
Effects of Dietary Multienzyme Complex Supplementation on Growth Performance, Digestive Capacity, Histomorphology, Blood Metabolites and Hepatic Glycometabolism in Snakehead (Channa argus)

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of Diets
2.2. Animals and Sampling Procedures
2.3. Analysis of Serum Biochemistry Index
2.4. Glycometabolism and Digestive Enzymes Analysis
2.5. Quantitative Real-Time PCR Analysis
2.6. Analysis of Hepatopancreas Morphology and Dorsal Muscle Component
2.6.1. Hematoxylin and Eosin (H&E) Staining
2.6.2. Oil Red O Staining
2.7. Electron Microscopy Examinations
2.8. Gas Chromatography−Mass Spectrometry (GC/MS) Analysis
2.9. Statistical Analysis
3. Results
3.1. Growth Performance, Feed Intake, and Morphological Parameters
3.2. Serum Biochemical Parameters
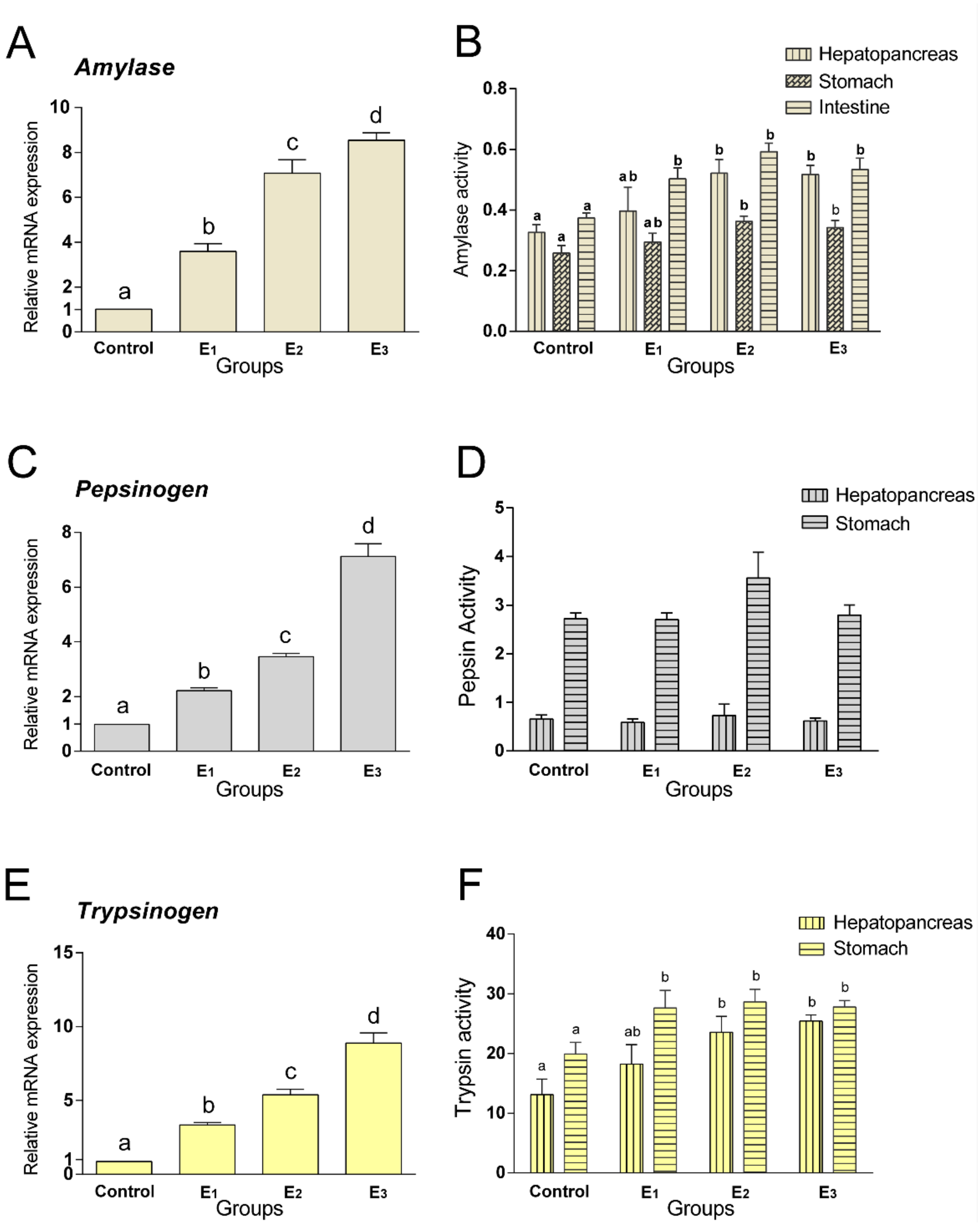
3.3. Digestive Enzymes Activities and Related Genes Expression
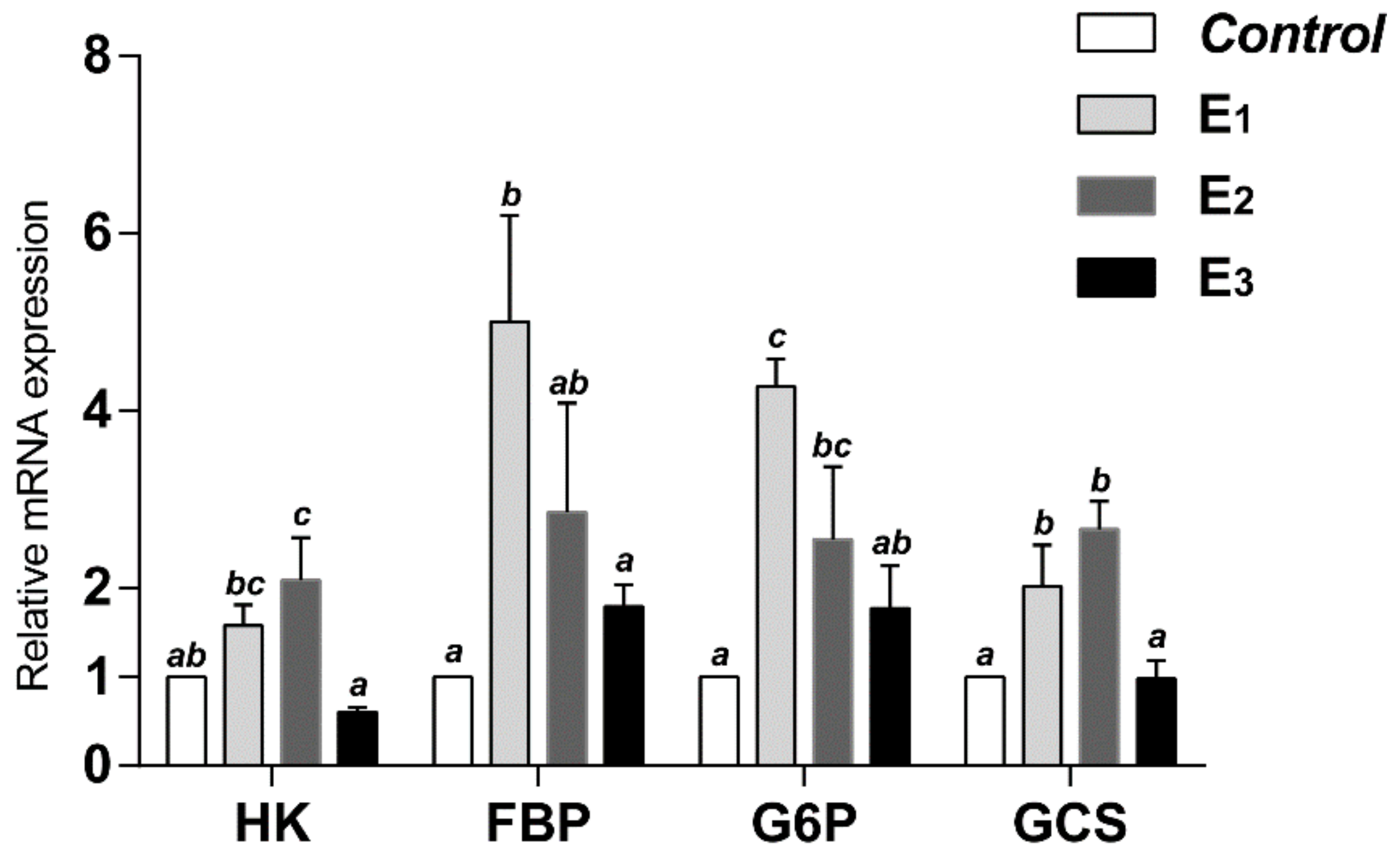
3.4. Glucose Metabolic Key Enzymes Activities and Relative mRNA Expression
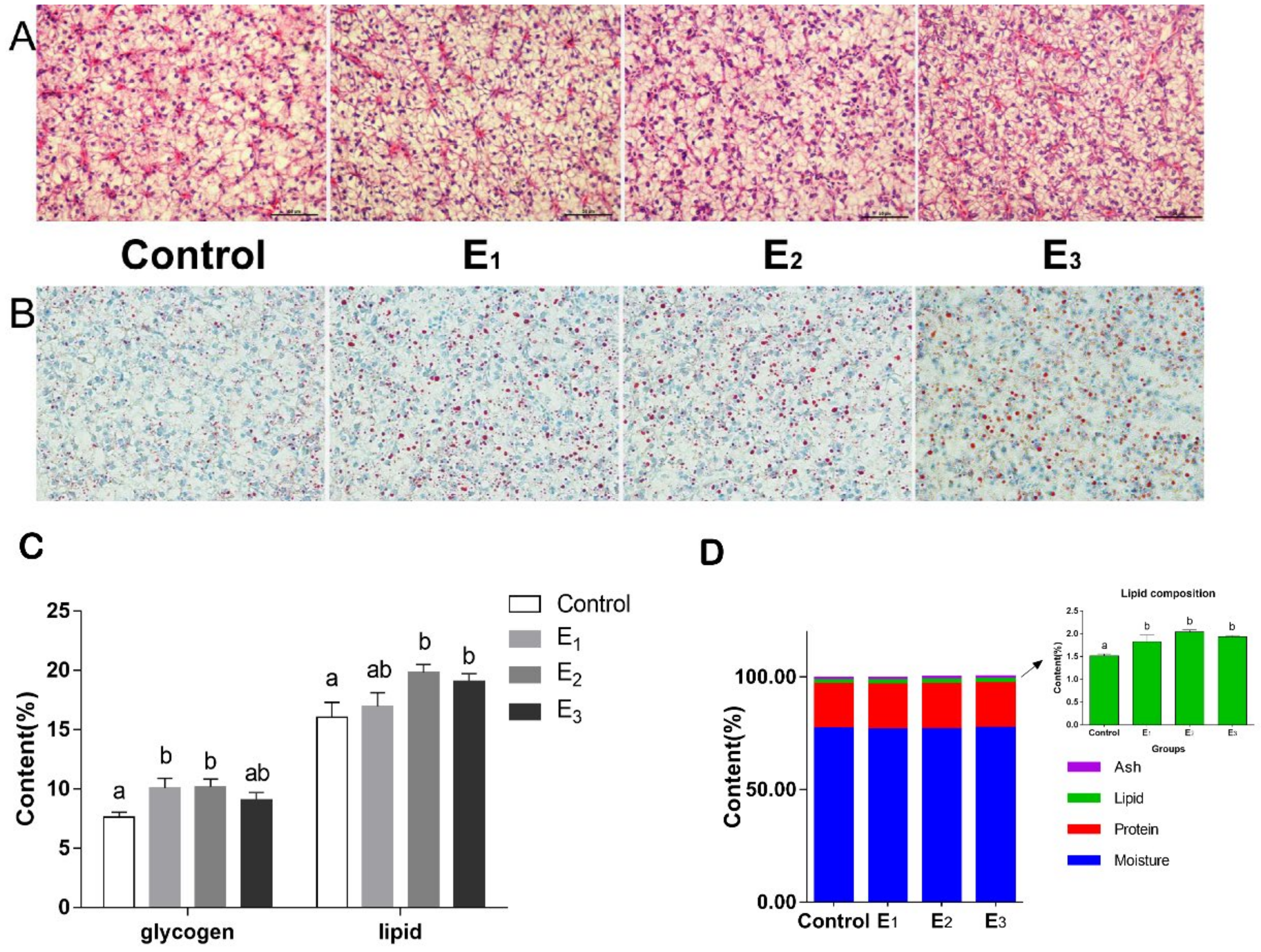
3.5. Hepatopancreas Morphology, and the Composition of Hepatopancreas and Dorsal Muscle
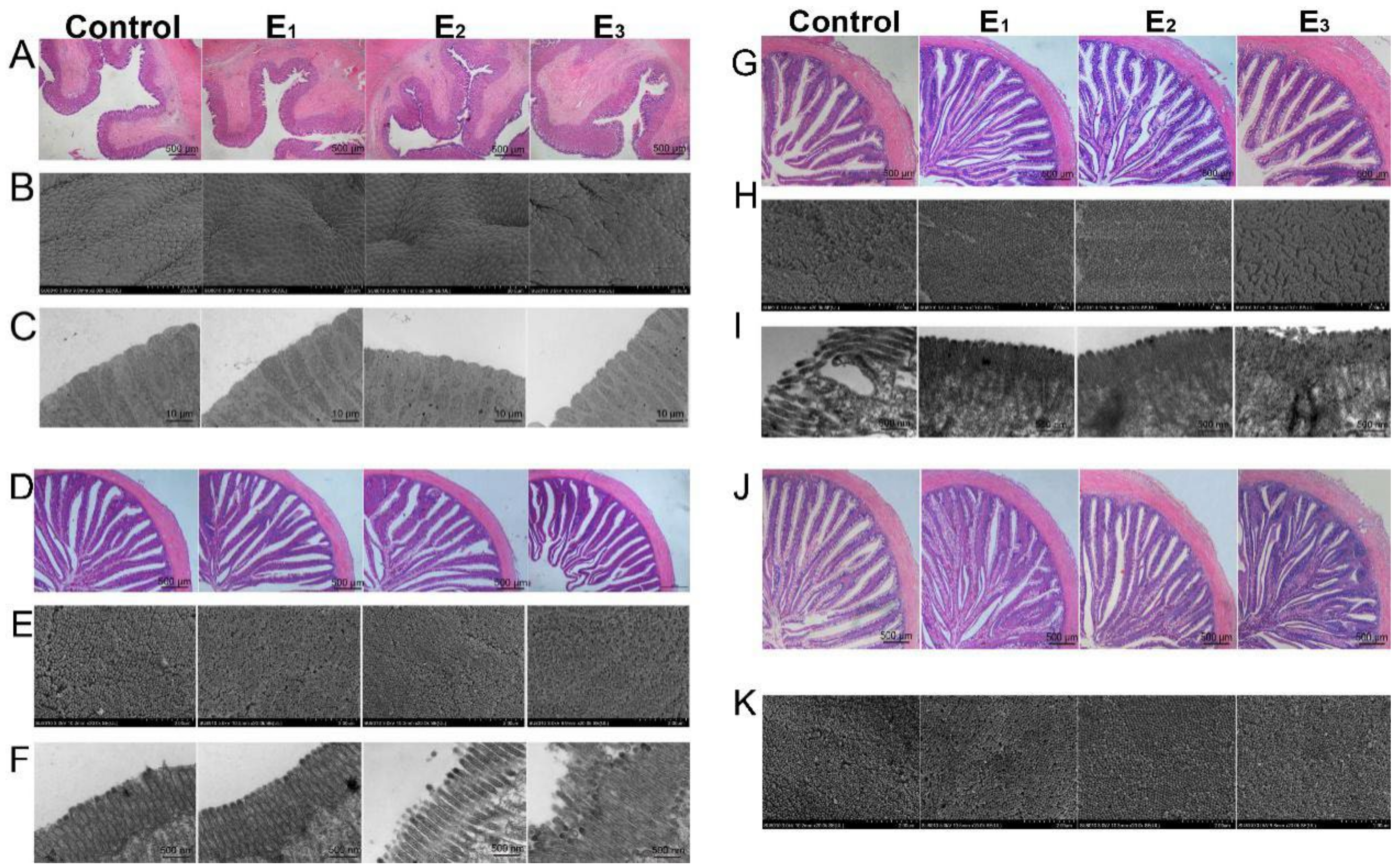
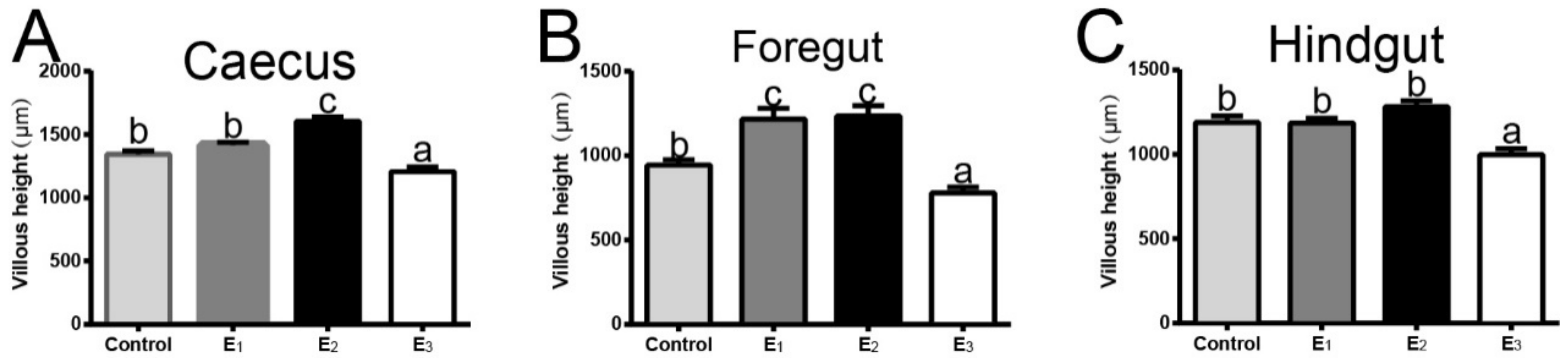
3.6. Gastrointestinal Morphology and Integrity
3.7. GC-MS Statistical Analysis and Discriminate Metabolites
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Muntaziana, M.P.A.; Amin, S.M.N.; Rah, M.A.; Rahim, A.A.; Marimuthu, K. Present culture status of the endangered snakehead, Channa striatus (Bloch, 1793). Asian J. Anim. Vet. Adv. 2013, 8, 369–375. [Google Scholar] [CrossRef]
- Sagada, G.; Chen, J.; Shen, B.; Huang, A.; Sun, L.; Jiang, J.; Jin, C. Optimizing protein and lipid levels in practical diet for juvenile northern snakehead fish (Channa argus). Anim. Nutr. 2017, 3, 156–163. [Google Scholar] [CrossRef] [PubMed]
- Dai, B.; Hou, Y.; Yong, H.; Qian, L. Effects of multienzyme complex and probiotic supplementation on the growth performance, digestive enzyme activity and gut microorganisms composition of snakehead (Channa argus). Aquac. Nutr. 2019, 25, 15–25. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.; Li, Z.; Xu, A.; Zheng, D.; Ye, Y.; Wang, Z. Effects of exogenous multienzyme complex supplementation in diets on growth performance, digestive enzyme activity and non-specific immunity of the Japanese seabass, Lateolabrax japonicus. Aquac. Nutr. 2020, 26, 306–315. [Google Scholar] [CrossRef]
- Komar, A.H.A.; Yaser, K. Effect of Different Feed Additives on Growth Performance and Production in Livestock. Int. J. Agric. For. 2019, 9, 16–31. [Google Scholar]
- Ravindran, V.; Son, J. Feed enzyme technology: Present status and future developments. Recent Pat. Food Nutr. Agric. 2011, 3, 102–109. [Google Scholar]
- Goda, A.M.A.; Mabrouk, A.H.H.; Wafa, E.H.; El-Afifi, T.M. Effect of using baker’s yeast and exogenous digestive enzymes as growth promoters on growth, fed utilization and hematological indices of Nile tilapia, Oreochromis niloticus fingerlings. J. Agric. Sci. Technol. 2012, 2, 15–28. [Google Scholar]
- Carvalho, V.V.; Paulino, M.F.; Detmann, E.; Chizzotti, M.L.; Martins, L.S.; Silva, A.G.; Lopes, S.A.; Moura, F.H. Effects of supplements containing different additives on nutritional and productive performance of beef cattle grazing tropical grass. Trop. Anim. Health Prod. 2017, 49, 983–988. [Google Scholar] [CrossRef]
- Ding, X.; Yao, L.; Hou, Y.; Hou, Y.; Wang, G.; Fan, J.; Qian, L. Effects of different carbohydrate levels in puffed feed on digestive tract morphological function and liver tissue structure of snakeheads (Channa argus). Aquac. Res. 2020, 51, 557–568. [Google Scholar] [CrossRef]
- Gracia, M.; Aranibar, M.; Lazaro, R.; Medel, P.; Mateos, G. Alphaamylase supplementation of broiler diets based on corn. Poult. Sci. 2003, 82, 436–442. [Google Scholar] [CrossRef]
- Tang, J.; James, M.N.G.; Hsu, I.N.; Jenkins, J.A.; Blundell, T.L. Structural evidence for gene duplication in the evolution of the acid proteases. Nature 1978, 271, 618–621. [Google Scholar] [CrossRef] [PubMed]
- Torres-Pitarch, A.; Hermans, D.; Manzanilla, E.G.; Bindelle, J.; Everaert, N.; Beckers, Y.; Torrallardonae, D.; Bruggemanc, G.; Gardinerf, G.E.; Lawlora, P.G. Effect of feed enzymes on digestibility and growth in weaned pigs: A systematic review and me-ta-analysis. Anim. Feed. Sci. Technol. 2017, 233, 145–159. [Google Scholar] [CrossRef]
- Kutlu, H.R.; Saber, S.N.; Kutay, H.; Celik, L.; Uzun, Y.; Toy, N.; Kutlu, M.; Yucelt, O.; Burgut, A.; Thiery, P.; et al. Effect of multi-enzyme produced by a single fungus on growth performance and some carcass parameters of broiler chicks fed on maize-soya based diets. Kafkas. Univ. Vet. Fak. Derg. 2019, 25, 221–230. [Google Scholar]
- Kumar, V.; Sinha, A.K.; Makkar, H.P.S.; De Boeck, G.; Becker, K. Phytate and phytase in fish nutrition. J. Anim. Physiol. Anim. Nutr. 2012, 96, 335–364. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Li, Z.; Gan, L.; Fan, H.; Guo, Y. Dietary supplemental Kluyveromyces marxianus alters the serum metabolite profile in broiler chickens. Food Funct. 2018, 9, 3776–3787. [Google Scholar] [CrossRef]
- Ibero-Baraibar, I.; Romo-Hualde, A.; Gonzalez-Navarro, C.; Zulet, M.A.; Martinez, J.A. The urinary metabolomic profile following the intake of meals supplemented with a cocoa extract in middle-aged obese subjects. Food Funct. 2016, 7, 1924–1931. [Google Scholar] [CrossRef] [PubMed]
- AOAC Association of Analytical Chemists. Official Methods of Analysis, 18th ed.; AOAC: Washington, DC, USA, 2007. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Kholif, A.E.; Abdo, M.M.; Anele, U.Y.; El-Sayed, M.M.; Morsy, T.A. Saccharomyces cerevisiae does not work synergistically with exogenous enzymes to enhance feed utilization, ruminal fermentation and lactational performance of Nubian goats. Livest. Sci. 2017, 206, 17–23. [Google Scholar] [CrossRef]
- Liu, W.; Wu, J.P.; Li, Z.; Duan, Z.Y.; Wen, H. Effects of dietary coated protease on growth performance, feed utilization, nutrient apparent digestibility, intestinal and hepatopancreas structure in juvenile Gibel carp (Carassius auratus gibelio). Aquac. Nutr. 2018, 24, 47–55. [Google Scholar] [CrossRef]
- Ghomi, M.R.; Shahriari, R.; Langroudi, H.F.; Nikoo, M.; von Elert, E. Effects of exogenous dietary enzyme on growth, body com-position, and fatty acid profiles of cultured great sturgeon Huso huso fingerlings. Aquac. Int. 2012, 20, 249–254. [Google Scholar] [CrossRef]
- Hlophe-Ginindza, S.N.; Moyo, N.A.G.; Ngambi, J.W.; Ncube, I. The effect of exogenous enzyme supplementation on growth perfor-mance and digestive enzyme activities in Oreochromis mossambicus fed kikuyu-based diets. Aquac. Res. 2016, 47, 3777–3787. [Google Scholar] [CrossRef]
- Lin, S.; Mai, K.; Tan, B. Effects of exogenous enzyme supplementation in diets on growth and feed utilization in tilapia, Oreochromis niloticus × O-aureus. Aquac. Res. 2007, 38, 1645–1653. [Google Scholar] [CrossRef]
- Owsley, W.F.; Orr, D.E.; Tribble, L.F. Effects of the age and diet on the development of the pancreas and synthesis and secretion of Pancreatic enzymes in young pig. J. Anim. Sci. 1986, 63, 479–504. [Google Scholar] [CrossRef] [PubMed]
- Xie, R.-T.; Amenyogbe, E.; Chen, G.; Huang, J.-S. Effects of feed fat level on growth performance, body composition and serum biochemical indices of hybrid grouper (Epinephelus fuscoguttatus × Epinephelus polyphekadion). Aquaculture 2021, 530, 735813. [Google Scholar] [CrossRef]
- Chew, S.F.; Wong, M.Y.; Tam, W.L.; Ip, Y.K. The snakehead Channa asiatica accumulates alanine during aerial exposure, but is incapable of sustaining locomotory activities on land through partial amino acid catabolism. J. Exp. Biol. 2003, 206, 693–704. [Google Scholar] [CrossRef] [Green Version]
- Azm, F.R.A.; Kong, F.; Tan, Q.; Zhu, Y.; Yu, H.; Yao, J.; Luo, Z. Effects of replacement of dietary rapeseed meal by distiller’s dried grains with solubles (DDGS) on growth performance, muscle texture, health and expression of muscle-related genes in grass carp (Ctenopharyngodon idellus). Aquaculture 2021, 533, 736169. [Google Scholar]
- Osman, L.M.; Ayres, J.G.; Garden, C.; Reglitz, K.; Lyon, J.; Douglas, J.G. A randomised trial of home energy efficiency improvement in the homes of elderly COPD patients. Eur. Respir. J. 2010, 35, 303–309. [Google Scholar] [CrossRef] [Green Version]
- Han, C.; Wei, Y.; Wang, X.; Ba, C.; Shi, W. Protective effect of Salvia miltiorrhiza polysaccharides on liver injury in chickens. Poul. Sci. 2019, 98, 3496–3503. [Google Scholar] [CrossRef]
- Diaz-Rosales, P.; Kanashiro, E.; Castro, C.; Magalhães, R.; Oliva-Teles, A.; Peres, H. Improved digestibility of plant feedstuff based diets in sea bass (Dicentrarchus labrax) with exogenous enzymes. Aquaculture 2014, 24, 333–334. [Google Scholar]
- Li, J.S.; Li, J.L.; Wu, T.T. Effects of non-starch polysaccharides enzyme, phytase and citric acid on activities of endogenous digestive enzymes of tilapia (Oreochromis niloticus × Oreochromis aureus). Aquac. Nutr. 2009, 15, 415–420. [Google Scholar] [CrossRef]
- Jiang, T.-T.; Feng, L.; Liu, Y.; Jiang, W.-D.; Jiang, J.; Li, S.-H.; Tang, L.; Kuang, S.-Y.; Zhou, X.-Q. Effects of exogenous xylanase supplementation in plant protein-enriched diets on growth performance, intestinal enzyme activities and microflora of juvenile Jian carp (Cyprinus carpio var. Jian). Aquac. Nutr. 2014, 20, 632–645. [Google Scholar] [CrossRef]
- Zhou, Y.; Yuan, X.; Liang, X.-F.; Fang, L.; Li, J.; Guo, X.; Bai, X.; He, S. Enhancement of growth and intestinal flora in grass carp: The effect of exogenous cellulase. Aquaculture 2013, 416, 1–7. [Google Scholar] [CrossRef]
- Gharaei, A.; Ghaffari, M.; Keyvanshokooh, S.; Akrami, R. Changes in metabolic enzymes, cortisol and glucose concentrations of Beluga (Huso huso) exposed to dietary methylmercury. Fish Physiol. Biochem. 2011, 37, 485–493. [Google Scholar] [CrossRef] [PubMed]
- Enes, P.; Panserat, S.; Kaushik, S.; Oliva-Teles, A. Nutritional regulation of hepatic glucose metabolism in fish. Fish Physiol. Biochem. 2009, 35, 519–539. [Google Scholar] [CrossRef] [PubMed]
- Enes, P.; Peres, H.; Couto, A.; Oliva-Teles, A. Growth performance and metabolic utilization of diets including starch, dextrin, maltose or glucose as carbohydrate source by gilthead sea bream (Sparus aurata) juveniles. Fish Physiol. Biochem. 2010, 36, 903–910. [Google Scholar] [CrossRef] [PubMed]
- Borrebaek, B.; Waagbo, R.; Christophersen, B.; Tranulis, M.A.; Hemre, G.I. Adaptable hexokinase with low-affinity for glucose in the liver of Atlantic Salmon (Salmo salar). Comp. Biochem. Physiol. 1993, 106, 833–836. [Google Scholar] [CrossRef]
- Jin, J.; Medale, F.; Kamalam, B.S.; Aguirre, P.; Véron, V.; Panserat, S. Comparison of glucose and lipid metabolic gene expressions between fat and lean lines of rainbow trout after a glucose load. PLoS ONE 2014, 9, e105548. [Google Scholar] [CrossRef] [Green Version]
- Fang, J.; Wang, H.; Fang, L.; Li, B.; Zhu, W.; Zhou, Y.; Mei, J.; Yuan, X.; Liu, L.W.; He, Y. Effects of heat shock treatment on glucose metabolism in grass carp (Ctenopharyngodon idellus) juveniles. Aquaculture 2014, 422, 211–217. [Google Scholar] [CrossRef]
- Yusuf, A.; Huang, X.; Chen, N.; Li, S.; Apraku, A.; Wang, W.; Davidad, M.A. Growth and metabolic responses of juvenile largemouth bass (Micropterus salmoides) to dietary vitamin c supplementation levels. Aquaculture 2021, 534, 736243. [Google Scholar] [CrossRef]
- Wei, Z.; Ma, J.; Pan, X.; Mu, H.; Li, J.; Shen, T.J.; Zhang, W.B.; Mai, K.S. Dietary hydroxyproline improves the growth and muscle quality of large yellow croaker Larimichthys crocea. Aquaculture 2016, 464, 497–504. [Google Scholar] [CrossRef]
- Zhao, Y.; Li, J.-Y.; Yin, L.; Feng, L.; Liu, Y.; Jiang, W.-D.; Wu, P.; Zhao, J.; Chen, D.-F.; Zhou, X.-Q.; et al. Effects of dietary glutamate supplementation on flesh quality, antioxidant defense and gene expression related to lipid metabolism and myogenic regulation in Jian carp (Cyprinus carpio var. Jian). Aquaculture 2019, 502, 212–222. [Google Scholar] [CrossRef]
- Sorgeloos, P. Effect of water exchange, salinity regime, stocking density and diets on growth and survival of domesticated black tiger shrimp Penaeus monodon (Fabricius,1798) reared in sand-based recirculating systems. Aquaculture 2012, 338, 253–259. [Google Scholar]
- Hernández, M.; López, M.; Alvarez, A.; Ferrandini, E.; García, B.G.; Garridob, M.D. Sensory, physical, chemical and microbiological changes in aquacultured meagre (Argyrosomus regius) fillets during ice storage. Food. Chem. 2009, 114, 237–245. [Google Scholar] [CrossRef]
- Refaey, M.M.; Li, D.; Tian, X.; Zhang, Z.; Zhang, X.; Li, L.; Tang, R. High stocking density alters growth performance, blood biochemistry, intestinal histology, and muscle quality of channel catfish Ictalurus punctatus. Aquaculture 2018, 492, 73–81. [Google Scholar] [CrossRef]
- Bakke-Mckellep, A.M.; Press, C.M.; Baeverfjord, G.; Krogdahl, Å.; Landsverk, T. Changes in immune and enzyme histochemical phenotypes of cells in the intestinal mucosa of Atlantic salmon, Salmo salar L., with soybean meal-induced enteritis. J. Fish Dis. 2000, 23, 115–127. [Google Scholar] [CrossRef]
- Li, P.; Yin, Y.-L.; Li, D.; Kim, S.W.; Wu, G. Amino acids and immune function. Br. J. Nutr. 2007, 98, 237. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Mai, K.; Trushenski, J.; Wu, G. New developments in fish amino acid nutrition: Towards functional and environmentally oriented aquafeeds. Amino Acids 2009, 37, 43–53. [Google Scholar] [CrossRef]
- Hou, Y.; Yao, K.; Yin, Y.; Wu, G. Endogenous synthesis of amino acids limits growth, lactation and reproduction of animals. Adv. Nutr. 2016, 7, 331–342. [Google Scholar] [CrossRef] [Green Version]
- Fang, Y.Z.; Yang, S.; Wu, G. Free radicals, antioxidants, and nutrition. Nutrition 2002, 8, 872–879. [Google Scholar] [CrossRef]
- Shamushaki, V.A.J.; Kasumyan, A.O.; Abedian, A.; Abtahi, B. Behavioural responses of the Persian sturgeon (Acipenser persicus) juveniles to free amino acid solutions. Mar. Freshw. Behav. Physiol. 2007, 40, 219–224. [Google Scholar] [CrossRef]
- Riley, W.W.; Higgs, D.A.; Dosanjh, B.S.; Eales, J.G. Influence of dietary arginine and glycine content on thyroid function and growth of juvenile rainbow trout, Oncorhynchus mykiss (Walbaum). Aquac. Nutr. 1996, 2, 235–242. [Google Scholar] [CrossRef]
- Wu, G.; Bazer, F.W.; Dai, Z.; Li, D.; Wang, J.; Wu, Z. Amino acid nutrition in animals: Protein Synthesis and Beyond. Annu. Rev. Anim. Biosci. 2014, 2, 387. [Google Scholar] [CrossRef] [PubMed]
- Debelle, L.; Tamburro, A.M. Elastin: Molecular description and function. Int. J. Biochem. Cell Biol. 1999, 31, 261–272. [Google Scholar] [CrossRef]
- Omura, Y.; Inagaki, M. Immunocytochemical localization of taurine in the fish retina under light and dark adaptations. Amino Acids 2000, 19, 593–604. [Google Scholar] [CrossRef] [PubMed]
- Gaylord, T.G.; Barrows, F.T.; Teague, A.M.; Johansen, K.A.; Overturf, K.E.; Shephed, B. Supplementation of taurine and methionine to all-plant protein diets for rainbow trout (Oncorhynchus mykiss). Aquaculture 2007, 269, 514–524. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients % | Nutrient Levels *** | ||||
|---|---|---|---|---|---|
| Corn starch | 5 | Soybean oil | 3 | Crude protein | 43.17 |
| Wheat flour | 15 | Fish oil | 2 | Ether extract | 13.5 |
| Fish meal | 40 | Beer yeast | 3 | Crude fiber | 1.76 |
| Soybean meal | 10.5 | Bentonite | 2 | Ash | 6.65 |
| Peanut meal | 9 | Phosphatide oil | 3 | ME (MJ/kg) | 2.49 |
| Rapeseed meal | 1 | Ca(H2PO4)2 | 1.5 | DE (MJ/kg) | 2.13 |
| SVO * | 3 | Premix ** | 2 | GE (MJ/kg) | 21.53 |
| Total | 100 | ||||
| Items | Control | E1 | E2 | E3 | SEM | p Value |
|---|---|---|---|---|---|---|
| IBW, g | 69.77 | 69.83 | 69.49 | 69.97 | 0.149 | 0.755 |
| FBW, g | 220.28 a | 229.31 ab | 238.71 b | 236.34 b | 2.719 | 0.041 |
| WGR, % | 215.72 a | 228.53 ab | 243.51 b | 237.80 b | 4.054 | 0.006 |
| FI, % | 2.04 | 2.06 | 1.97 | 2.03 | 0.165 | 0.292 |
| SGR, % day−1 | 1.92 a | 1.98 b | 2.06 c | 2.03 bc | 0.017 | <0.01 |
| CF, g cm−3 | 1.47 | 1.43 | 1.47 | 1.54 | 0.021 | 0.271 |
| VSI, % | 8.96 | 9.28 | 9.12 | 8.94 | 0.162 | 0.887 |
| HSI, % | 2.04 | 2.34 | 2.38 | 2.41 | 0.103 | 0.529 |
| ISI, % | 76.74 | 77.31 | 78.16 | 75.83 | 1.035 | 0.894 |
| FCR | 1.18 | 1.21 | 1.11 | 1.12 | 0.020 | 0.038 |
| SR, % | 100 | 97.33 | 98.67 | 97.33 | 0.513 | 0.202 |
| Items | Control | E1 | E2 | E3 | SEM | p Value |
|---|---|---|---|---|---|---|
| GLU (mmol/L) | 18.56 | 17.41 | 16.01 | 15.59 | 0.579 | 0.260 |
| TP (g/L) | 37.28 | 37.42 | 40.87 | 42.34 | 1.032 | 0.217 |
| ALB (g/L) | 17.35 | 19.13 | 17.97 | 18.34 | 0.051 | 0.683 |
| BUN (mmol/L) | 16.48 a | 15.33 ab | 13.68 b | 14.63 ab | 0.712 | 0.025 |
| ALT (U/L) | 5.20 a | 4.52 ab | 3.45 b | 5.83 a | 0.544 | 0.044 |
| AST (U/L) | 8.90 a | 5.86 c | 4.70 c | 7.60 b | 0.781 | 0.048 |
| TG (mmol/L) | 2.28 a | 1.80 ab | 1.25 b | 1.82 ab | 0.178 | 0.020 |
| TC (mmol/L) | 6.05 | 5.59 | 5.10 | 6.03 | 0.254 | 0.521 |
| Items | Control | E1 | E2 | E3 | SEM | p Value |
|---|---|---|---|---|---|---|
| HK, U/gprot | 8.57 a | 10.93 ab | 13.24 b | 12.16 b | 0.620 | 0.026 |
| PFK, U/mg | 2.94 | 3.76 | 3.59 | 3.62 | 0.212 | 0.570 |
| PK, U/gprot | 49.47 | 42.96 | 42.82 | 46.13 | 2.713 | 0.395 |
| PC, U/mgprot | 36.09 | 25.10 | 35.16 | 29.29 | 2.289 | 0.299 |
| PEPCK, U/mg | 0.901 | 0.824 | 0.863 | 1.025 | 0.076 | 0.837 |
| FBP, nmol/min/mgprot | 0.900 b | 1.071 b | 1.848 a | 1.012 b | 0.138 | 0.038 |
| G6P, nmol/min/mgprot | 23.97 a | 37.35 b | 34.56 b | 33.21 b | 1.869 | 0.043 |
| G6PDH, U/mgprot | 1.551 | 1.852 | 1.735 | 1.392 | 0.094 | 0.352 |
| GCS, U/mgprot | 0.501 a | 0.824 b | 0.941 b | 0.830 b | 0.063 | 0.045 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ding, X.; Nie, X.; Yuan, C.; Jiang, L.; Ye, W.; Qian, L. Effects of Dietary Multienzyme Complex Supplementation on Growth Performance, Digestive Capacity, Histomorphology, Blood Metabolites and Hepatic Glycometabolism in Snakehead (Channa argus). Animals 2022, 12, 380. https://doi.org/10.3390/ani12030380
Ding X, Nie X, Yuan C, Jiang L, Ye W, Qian L. Effects of Dietary Multienzyme Complex Supplementation on Growth Performance, Digestive Capacity, Histomorphology, Blood Metabolites and Hepatic Glycometabolism in Snakehead (Channa argus). Animals. 2022; 12(3):380. https://doi.org/10.3390/ani12030380
Chicago/Turabian StyleDing, Xiaoqing, Xinzheng Nie, Chunchun Yuan, Lai Jiang, Wenxin Ye, and Lichun Qian. 2022. "Effects of Dietary Multienzyme Complex Supplementation on Growth Performance, Digestive Capacity, Histomorphology, Blood Metabolites and Hepatic Glycometabolism in Snakehead (Channa argus)" Animals 12, no. 3: 380. https://doi.org/10.3390/ani12030380