
Effects of Leptin, Growth Hormone and Photoperiod on Pituitary SOCS-3 Expression in Sheep


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Characteristics
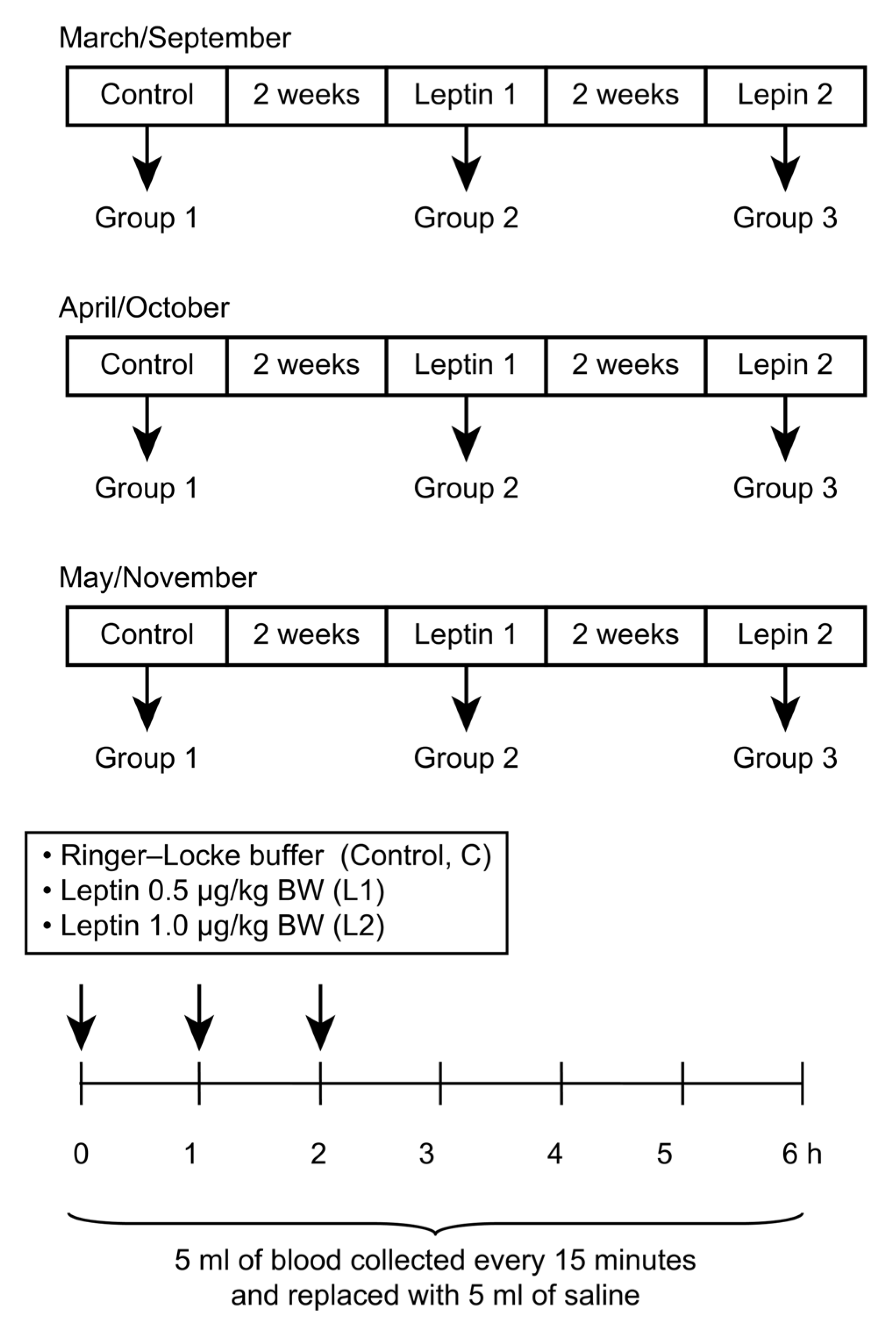
2.2. Experiment I—The In Vivo Influence of Central Leptin Treatment on GH Concentrationin Sheep
2.2.1. Animals and Treatments
2.2.2. Treatments
2.2.3. RIA for GH
2.2.4. Data Analysis and Statistical Tests
2.3. Experiment II—The In Vitro Influence of Leptin and GH on SOCS-3 Transcript Levels in Adenohypophysis Explants
2.3.1. Animals and Treatments
2.3.2. Molecular Analysis
2.3.3. Molecular Data Analysis
3. Results
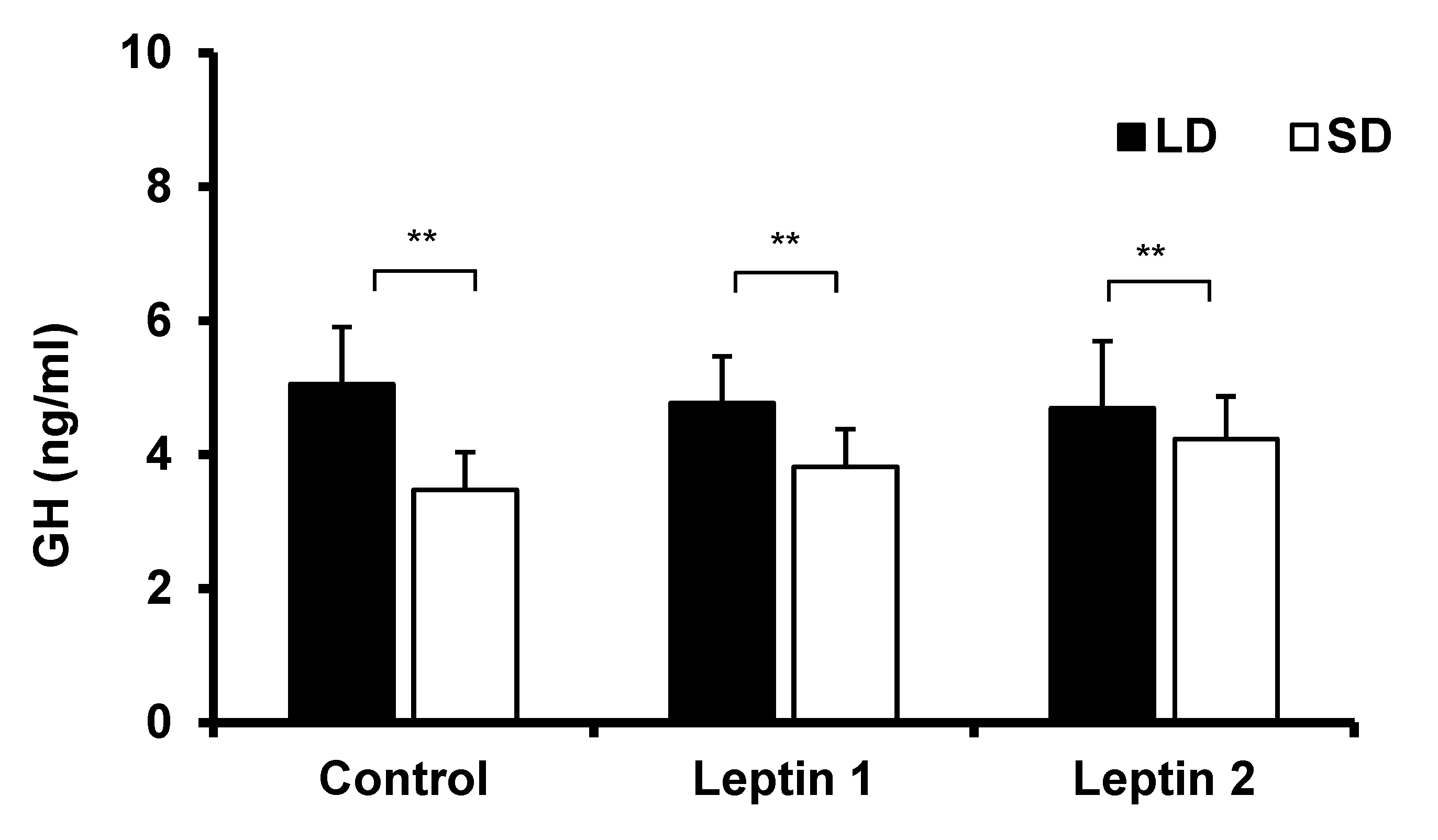
3.1. Experiment I—The In Vivo Influence of Central Leptin Treatment on GH Concentrations in Sheep
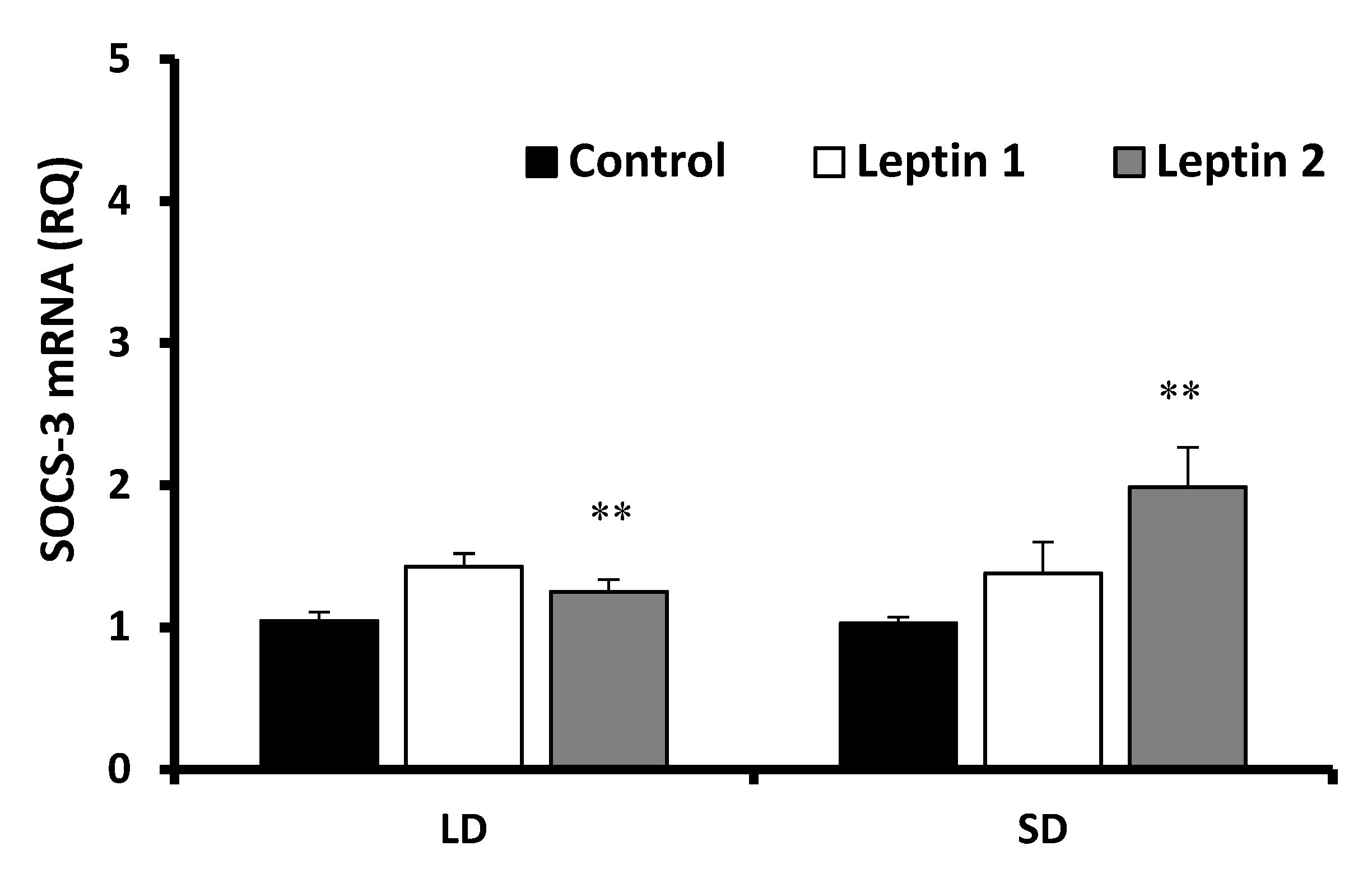
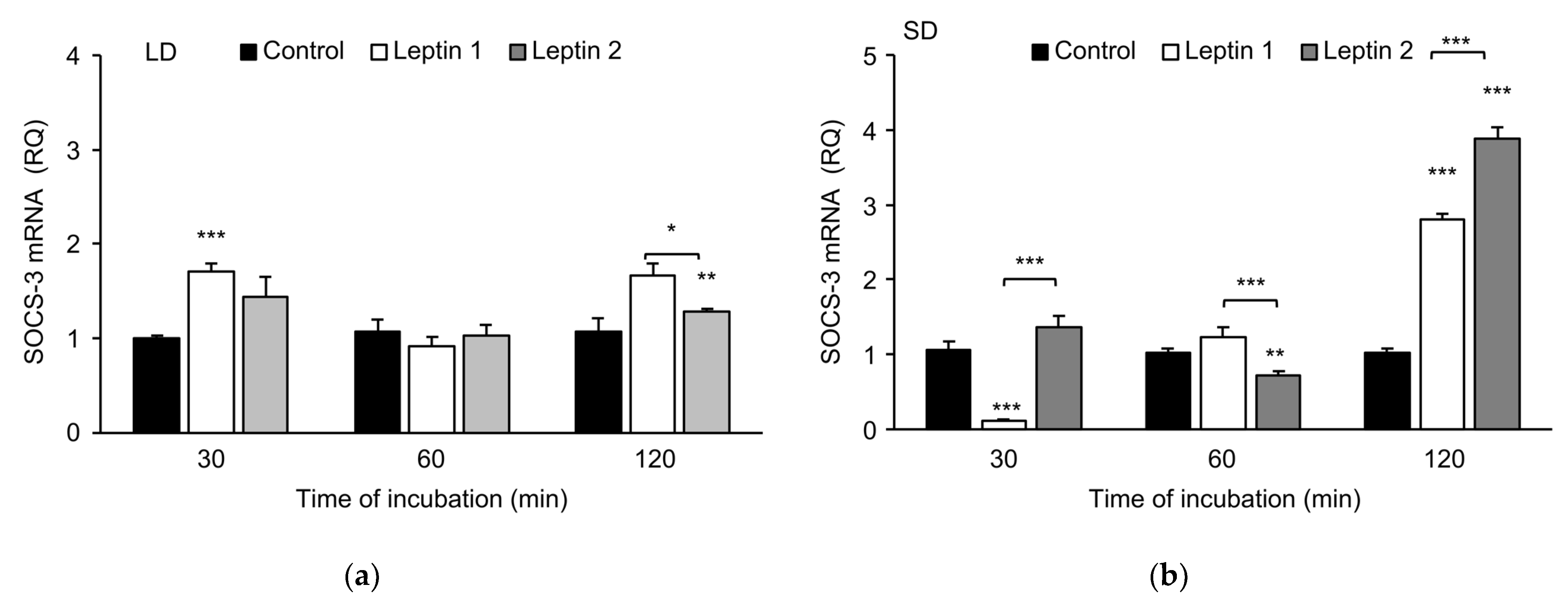
3.2. Experiment II—The In Vitro Influence of Leptin and GH on SOCS-3 Transcript Levels in Adenohypophysis Explants
3.2.1. Leptin Effects
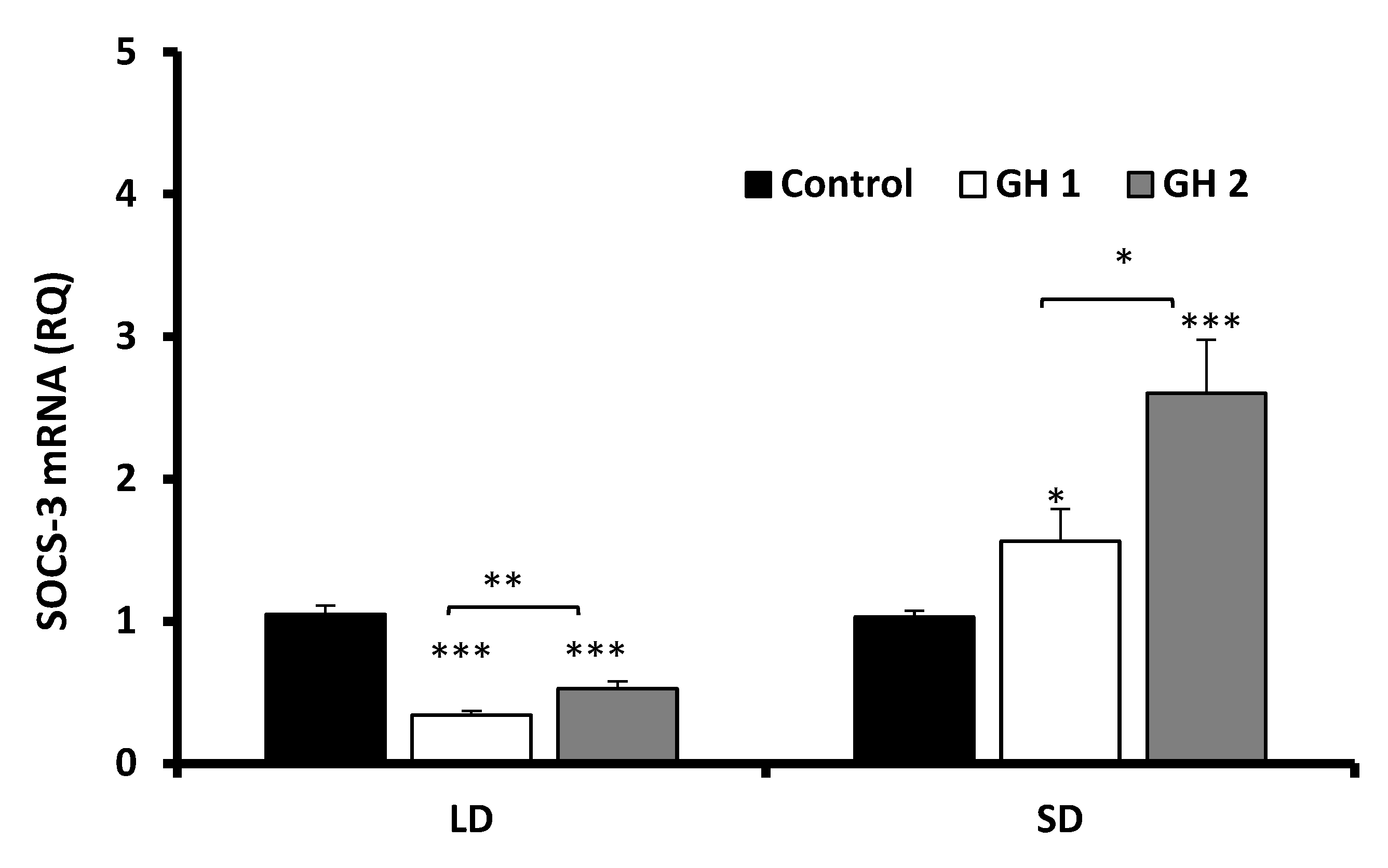
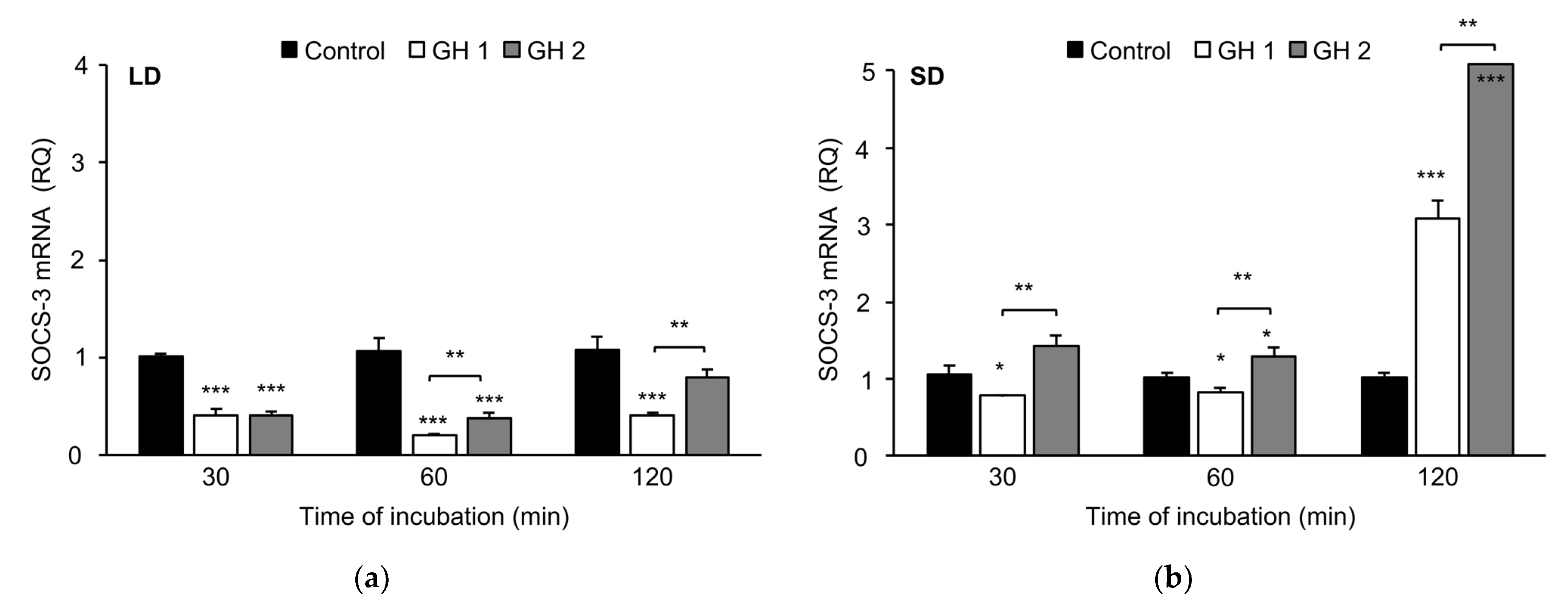
3.2.2. Growth Hormone Effects
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Molik, E.; Misztal, T.; Romanowicz, K.; Zieba, D.A. The effects of melatonin on prolactin and growth hormone secretion in ewes under different photoperiods, during the early post partum period. Small Rumin. Res. 2010, 94, 137–141. [Google Scholar] [CrossRef]
- Iqbal, J.; Pompolo, S.; Considine, R.V.; Clarke, I.J. Localization of leptin receptor-like immunoreactivity in the corticotropes, somatotropes, and gonadotropes in the ovine anterior pituitary. Endocrinology 2000, 141, 1515–1520. [Google Scholar] [CrossRef] [PubMed]
- Cocchi, D.; Colonna, V.D.G.; Bagnasco, M.; Bonacci, D.; Muller, E.E. Leptin regulates GH secretion in the rat by acting on GHRH and somatostatinergic functions. J. Endocrinol. 1999, 162, 95–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roh, S.G.; Nie, G.Y.; Loneragan, K.; Gertler, A.; Chen, C. Direct modification of somatotrope function by long-term leptin treatment of primary cultured ovine pituitary cells. Endocrinology 2001, 142, 5167–5171. [Google Scholar] [CrossRef] [PubMed]
- Accorsi, P.A.; Gamberoni, M.; Isani, G.; Govoni, N.; Viggiani, R.; Monari, M.; De Ambrogi, M.; Munno, A.; Tamanini, C.; Seren, E. Leptin does not seem to influence glucose uptake by bovine mammary explants. J. Physiol. Pharmacol. 2005, 56, 689–698. [Google Scholar]
- Kirsz, K.; Szczesna, M.; Molik, E.; Misztal, T.; Wojtowicz, A.K.; Zieba, D.A. Seasonal changes in the interactions among leptin, ghrelin, and orexin in sheep. J. Anim. Sci. 2012, 90, 2524–2531. [Google Scholar] [CrossRef]
- Henry, B.A.; Goding, J.W.; Alexander, W.S.; Tilbrook, A.J.; Canny, B.J.; Dunshea, F.; Rao, A.; Mansell, A.; Clarke, I.J. Central administration of leptin to ovariectomized ewes inhibits food intake without affecting the secretion of hormones from the pituitary gland: Evidence for a dissociation of effects on appetite and neuroendocrine function. Endocrinology 1999, 140, 1175–1182. [Google Scholar] [CrossRef]
- Henry, B.A.; Goding, J.W.; Tilbrook, A.J.; Dunshea, F.R.; Clarke, I.J. Intracerebroventricular infusion of leptin elevates the secretion of luteinising hormone without affecting food intake in long-term food-restricted sheep, but increases growth hormone irrespective of bodyweight. J. Endocrinol. 2001, 168, 67–77. [Google Scholar] [CrossRef] [Green Version]
- Morrison, C.D.; Daniel, J.A.; Holmberg, B.J.; Djiane, J.; Raver, N.; Gertler, A.; Keisler, D.H. Central infusion of leptin into well-fed and undernourished ewe lambs: Effects on feed intake and serum concentrations of growth hormone and luteinizing hormone. J. Endocrinol. 2001, 168, 317–324. [Google Scholar] [CrossRef] [Green Version]
- Adams, T.E.; Hansen, J.A.; Starr, R.; Nicola, N.A.; Hilton, D.J.; Billestrup, N. Growth hormone preferentially induces the rapid, transient expression of SOCS-3, a novel inhibitor of cytokine receptor signaling. J. Biol. Chem. 1998, 273, 1285–1287. [Google Scholar] [CrossRef] [Green Version]
- Zieba, D.A.; Biernat, W.; Szczesna, M.; Kirsz, K.; Barć, J.; Misztal, T. Changes in expression of the genes for the leptin signaling in hypothalamic-pituitary selected areas and endocrine responses to long-term manipulation in body weight and resistin in ewes. Int. J. Mol. Sci. 2020, 21, 4238. [Google Scholar] [CrossRef] [PubMed]
- Biernat, W.; Szczęsna, M.; Kirsz, K.; Zieba, D.A. Seasonal and nutritional fluctuations in the mRNA levels of the short form of the leptin receptor (LRa) in the hypothalamus and anterior pituitary in resistin-treated sheep. Animals 2021, 11, 2451. [Google Scholar] [CrossRef] [PubMed]
- Bjørbæk, C.; Lavery, H.J.; Bates, S.H.; Olson, R.K.; Davis, S.M.; Flier, J.S.; Myers, M.G. SOCS3 mediates feedback inhibition of the leptin receptor via Tyr985. J. Biol. Chem. 2000, 275, 40649–40657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tups, A.; Ellis, C.; Moar, K.M.; Logie, T.J.; Adam, C.L.; Mercer, J.G.; Klingenspor, M. Photoperiodic regulation of leptin sensitivity in the Siberian hamster, Phodopus sungorus, is reflected in arcuate nucleus SOCS-3 (suppresor of cytokine signaling) gene expression. Endocrinology 2004, 145, 1185–1193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bjørbæk, C.; Elmquist, J.K.; Frantz, J.D.; Shoelson, S.E.; Flier, J.S. Identification of SOCS-3 as a potential mediator of central leptin resistance. Mol. Cell 1998, 1, 619–625. [Google Scholar] [CrossRef]
- Adam, C.L.; Mercer, J.G. Appetite regulation and seasonality: Implications for obesity. Proc. Nutr. Soc. 2004, 63, 413–419. [Google Scholar] [CrossRef] [Green Version]
- Zieba, D.A.; Szczesna, M.; Klocek-Gorka, B.; Molik, E.; Misztal, T.; Williams, G.L.; Romanowicz, K.; Stepien, E.; Keisler, D.H.; Murawski, M. Seasonal effects of central leptin infusion on secretion of melatonin and prolactin and on SOCS-3 gene expression in ewes. J. Endocrinol. 2008, 198, 147–155. [Google Scholar] [CrossRef] [Green Version]
- Szczesna, M.; Zieba, D.A.; Klocek-Gorka, B.; Misztal, T.; Stepien, E. Seasonal effects of central leptin infusion and prolactin treatment on pituitary SOCS-3 gene expression in ewes. J. Endocrinol. 2011, 208, 81–88. [Google Scholar] [CrossRef] [Green Version]
- Zieba, D.A.; Amstalden, M.; Morton, S.; Gallino, J.L.; Edwards, J.F.; Harms, P.G.; Williams, G.L. Effects of leptin on basal and GHRH-stimulated GH secretion from the bovine adenohypophysis are dependent upon nutritional status. J. Endocrinol. 2003, 178, 83–89. [Google Scholar] [CrossRef] [Green Version]
- Molik, E.; Zieba, D.A.; Misztal, T.; Romanowicz, K.; Wszola, M.; Wierzchos, E.; Nowakowski, M. The role of orexin A in the control of prolactin and growth hormone secretions in sheep—In vitro study. J. Physiol. Pharmacol. 2008, 59, 91–100. [Google Scholar]
- Russel, A.J.F.; Doney, J.M.; Gunn, R.G. Subjective assessment of body fat in live sheep. J. Agric. Sci. 1969, 72, 451–454. [Google Scholar] [CrossRef]
- Norms. Nutrient Requirements for Cattle and Sheep in the Traditional System; IZ: Krakow, Poland, 1993. [Google Scholar]
- Biernat, W.; Kirsz, K.; Szczesna, M.; Zieba, D.A. Resistin regulates reproductive hormone secretion from the ovine adenohypophysis depending on season. Domest. Anim. Endocrinol. 2018, 65, 95–100. [Google Scholar] [CrossRef]
- Traczyk, W.; Przekop, F. Methods of investigation of the function of the hypothalamus and hypophysis in chronic experiment in sheep. Acta Physiol. Pol. 1963, 14, 217–226. [Google Scholar]
- Zieba, D.A.; Amstalden, M.; Maciel, M.N.; Keisler, D.H.; Raver, N.; Gertler, A.; Williams, G.L. Divergent effects of leptin on luteinizing hormone and insulin secretion are dose dependent. Exp. Biol. Med. 2003, 228, 325–330. [Google Scholar] [CrossRef]
- Zieba, D.A.; Amstalden, M.; Morton, S.; Maciel, M.N.; Keisler, D.H.; Williams, G.L. Regulatory roles of leptin at the hypothalamic-hypophyseal axis before and after sexual maturation in cattle. Biol. Reprod. 2004, 71, 804–812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adam, C.L.; Findlay, P.A.; Miller, D.W. Blood–brain leptin transport and appetite and reproductive neuroendocrine responses to intracerebroventricular leptin injection in sheep: Influence of photoperiod. Endocrinology 2006, 147, 4589–4598. [Google Scholar] [CrossRef] [Green Version]
- Dvorak, P.; Becka, S.; Krejci, P.; Chrpova, M. Radioimmunoassay of bovine growth hormone. Radiochem. Radioanal. Lett. 1978, 34, 160–166. [Google Scholar]
- Zieba, D.A.; Klocek, B.; Williams, G.L.; Romanowicz, R.; Boliglowa, L.; Wozniak, M. In vitro evidence that leptin suppresses melatonin secretion during long days and stimulates its secretion during short days in seasonal breeding ewes. Domest. Anim. Endocrinol. 2007, 33, 358–365. [Google Scholar] [CrossRef]
- Denis, R.G.; Bing, C.; Brocklehurst, S.; Harrold, J.A.; Vernon, R.G.; Williams, G. Diurnal changes in hypothalamic neuropeptide and SOCS-3 expression: Effects of lactation and relationship with serum leptin and food intake. J. Endocrinol. 2004, 183, 173–181. [Google Scholar] [CrossRef] [Green Version]
- Baskin, D.G.; Breininger, J.F.; Schwartz, M.W. SOCS-3 expression in leptin-sensitive neurons of the hypothalamus of fed and fasted rats. Regul. Pept. 2000, 92, 9–15. [Google Scholar] [CrossRef]
- Amstalden, M.; Spencer, T.E.; Harms, P.G.; Williams, G.L. Expression of leptin receptor and suppressor of cytokine signaling-3 genes in adenohypophysis of normal-fed and fasted cows. Reprod. Biol. 2005, 5, 237–245. [Google Scholar] [PubMed]
- Kirsz, K.; Szczesna, M.; Dudek, K.; Bartlewski, P.M.; Zieba, D.A. Influence of season and nutritional status on the direct effects of leptin, orexin-A and ghrelin on luteinizing hormone and growth hormone secretion in the ovine pituitary explant model. Domest. Anim. Endocrinol. 2014, 48, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Misztal, T.; Romanowicz, K.; Barcikowski, B. Melatonin modulation of the daily prolactin secretion in intact and ovariectomized ewes. Neuroendocrinology 1999, 69, 105–112. [Google Scholar] [CrossRef]
- Rousseau, K.; Atcha, Z.; Cagampang, F.R.A.; Le Rouzic, P.; Stirland, J.A.; Ivanov, T.R.; Ebling, F.J.P.; Klingenspor, M.; Loudon, A.S.I. Photoperiodic regulation of leptin resistance in the seasonally breeding siberian hamster (Phodopussungorus). Endocrinology 2002, 143, 3083–3095. [Google Scholar] [CrossRef] [PubMed]
- Klingenspor, M.; Niggemann, H.; Heldmaier, G. Modulation of leptin sensitivity by short photoperiod acclimation in the Djungarian hamster, Phodopussungorus. J. Comp. Physiol. B 2000, 170, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Mercer, J.G.; Moar, K.M.; Ross, A.W.; Hoggard, N.; Morgan, P.J. Photoperiod regulates arcuate nucleus POMC, AGRP, and leptin receptor mRNA in Siberian hamster hypothalamus. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2000, 278, R271–R281. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5′-3′) | Probe Sequence (5′-3′) | Amplicon Size | GenBank Accession Number |
|---|---|---|---|---|
| CPH | CGGCTCCCAGTTCTTCATCA | FAM-CGTTCCGACTCCGC-MGB | 64 bp | D14074 |
| ACTACGTGCTTCCCATCCAAA | ||||
| SOCS-3 | CCTCAAGACCTTCAGCTCCAA | FAM-AGCGAGTACCAGCTGG-MGB | 68 bp | NM_174466 |
| CTTGCGCACTGCGTTCAC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zieba, D.A.; Szczesna, M.; Kirsz, K.; Biernat, W. Effects of Leptin, Growth Hormone and Photoperiod on Pituitary SOCS-3 Expression in Sheep. Animals 2022, 12, 403. https://doi.org/10.3390/ani12030403
Zieba DA, Szczesna M, Kirsz K, Biernat W. Effects of Leptin, Growth Hormone and Photoperiod on Pituitary SOCS-3 Expression in Sheep. Animals. 2022; 12(3):403. https://doi.org/10.3390/ani12030403
Chicago/Turabian StyleZieba, Dorota Anna, Malgorzata Szczesna, Katarzyna Kirsz, and Weronika Biernat. 2022. "Effects of Leptin, Growth Hormone and Photoperiod on Pituitary SOCS-3 Expression in Sheep" Animals 12, no. 3: 403. https://doi.org/10.3390/ani12030403