crossing the substrate gap involving free flight movements in which the hind limbs provide propulsion |
| Quadrupedal running | Similar to quadrupedal walking, but faster and with a brief period of free flight in the air (i.e., all limbs off the ground) |
| Bridging | Movement to cross the substrate gap with the hind limbs grasping one side of the substrate and the forelimbs grasping the other side of the substrate as the body slowly moves towards the front |
| Postural mode | |
| Sitting | Relies primarily on the ischia to support most of the body’s weight |
| Quadrupedal standing | Relies primarily on the front and hind limbs to support most of the body weight, with the trunk horizontal |
| Bipedal standing | Relies primarily on the hind limbs to support most of the bodyweight, upright or bent, with the forelimbs sometimes touching the substrates. |
| Lying | The torso rests relatively horizontally above the substrate, primarily supporting the weight |
| Suspension | The forelimbs or hindlimbs grip the substrate firmly to support the body, and the rest of the body does not touch the substrate, and the torso is relatively extended |
| Clinging | Body against a vertical substrate with hands gripping or holding the substrate firmly to maintain stability |
| Substrate height | |
| Ground | Substrate with only 0 m vertical height |
| Lower strata | 0–5 m height above ground |
| Middle strata | 5–10 m height above ground |
| Upper strata | Height above 10 m above ground |
| Substrate size | |
| Twigs | Terminal branches usually smaller than 2 cm in diameter |
| Branches | Monkeys can usually grasp branches between 2 cm and 10 cm in diameter |
| Boughs | Substrate diameter greater than 10 cm |
| Mean | SD | |
|---|---|---|
| Locomotor mode | ||
| Climbing | 24.9 | 6.3 |
| Bridging | 2.7 | 2.8 |
| Quadrupedal running | 7.8 | 4.0 |
| Quadrupedal walking | 57.9 | 9.4 |
| Leaping | 7.1 | 5.6 |
| Postural mode | ||
| Clinging | 2.8 | 1.4 |
| Bipedal standing | 1.0 | 0.7 |
| Quadrupedal standing | 9.0 | 4.8 |
| Suspension | 0.8 | 0.7 |
| Siting | 85.8 | 4.9 |
| Lying | 1.5 | 0.9 |
| Substrate height | ||
| Ground | 61.3 | 10.9 |
| Lower forest strata | 18.9 | 4.4 |
| Middle forest strata | 12.8 | 5.6 |
| Upper forest strata | 7.0 | 5.9 |
| Substrate size | ||
| Twigs | 8.3 | 3.9 |
| Branches | 55.5 | 5.1 |
| Boughs | 36.2 | 5.1 |
| Adult Males | Adult Females | Subadults | Juveniles | Infants | |
|---|---|---|---|---|---|
| Substrate height | |||||
| Ground | 80.83 | 76.03 | 61.57 | 41.61 a,b | 43.04 a,b |
| Lower | 10.13 | 10.46 | 14.70 | 29.37 a,b | 30.75 a,b |
| Middle | 8.20 | 9.13 | 21.00 | 24.74 a,b | 23.19 |
| Upper | 0.83 | 4.38 | 2.73 | 4.82 | 3.00 |
| Substrate size | |||||
| Bough | 24.24 | 19.29 | 12.65 | 11.72 a | 10.32 a,b |
| Branch | 58.47 | 63.39 | 61.45 | 58.17 | 48.78 |
| Twig | 17.29 | 17.32 | 25.90 | 30.11 a,b | 40.90 a,b |
| Adult Males | Adult Females | Subadults | Juveniles | Infants | |
|---|---|---|---|---|---|
| Substrate height | |||||
| Ground | 59.53 | 45.57 | 43.32 | 31.63 a | 30.12 a |
| Lower | 19.55 | 28.25 | 25.25 | 30.42 | 37.10 |
| Middle | 13.36 | 18.62 | 23.39 | 28.78 | 29.00 |
| Upper | 7.56 | 7.56 | 8.04 | 9.17 | 3.55 |
| Substrate size | |||||
| Bough | 2.94 | 0.51 | 3.26 | 1.26 | 2.65 |
| Branch | 59.72 | 61.73 | 67.73 | 40.16 a,b,c | 23.96 a,b,c |
| Twig | 37.34 | 37.77 | 29.01 | 58.57 a,b,c | 73.39 a,b,c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, P.-H.; Li, W.-B.; Li, B.-W.; Li, Y.-D.; Wang, X.; Li, J.-H. Positional Behavior and Substrate Use in Wild Tibetan Macaques. Animals 2022, 12, 767. https://doi.org/10.3390/ani12060767
Li P-H, Li W-B, Li B-W, Li Y-D, Wang X, Li J-H. Positional Behavior and Substrate Use in Wild Tibetan Macaques. Animals. 2022; 12(6):767. https://doi.org/10.3390/ani12060767
Chicago/Turabian StyleLi, Peng-Hui, Wen-Bo Li, Bo-Wen Li, Ya-Dong Li, Xi Wang, and Jin-Hua Li. 2022. "Positional Behavior and Substrate Use in Wild Tibetan Macaques" Animals 12, no. 6: 767. https://doi.org/10.3390/ani12060767