
The Role of Aquaporin 7 in the Movement of Water and Cryoprotectants in Bovine In Vitro Matured Oocytes

 , , , ,
, , , ,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Oocyte Collection and In Vitro Maturation
2.3. Analysis of AQP3, AQP7 and AQP9 Expression
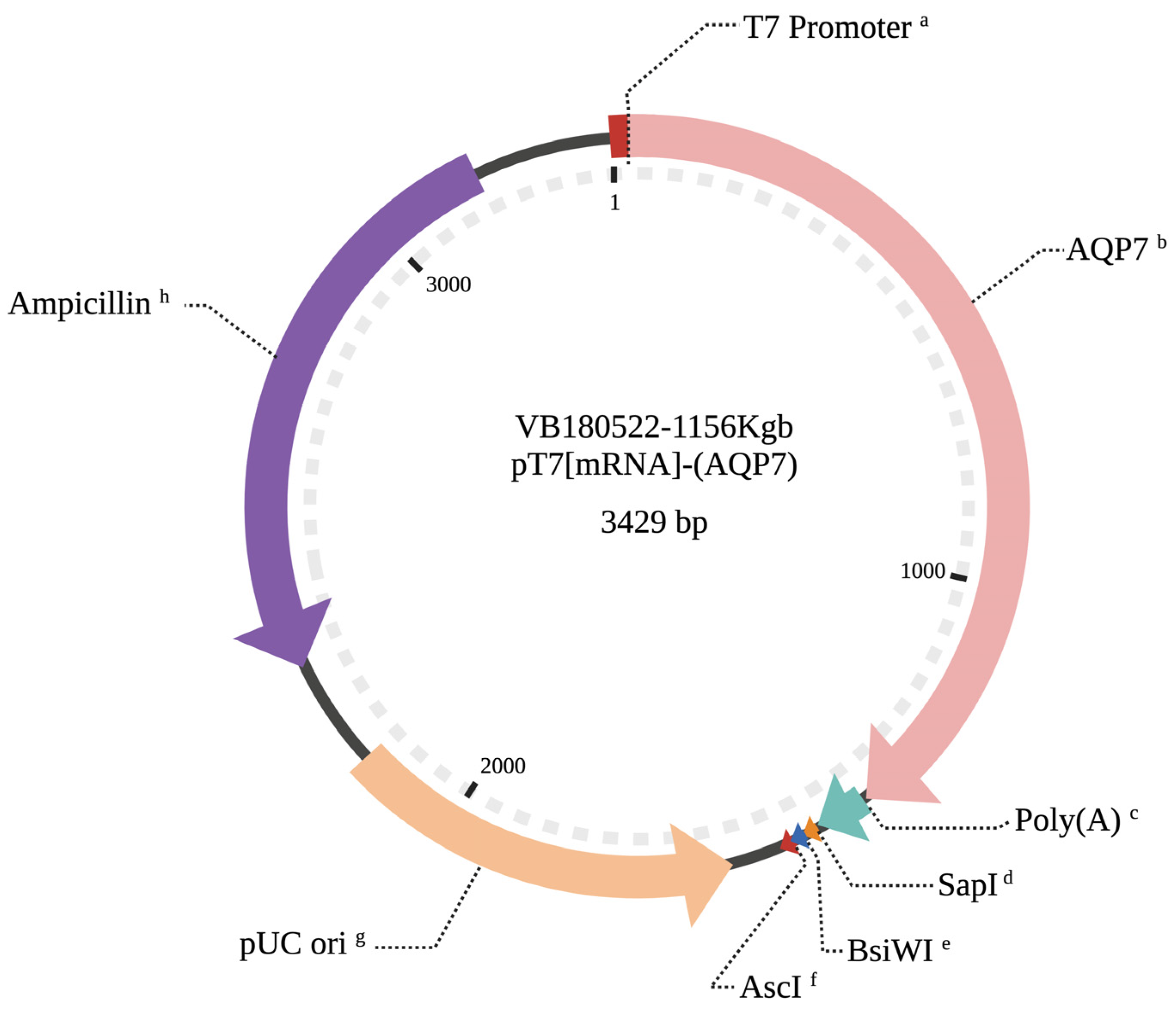
2.4. Preparation of AQP7 cRNA
2.5. Microinjection of AQP7 cRNA into Bovine Oocytes
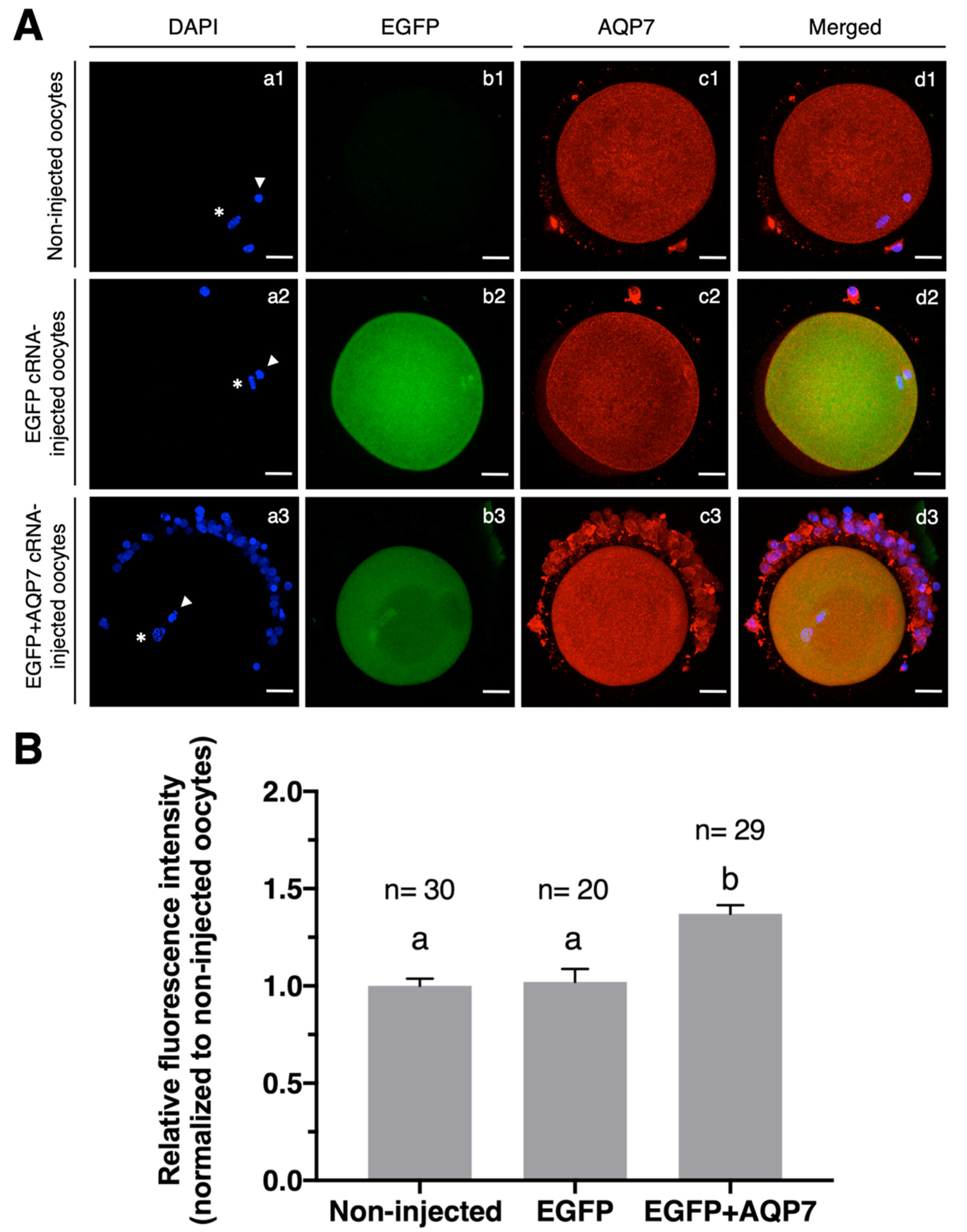
2.6. Expression of AQP7 in cRNA-Injected Oocytes
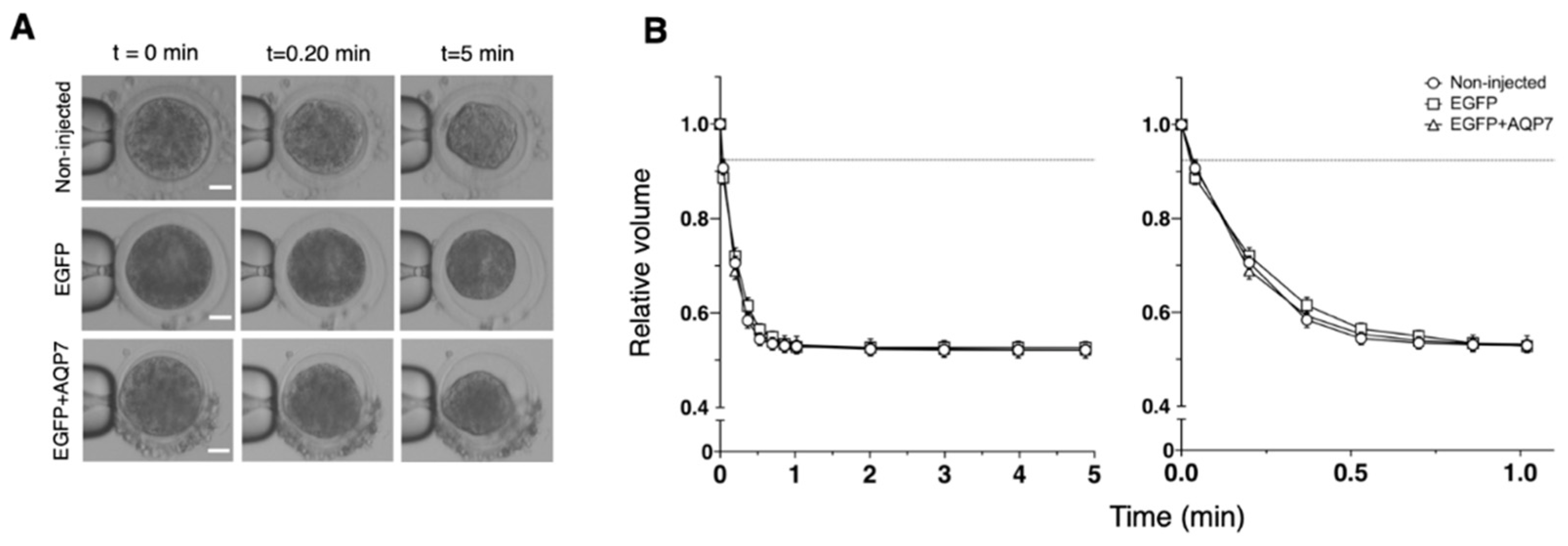
2.7. Measurement of Permeability to Water and Cryoprotectants
2.8. Experimental Design
2.9. Statistical Analysis
3. Results
3.1. Analysis of AQP3, AQP7 and AQP9 Expression in Oocytes after Treatment with Cryoprotection Solutions
3.2. Measurement of Permeability to Water and Cryoprotectants of In Vitro Matured Oocytes in Which AQP7 Was Artificially Overexpressed
3.2.1. Expression of AQP7 in cRNA-Injected Oocytes
3.2.2. Permeability to Water and Cryoprotectants of AQP7 cRNA-Injected Oocytes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Diez, C.; Munoz, M.; Caamano, J.N.; Gomez, E. Cryopreservation of the bovine oocyte: Current status and perspectives. Reprod. Domest. Anim. Zuchthyg. 2012, 47 (Suppl. 3), 76–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mogas, T. Update on the vitrification of bovine oocytes and invitro-produced embryos. Reprod. Fertil. Dev. 2018, 31, 105–117. [Google Scholar] [CrossRef] [PubMed]
- Rall, W.F.; Fahy, G.M. Ice-free cryopreservation of mouse embryos at -196 degrees C by vitrification. Nature 1985, 313, 573–575. [Google Scholar] [CrossRef] [PubMed]
- Agca, Y.; Liu, J.; Rutledge, J.J.; Critser, E.S.; Critser, J.K. Effect of osmotic stress on the developmental competence of germinal vesicle and metaphase II stage bovine cumulus oocyte complexes and its relevance to cryopreservation. Mol. Reprod. Dev. 2000, 55, 212–219. [Google Scholar] [CrossRef]
- Mullen, S.F.; Li, M.; Li, Y.; Chen, Z.J.; Critser, J.K. Human oocyte vitrification: The permeability of metaphase II oocytes to water and ethylene glycol and the appliance toward vitrification. Fertil. Steril. 2008, 89, 1812–1825. [Google Scholar] [CrossRef] [Green Version]
- Mullen, S.F.; Agca, Y.; Broermann, D.C.; Jenkins, C.L.; Johnson, C.A.; Critser, J.K. The effect of osmotic stress on the metaphase II spindle of human oocytes, and the relevance to cryopreservation. Hum. Reprod. 2004, 19, 1148–1154. [Google Scholar] [CrossRef] [Green Version]
- Fahy, G.M.; Wowk, B.; Wu, J.; Paynter, S. Improved vitrification solutions based on the predictability of vitrification solution toxicity. Cryobiology 2004, 48, 22–35. [Google Scholar] [CrossRef]
- Verkman, A.S.; Anderson, M.O.; Papadopoulos, M.C. Aquaporins: Important but elusive drug targets. Nat. Rev. Drug Discov. 2014, 13, 259–277. [Google Scholar] [CrossRef] [Green Version]
- Agca, Y.; Liu, J.; McGrath, J.J.; Peter, A.T.; Critser, E.S.; Critser, J.K. Membrane permeability characteristics of metaphase II mouse oocytes at various temperatures in the presence of Me2SO. Cryobiology 1998, 36, 287–300. [Google Scholar] [CrossRef]
- Agca, Y.; Liu, J.; Critser, E.S.; McGrath, J.J.; Critser, J.K. Temperature-dependent osmotic behavior of germinal vesicle and metaphase II stage bovine oocytes in the presence of Me2SO in relationship to cryobiology. Mol. Reprod. Dev. 1999, 53, 59–67. [Google Scholar] [CrossRef]
- Benson, J.D.; Kearsley, A.J.; Higgins, A.Z. Mathematical optimization of procedures for cryoprotectant equilibration using a toxicity cost function. Cryobiology 2012, 64, 144–151. [Google Scholar] [CrossRef] [PubMed]
- King, L.S.; Kozono, D.; Agre, P. From structure to disease: The evolving tale of aquaporin biology. Nat. Rev. Mol. Cell. Biol. 2004, 5, 687–698. [Google Scholar] [CrossRef] [PubMed]
- Verkman, A.S.; Mitra, A.K. Structure and function of aquaporin water channels. Am. J. Physiol. Renal. Physiol. 2000, 278, F13-28. [Google Scholar] [CrossRef]
- Huang, H.F.; He, R.H.; Sun, C.C.; Zhang, Y.; Meng, Q.X.; Ma, Y.Y. Function of aquaporins in female and male reproductive systems. Hum. Reprod. Update 2006, 12, 785–795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edashige, K.; Ohta, S.; Tanaka, M.; Kuwano, T.; Valdez, D.M., Jr.; Hara, T.; Jin, B.; Takahashi, S.; Seki, S.; Koshimoto, C.; et al. The role of aquaporin 3 in the movement of water and cryoprotectants in mouse morulae. Biol. Reprod. 2007, 77, 365–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, B.; Kawai, Y.; Hara, T.; Takeda, S.; Seki, S.; Nakata, Y.; Matsukawa, K.; Koshimoto, C.; Kasai, M.; Edashige, K. Pathway for the movement of water and cryoprotectants in bovine oocytes and embryos. Biol. Reprod. 2011, 85, 834–847. [Google Scholar] [CrossRef]
- Morató, R.; Chauvigné, F.; Novo, S.; Bonet, S.; Cerdà, J. Enhanced water and cryoprotectant permeability of porcine oocytes after artificial expression of human and zebrafish aquaporin-3 channels. Mol. Reprod. Dev. 2014, 81, 450–461. [Google Scholar] [CrossRef] [PubMed]
- Yamaji, Y.; Seki, S.; Matsukawa, K.; Koshimoto, C.; Kasai, M.; Edashige, K. Developmental ability of vitrified mouse oocytes expressing water channels. J. Reprod. Dev. 2011, 57, 403–408. [Google Scholar] [CrossRef] [Green Version]
- Edashige, K.; Yamaji, Y.; Kleinhans, F.W.; Kasai, M. Artificial expression of aquaporin-3 improves the survival of mouse oocytes after cryopreservation. Biol. Reprod. 2003, 68, 87–94. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Martinez, T.; Vendrell-Flotats, M.; Martinez-Rodero, I.; Ordonez-Leon, E.A.; Alvarez-Rodriguez, M.; Lopez-Bejar, M.; Yeste, M.; Mogas, T. Glutathione Ethyl Ester Protects In Vitro-Maturing Bovine Oocytes against Oxidative Stress Induced by Subsequent Vitrification/Warming. Int. J. Mol. Sci. 2020, 21, 7547. [Google Scholar] [CrossRef]
- Jin, B.; Higashiyama, R.; Nakata, Y.; Yonezawa, J.; Xu, S.; Miyake, M.; Takahashi, S.; Kikuchi, K.; Yazawa, K.; Mizobuchi, S.; et al. Rapid movement of water and cryoprotectants in pig expanded blastocysts via channel processes: Its relevance to their higher tolerance to cryopreservation. Biol. Reprod. 2013, 89, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, Y.J.; Zhang, X.Y.; Ding, G.L.; Li, R.; Wang, L.; Jin, L.; Lin, X.H.; Gao, L.; Sheng, J.Z.; Huang, H.F. Aquaporin7 plays a crucial role in tolerance to hyperosmotic stress and in the survival of oocytes during cryopreservation. Sci. Rep. 2015, 5, 17741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamagata, K.; Yamazaki, T.; Yamashita, M.; Hara, Y.; Ogonuki, N.; Ogura, A. Noninvasive visualization of molecular events in the mammalian zygote. Genesis 2005, 43, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Redondo, I.; Ramos-Ibeas, P.; Pericuesta, E.; Fernández-González, R.; Laguna-Barraza, R.; Gutiérrez-Adán, A. Minor Splicing Factors Zrsr1 and Zrsr2 Are Essential for Early Embryo Development and 2-Cell-Like Conversion. Int. J. Mol. Sci. 2020, 21, 4115. [Google Scholar] [CrossRef] [PubMed]
- Luciano, A.M.; Lodde, V.; Beretta, M.S.; Colleoni, S.; Lauria, A.; Modina, S. Developmental capability of denuded bovine oocyte in a co-culture system with intact cumulus-oocyte complexes: Role of cumulus cells, cyclic adenosine 3’,5’-monophosphate, and glutathione. Mol. Reprod. Dev. 2005, 71, 389–397. [Google Scholar] [CrossRef] [PubMed]
- Kleinhans, F.W. Membrane permeability modeling: Kedem-Katchalsky vs a two-parameter formalism. Cryobiology 1998, 37, 271–289. [Google Scholar] [CrossRef]
- Jacobs, M.H.; Stewart, D.R. A simple method for the quantitative measurement of cell permeability. J. Cell. Comp. Physiol. 1932, 1, 71–82. [Google Scholar] [CrossRef]
- Jacobs, M.H. The simultaneous measurement of cell permeability to water and to dissolved substances. J. Cell. Comp. Physiol. 1933, 2, 427–444. [Google Scholar] [CrossRef]
- Dormand, J.R.; Prince, P.J. A family of embedded Runge-Kutta formulae. J. Comput. Appl. Math. 1980, 6, 19–26. [Google Scholar] [CrossRef] [Green Version]
- Shampine, L.F.; Reichelt, M.W. The MATLAB ODE Suite. SIAM J. Sci. Comput. 1997, 18, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Lagarias, J.C.; Reeds, J.A.; Wright, M.H.; Wright, P.E. Convergence Properties of the Nelder--Mead Simplex Method in Low Dimensions. SIAM J. Optim. 1998, 9, 112–147. [Google Scholar] [CrossRef] [Green Version]
- Ruffing, N.A.; Steponkus, P.L.; Pitt, R.E.; Parks, J.E. Osmometric behavior, hydraulic conductivity, and incidence of intracellular ice formation in bovine oocytes at different developmental stages. Cryobiology 1993, 30, 562–580. [Google Scholar] [CrossRef] [PubMed]
- Vian, A.M.; Higgins, A.Z. Membrane permeability of the human granulocyte to water, dimethyl sulfoxide, glycerol, propylene glycol and ethylene glycol. Cryobiology 2014, 68, 35–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2006. [Google Scholar] [CrossRef]
- Ohashi, S.; Naito, K.; Liu, J.; Sheng, Y.; Yamanouchi, K.; Tojo, H. Expression of Exogenous Proteins in Porcine Maturing Oocytes after mRNA Injection: Kinetic Analysis and Oocyte Selection Using EGFP mRNA. J. Reprod. Dev. 2001, 47, 351–357. [Google Scholar] [CrossRef] [Green Version]
- Edashige, K. Permeability of the plasma membrane to water and cryoprotectants in mammalian oocytes and embryos: Its relevance to vitrification. Reprod. Med. Biol. 2017, 16, 36–39. [Google Scholar] [CrossRef] [PubMed]
- Sales, A.D.; Lobo, C.H.; Carvalho, A.A.; Moura, A.A.; Rodrigues, A.P. Structure, function, and localization of aquaporins: Their possible implications on gamete cryopreservation. Genet. Mol. Res. GMR 2013, 12, 6718–6732. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.J.; Xiong, Y.; Ding, G.L.; Zhang, D.; Meng, Y.; Huang, H.F.; Sheng, J.Z. Cryoprotectants up-regulate expression of mouse oocyte AQP7, which facilitates water diffusion during cryopreservation. Fertil. Steril. 2013, 99, 1428–1435. [Google Scholar] [CrossRef]
- Balboula, A.Z.; Yamanaka, K.; Sakatani, M.; Hegab, A.O.; Zaabel, S.M.; Takahashi, M. Intracellular cathepsin B activity is inversely correlated with the quality and developmental competence of bovine preimplantation embryos. Mol. Reprod. Dev. 2010, 77, 1031–1039. [Google Scholar] [CrossRef]
- Kang, M.K.; Han, S.J. Post-transcriptional and post-translational regulation during mouse oocyte maturation. BMB Rep. 2011, 44, 147–157. [Google Scholar] [CrossRef] [Green Version]
- Sugiura, R.; Satoh, R.; Ishiwata, S.; Umeda, N.; Kita, A. Role of RNA-Binding Proteins in MAPK Signal Transduction Pathway. J. Signal Transduct. 2011, 2011, 109746. [Google Scholar] [CrossRef] [Green Version]
- Arima, H.; Yamamoto, N.; Sobue, K.; Umenishi, F.; Tada, T.; Katsuya, H.; Asai, K. Hyperosmolar mannitol simulates expression of aquaporins 4 and 9 through a p38 mitogen-activated protein kinase-dependent pathway in rat astrocytes. J. Biol. Chem. 2003, 278, 44525–44534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fong, B.; Watson, P.H.; Watson, A.J. Mouse preimplantation embryo responses to culture medium osmolarity include increased expression of CCM2 and p38 MAPK activation. BMC Dev. Biol. 2007, 7, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bell, C.E.; Lariviere, N.M.; Watson, P.H.; Watson, A.J. Mitogen-activated protein kinase (MAPK) pathways mediate embryonic responses to culture medium osmolarity by regulating Aquaporin 3 and 9 expression and localization, as well as embryonic apoptosis. Hum. Reprod. 2009, 24, 1373–1386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valdez, D.M., Jr.; Tsuchiya, R.; Seki, S.; Saida, N.; Niimi, S.; Koshimoto, C.; Matsukawa, K.; Kasai, M.; Edashige, K. A trial to cryopreserve immature medaka (Oryzias latipes) oocytes after enhancing their permeability by exogenous expression of aquaporin 3. J. Reprod. Dev. 2013, 59, 205–213. [Google Scholar] [CrossRef] [Green Version]
- Chauvigne, F.; Lubzens, E.; Cerda, J. Design and characterization of genetically engineered zebrafish aquaporin-3 mutants highly permeable to the cryoprotectant ethylene glycol. BMC Biotechnol. 2011, 11, 34. [Google Scholar] [CrossRef] [Green Version]
- Agca, Y.; Liu, J.; Peter, A.T.; Critser, E.S.; Critser, J.K. Effect of developmental stage on bovine oocyte plasma membrane water and cryoprotectant permeability characteristics. Mol. Reprod. Dev. 1998, 49, 408–415. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Description | Values | Symbol |
|---|---|---|
| Universal gas constant | 8.314 m3 Pa K−1 mol−1 | R |
| Absolute temperature | 298 K | T |
| Partial molar volume of water | 18.02 × 1012 μm3 mol−1 | υw |
| Partial molar volume of CPA: | υs | |
| 55.8 × 10−6 m3 mol−1 71.3 × 10−6 m3 mol−1 | |
| Osmotically inactive volume (MII oocytes): | 0.25 [16,32] | Vb |
| Treatment | n | % Survival | % EGFP * | %AQP7 § | % MII # |
|---|---|---|---|---|---|
| Non-injected | 105 | 100 a | - | - | 74.73 ± 1.93 a |
| EGFP | 84 | 71.48 ± 8.15 b | 88.31 ± 9.36 | 100 | 56.55 ± 3.92 b |
| EGFP+AQP7 | 135 | 62.11 ± 2.25 b | 82.88 ± 4.25 | 100 | 56.54 ± 1.24 b |
| Treatment | Sucrose |
|---|---|
| Non-injected | 2.19 ± 0.15 a |
| EGFP | 2.05 ± 0.18 a |
| EGFP+AQP7 | 2.17 ± 0.25 a |
| Treatment | Me2SO | EG |
|---|---|---|
| Non-injected | 2.06 ± 0.13 a | 1.38 ± 0.12 a |
| EGFP | 1.98 ± 0.24 a | 1.44 ± 0.10 a |
| EGFP+AQP7 | 3.96 ± 0.57 b | 1.39 ± 0.11 a |
| Treatment | Me2SO | EG |
|---|---|---|
| Non-injected | 0.58 ± 0.05 | 0.78 ± 0.09 |
| EGFP | 0.58 ± 0.05 | 1.04 ± 0.17 |
| EGFP+AQP7 | 0.57 ± 0.04 | 0.76 ± 0.10 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Martínez, T.; Martínez-Rodero, I.; Roncero-Carol, J.; Vendrell-Flotats, M.; Gardela, J.; Gutiérrez-Adán, A.; Ramos-Ibeas, P.; Higgins, A.Z.; Mogas, T. The Role of Aquaporin 7 in the Movement of Water and Cryoprotectants in Bovine In Vitro Matured Oocytes. Animals 2022, 12, 530. https://doi.org/10.3390/ani12040530
García-Martínez T, Martínez-Rodero I, Roncero-Carol J, Vendrell-Flotats M, Gardela J, Gutiérrez-Adán A, Ramos-Ibeas P, Higgins AZ, Mogas T. The Role of Aquaporin 7 in the Movement of Water and Cryoprotectants in Bovine In Vitro Matured Oocytes. Animals. 2022; 12(4):530. https://doi.org/10.3390/ani12040530
Chicago/Turabian StyleGarcía-Martínez, Tania, Iris Martínez-Rodero, Joan Roncero-Carol, Meritxell Vendrell-Flotats, Jaume Gardela, Alfonso Gutiérrez-Adán, Priscila Ramos-Ibeas, Adam Z. Higgins, and Teresa Mogas. 2022. "The Role of Aquaporin 7 in the Movement of Water and Cryoprotectants in Bovine In Vitro Matured Oocytes" Animals 12, no. 4: 530. https://doi.org/10.3390/ani12040530