
Nocardiosis in Free-Ranging Cetaceans from the Central-Eastern Atlantic Ocean and Contiguous Mediterranean Sea

 , , , , and
, , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Beaman, B.L.; Sugar, A.M. Nocardia in naturally acquired and experimental infections in animals. Epidemiol. Infect. 1983, 91, 393–419. [Google Scholar]
- Corti, M.E.; Fioti, M.E.V. Nocardiosis: A review. Int. J. Infect. Dis. 2003, 7, 243–250. [Google Scholar] [CrossRef] [Green Version]
- Parte, A.C.; Carbasse, J.S.; Meier-Kolthoff, J.P.; Reimer, L.C.; Goker, M. List of Prokaryotic names with Standing in Nomencla-ture (LPSN) moves to the DSMZ. Int. J. Syst. Evol. Microbiol. 2020, 70, 5607–5612. [Google Scholar] [CrossRef] [PubMed]
- Peters-Kennedy, J.M. Integumentary System. In Jubb, Kennedy and Palmer’s Pathology of Domestic Animals, 6th ed.; Maxie, M.G., Ed.; Elsevier: Maryland, MO, USA, 2016; pp. 511–580. [Google Scholar]
- Pier, A.C.; Fichtner, R.E. Distribution of serotypes of Nocardia asteroides from animal, human, and environmental sources. J. Clin. Microbiol. 1981, 13, 548–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conville, P.S.; Brown-Elliott, B.A.; Smith, T.; Zelazny, A.M. The Complexities of Nocardia Taxonomy and Identification. J. Clin. Microbiol. 2018, 56, e01419-17. [Google Scholar] [CrossRef] [Green Version]
- Mehta, H.H.; Shamoo, Y. Pathogenic Nocardia: A diverse genus of emerging pathogens or just poorly recognized? PLoS Pathog. 2020, 16, e1008280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lenskaya, V.; DeChavez, V.; Kaufman, B.; Caplivski, D. Primary cutaneous nocardiosis caused by Nocardia nova with possible apremilast contribution. IDCases 2021, 24, e01078. [Google Scholar] [CrossRef] [PubMed]
- Lerner, P.I. Nocardiosis, In Clinical Infectious Diseases; Oxford University Press: Oxford, UK, 1996; Volume 22. [Google Scholar]
- Shapiro, P.E.; Grossman, M.E. Disseminated Nocardia asteroides with pustules. J. Am. Acad. Dermatol. 1989, 20, 889–892. [Google Scholar] [CrossRef]
- Maekawa, S.; Yoshida, T.; Wang, P.-C.; Chen, S.-C. Current knowledge of nocardiosis in teleost fish. J. Fish Dis. 2018, 41, 413–419. [Google Scholar] [CrossRef] [PubMed]
- St Leger, J.A.; Begeman, L.; Fleetwood, M.; Frasca, S., Jr.; Garner, M.M.; Lair, S.; Trembley, S.; Linn M., J.; Terio, K.A. Comparative pathology of nocardiosis in marine mammals. Vet. Pathol. 2009, 46, 299–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- St. Leger, J.; Raverty, S.; Mena, A. Pathology of Wildlife and Zoo Animals; Terio, K.A., McAloose, D., St. Leger, J., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 533–568. [Google Scholar]
- Degollada, E.; Domingo, M.; Alonso, J.M.; Alegre, F.; Tello, M.; Lopez, A.; Domínguez, L. Nocardiosis in a striped dolphin (Stenella coeruleoalba). In Proceedings of the third ECS workshop on Cetacean Pathology: Lung Pathology, ECS Newsletter n., Lisbon, Portugal, 14 March 1996; pp. 16–18. [Google Scholar]
- Ito, S.; Hirai, T.; Hamabe, S.; Subangkit, M.; Okabayashi, T.; Goto, Y.; Nishida, S.; Kurita, T.; Yamaguchi, R. Suppurative necrotizing bronchopneumonia caused by Nocardia cyriacigeorgica infection in a stranded striped dolphin (Stenella coeruleoalba) in Japan. J. Vet. Med Sci. 2021, 83, 146–150. [Google Scholar] [CrossRef] [PubMed]
- Pier, A.C.; Takayama, A.K.; Miyahara, A.Y. Cetacean Nocardiosis. J. Wildl. Dis. 1970, 6, 112–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martineau, D.; Lagacé, A.; Béland, P.; Higgins, R.; Armstrong, D.; Shugart, L. Pathology of stranded beluga whales (Delphinapterus leucas) from the St. Lawrence Estuary, Québec, Canada. J. Comp. Pathol. 1988, 98, 287–310. [Google Scholar] [CrossRef]
- Arbelo, M.; de Los Monteros, A.E.; Herráez, P.; Andrada, M.; Sierra, E.; Rodríguez, F.; Jepson, P.D.; Fernandez, A. Pathology and causes of death of stranded cetaceans in the Canary Islands (1999–2005). Dis. Aquat. Org. 2013, 103, 87–99. [Google Scholar] [CrossRef]
- Díaz-Delgado, J.; Fernández, A.; Sierra, E.; Sacchini, S.; Andrada, M.; Vela, A.I.; Óscar, Q.-C.; Paz, Y.; Zucca, D.; Groch, K.; et al. Pathologic findings and causes of death of stranded cetaceans in the Canary Islands (2006–2012). PLoS ONE 2018, 13, e0204444. [Google Scholar] [CrossRef] [Green Version]
- Kuiken, T.; García-Hartmann, M. Dissection techniques and tissue sampling. In Proceedings of the First ECS Workshop on Cetacean Pathology, Leiden, The Netherlands, 13–14 September 1991. [Google Scholar]
- Geraci, J.R.; Lounsbury, V.J. Marine Mammals Ashore: A Field Guide for Strandings, 2nd ed.; National Aquarium in Baltimore: Baltimore, MD, USA, 2005. [Google Scholar]
- Pérez-Sancho, M.; Cerdá, I.; Fernández-Bravo, A.; Domínguez, L.; Figueras, M.J.; Fernández-Garayzábal, J.F.; Vela, A.I. Limited performance of MALDI-TOF for identification of fish Aeromonas isolates at species level. J. Fish Dis. 2018, 41, 1485–1493. [Google Scholar] [CrossRef]
- Vela, A.I.; Collins, M.D.; Lawson, P.A.; García, N.; Domínguez, L.; Fernandez-Garayzabal, J.F. Uruburuella suis gen. nov., sp. nov., isolated from clinical specimens of pigs. Int. J. Syst. Evol. Microbiol. 2005, 55, 643–647. [Google Scholar] [CrossRef] [Green Version]
- Yoon, S.-H.; Ha, S.-M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A taxonomically united database of 16S rRNA gene sequences and whole-genome assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef]
- Sierra, E.; Sanchez, S.; Saliki, J.T.; Blas-Machado, U.; Arbelo, M.; Zucca, D.; Fernández, A. Retrospective study of etiologic agents associated with nonsuppurative meningoencepha-litis in stranded cetaceans in the Canary Islands. J. Clin. Microbiol. 2014, 52, 2390–2397. [Google Scholar] [CrossRef] [Green Version]
- VanDevanter, D.R.; Warrener, P.; Bennett, L.; Schultz, E.R.; Coulter, S.; Garber, R.L.; Rose, T.M. Detection and analysis of diverse herpesviral species by consensus primer PCR. J. Clin. Microbiol. 1996, 34, 1666–1671. [Google Scholar] [CrossRef] [Green Version]
- Saubolle, M.A.; Sussland, D. Nocardiosis: Review of clinical and laboratory experience. J. Clin. Microbiol. 2003, 41, 4497–4501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fellows, G.A.; Kalsi, P.S.; Martin, A.J. Nocardia farcinica brain abscess in a patient without immunocompromise. Br. J. Neurosurg. 2007, 21, 301–303. [Google Scholar] [CrossRef]
- Greene, C. Actinomycosis and nocardiosis. In Infectious Diseases of Dog and Cat, 3rd ed.; Elsevier: St. Louis, MO, USA, 2006; pp. 451–461. [Google Scholar]
- Ribeiro, M.G.; Salerno, T.; De Mattos-Guaraldi, A.L.; Camello, T.C.F.; Langoni, H.; Siqueira, A.K.; Paes, A.C.; Fernandes, M.C.; Lara, G.H.B. Nocardiosis: An overview and additional report of 28 cases in cattle and dogs. Rev. Inst. Med. Trop. São Paulo 2008, 50, 177–185. [Google Scholar] [CrossRef] [Green Version]
- Soto, S.; González, B.; Willoughby, K.; Maley, M.; Olvera, A.; Kennedy, S.; Marco, A.; Domingo, M. Systemic Herpesvirus and Morbillivirus Co-Infection in a Striped Dolphin (Stenella coeruleoalba). J. Comp. Pathol. 2012, 146, 269–273. [Google Scholar] [CrossRef] [PubMed]
- Bawa, B.; Bai, J.; Whitehair, M.; Purvis, T.; Debey, B.M. Bovine abortion associated with Nocardia farcinica. J. Vet. Diagn. Investig. 2010, 22, 108–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown-Elliott, B.A.; Brown, J.M.; Conville, P.S.; Wallace, R.J. Clinical and Laboratory Features of the Nocardia spp. Based on Current Molecular Taxonomy. Clin. Microbiol. Rev. 2006, 19, 259–282. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Huang, A.; Fang, Q. Disseminated nocardiosis caused by Nocardia otitidiscaviarum in an immunocompetent host: A case report and literature review. Exp. Ther. Med. 2016, 12, 3339–3346. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Zhang, Y.; Gong, J.; Jiang, S.; Huang, Y.; Wang, L.; Xu, J.; Qiu, C. CT findings of pulmonary nocardiosis: A report of 9 cases. J. Thorac. Dis. 2017, 9, 4785–4790. [Google Scholar] [CrossRef] [Green Version]
- Engelbrecht, A.; Saad, H.; Gross, H.; Kaysser, L. Natural Products from Nocardia and Their Role in Pathogenicity. Microb. Physiol. 2021, 31, 217–232. [Google Scholar] [CrossRef]
- Derungs, T.; Leo, F.; Loddenkemper, C.; Schneider, T. Treatment of disseminated nocardiosis: A host–pathogen approach with adjuvant interferon gamma. Lancet Infect. Dis. 2021, 21, e334–e340. [Google Scholar] [CrossRef]



{kind=link}
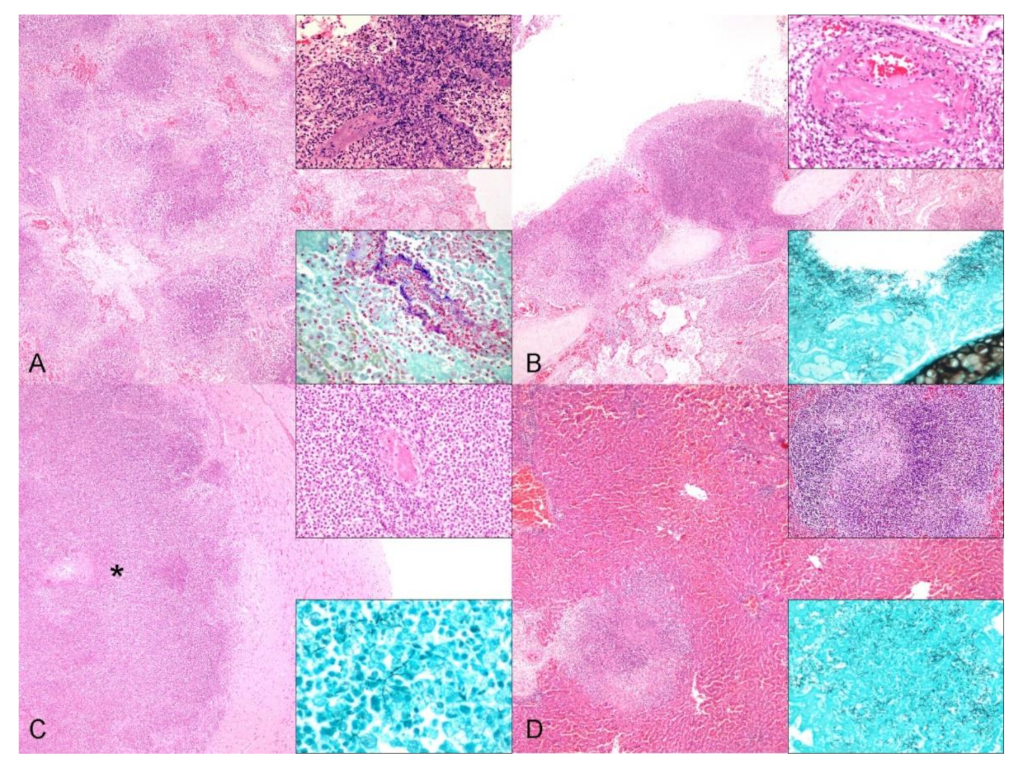{kind=link}
| Case | Case No. | Species | Stranding Location | Gender | Age | BC | DC | ND |
|---|---|---|---|---|---|---|---|---|
| 297_14 | 1 | SD | Almeria | F | adult | 3 | 2 | 19 March 2014 |
| 292_14 | 2 | SD | Cádiz | F | adult | 3 | 2 | 22 April 2014 |
| 34_16 | 3 | SD | Málaga | M | adult | 1 | 2 | 11 November 2015 |
| 148_01 | 4 | SD | Gran Canaria | M | adult | 4 | 2 | 26 November 2001 |
| 1103_20 | 5 | ABD | Gran Canaria | M | Calf | 1 | 2 | 14 June 2020 |
| Gross Findings | Af/Ev (%) |
|---|---|
| Pulmonary pyogranulomatous lymphadenitis (lymphadenomegaly) | 5/5 (100) |
| Pleural, pulmonary, and bronchial pyogranulomas | 5/5 (100) |
| Cerebral abscess | 3/5 (60) |
| Disseminated nodules across liver capsule and/or parenchyma | 3/5 (60) |
| Adrenal gland pyogranulomas | 3/5 (60) |
| Pyogranulomatous pericarditis | 2/5 (40) |
| Peritoneal (serosal) pyogranulomas | 3/5 (60) |
| Endotracheal mucosal pyogranulomas | 2/5 (40) |
| Pharyngeal (tonsil) pyogranulomas | 2/5 (40) |
| Splenic pyogranulomas | 2/5 (40) |
| Mesenteric, tracheobronchial pyogranulomatous lymphadenitis (lymphadenomegaly) | 2/5 (40) |
| Renal pyogranulomas | 2/5 (40) |
| Pancreatic pyogranulomas | 1/5 (25) |
| Hypophyseal pyogranulomas | 1/5 (25) |
| Microscopic Findings | Af/Ev (%) |
|---|---|
| Pyogranulomatous and necrotizing bronchopneumonia and pleuritis | 5/5 (100) |
| Pyogranulomatous and necrotizing pulmonary lymphadenitis | 5/5 (100) |
| Multiorgan vascular vasculitis/fibrinoid necrosis | 5/5 (100) |
| Intralesional filamentous bacteria | 5/5 (100) |
| Suppurative thromboembolic meningoencephalitis with necrosis | 3/5 (60) |
| Suppurative/pyogranulomatous cortical/medullar adrenalitis | 3/5 (60) |
| Pyogranulomatous and necrotizing hepatitis | 3/5 (60) |
| Necrosuppurative splenitis | 2/5 (40) |
| Pyogranulomatous and necrotizing nephritis | 2/5 (40) |
| Necrosuppurative pharyngeal tonsilitis | 2/5 (40) |
| Necrosuppurative pancreatitis | 2/5 (40) |
| Pyogranulomatous tracheobronchial/mesenteric lymphadenitis | 2/5 (40) |
| Fibrinosuppurative and necrotizing pericarditis | 2/5 (40) |
| Necrosuppurative hypophysitis | 1/5 (25) |
| Lymphocytic meningitis with perivascular cuffs and gliosis | 1/5 (25) |
| Pyogranulomatous mielitis | 1/5 (25) |
| Pyogranulomatous (aortic) arteritis | 1/5 (25) |
| Case No. | Lung | Pleura | Liver | Kidney | Spleen | Peritoneum (Serosae) | Heart | Pulmonary (LN) | Adrenal gland | Pharyngeal Tonsil | Pancreas | Trachea | Hypophysis | Brain | Other Lymph Nodes | Organism |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | +++ | ++ | ++ | ? | +++ | ++ | ++ | +++ | +++ | + | - | +++ | +++ | + | + (PS); + (MS) | N. farcinica |
| 2 | +++ | - | - | - | - | + | - | +++ | - | ++ | - | ++ | - | - | - | N. farcinica |
| 3 | +++ | - | ++ | ++ | - | - | - | ++ | ++ | - | - | - | - | - | - | Nocardia sp.* |
| 4 | +++ | ++ | + | +++ | ++ | + | ++a | +++ | ++ | - | +++ | - | - | +++ | - | N. otitidiscaviarum |
| 5 | +++ | - | - | - | - | - | - | +++ | - | - | - | - | - | +++ | +++ (TBr) | N. otitidiscaviarum |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Díaz-Santana, P.; Fernández, A.; Díaz-Delgado, J.; Vela, A.I.; Domínguez, L.; Suárez-Santana, C.; Puig-Lozano, R.; Fernández-Maldonado, C.; Sierra, E.; Arbelo, M. Nocardiosis in Free-Ranging Cetaceans from the Central-Eastern Atlantic Ocean and Contiguous Mediterranean Sea. Animals 2022, 12, 434. https://doi.org/10.3390/ani12040434
Díaz-Santana P, Fernández A, Díaz-Delgado J, Vela AI, Domínguez L, Suárez-Santana C, Puig-Lozano R, Fernández-Maldonado C, Sierra E, Arbelo M. Nocardiosis in Free-Ranging Cetaceans from the Central-Eastern Atlantic Ocean and Contiguous Mediterranean Sea. Animals. 2022; 12(4):434. https://doi.org/10.3390/ani12040434
Chicago/Turabian StyleDíaz-Santana, Pablo, Antonio Fernández, Josue Díaz-Delgado, Ana Isabel Vela, Lucas Domínguez, Cristian Suárez-Santana, Raquel Puig-Lozano, Carolina Fernández-Maldonado, Eva Sierra, and Manuel Arbelo. 2022. "Nocardiosis in Free-Ranging Cetaceans from the Central-Eastern Atlantic Ocean and Contiguous Mediterranean Sea" Animals 12, no. 4: 434. https://doi.org/10.3390/ani12040434