
Thyroid Hormones in Hemolymph of Red King Crabs from the Barents Sea

 , ,
, ,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
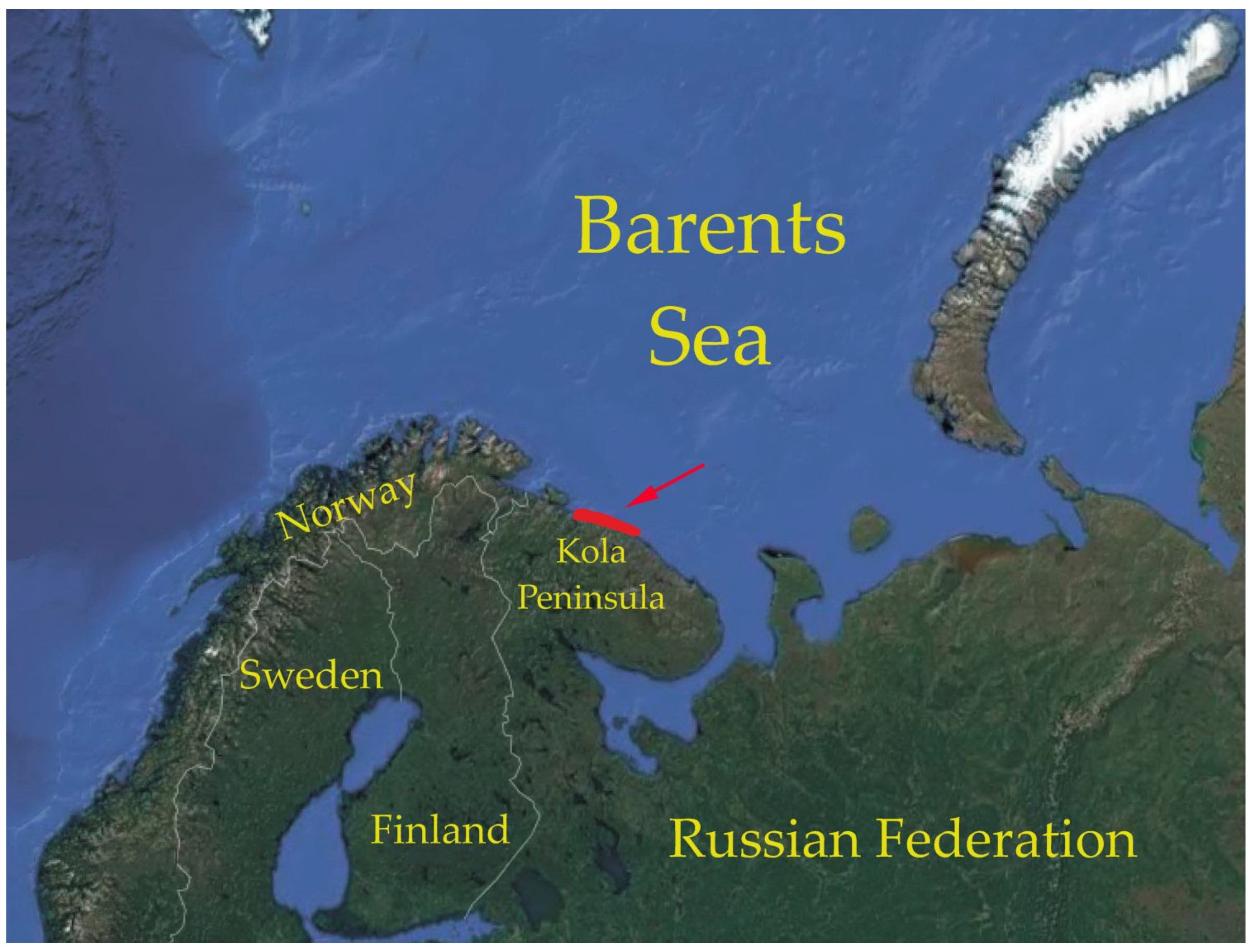
2. Materials and Methods
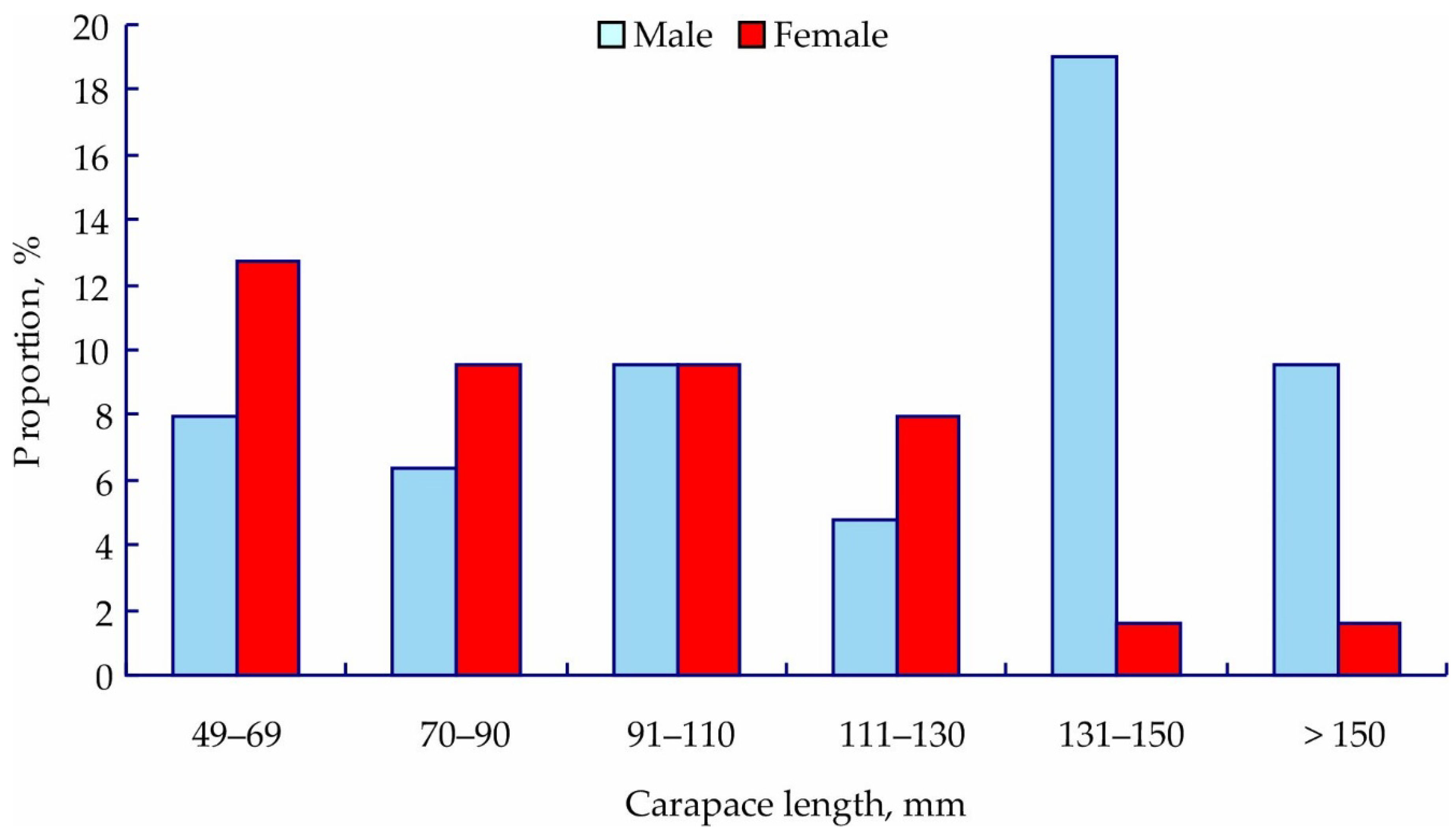
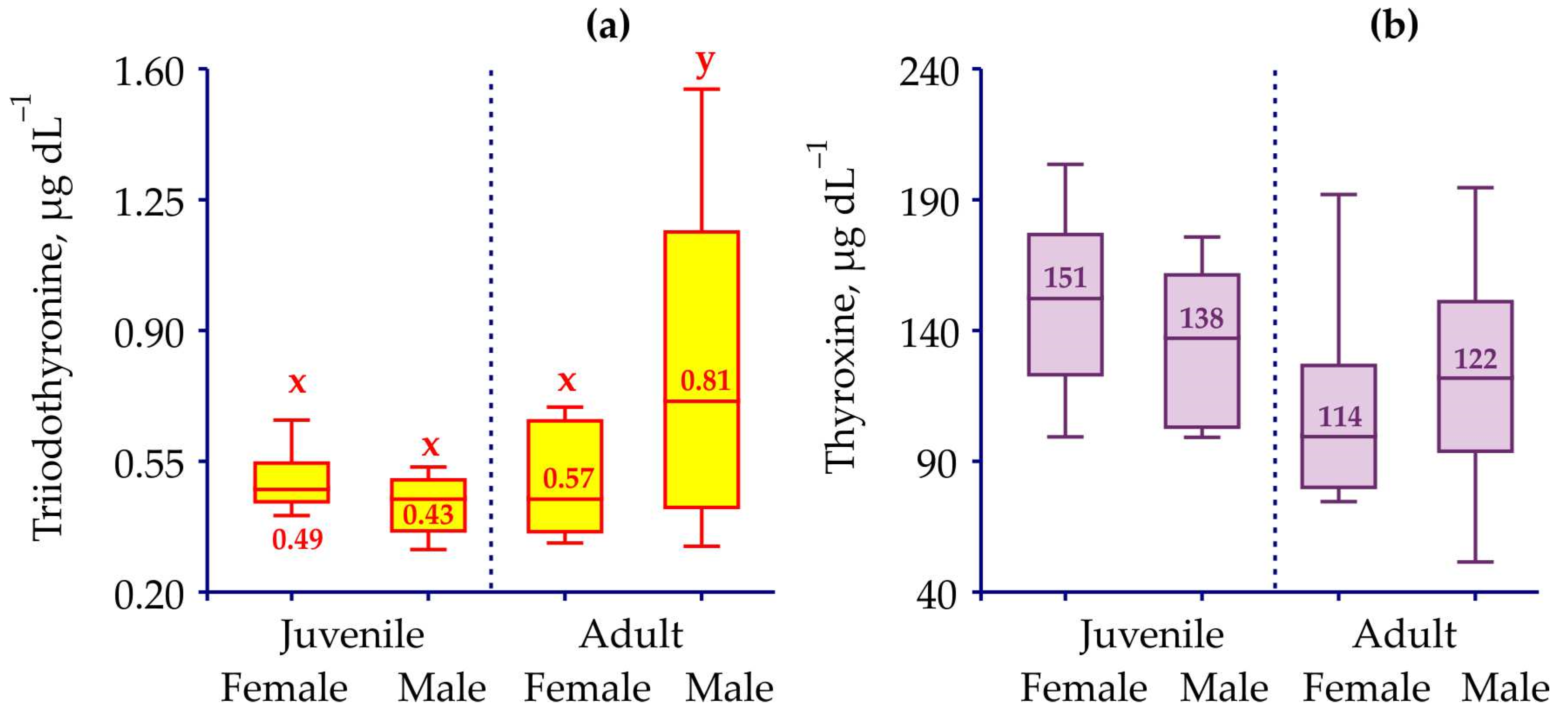
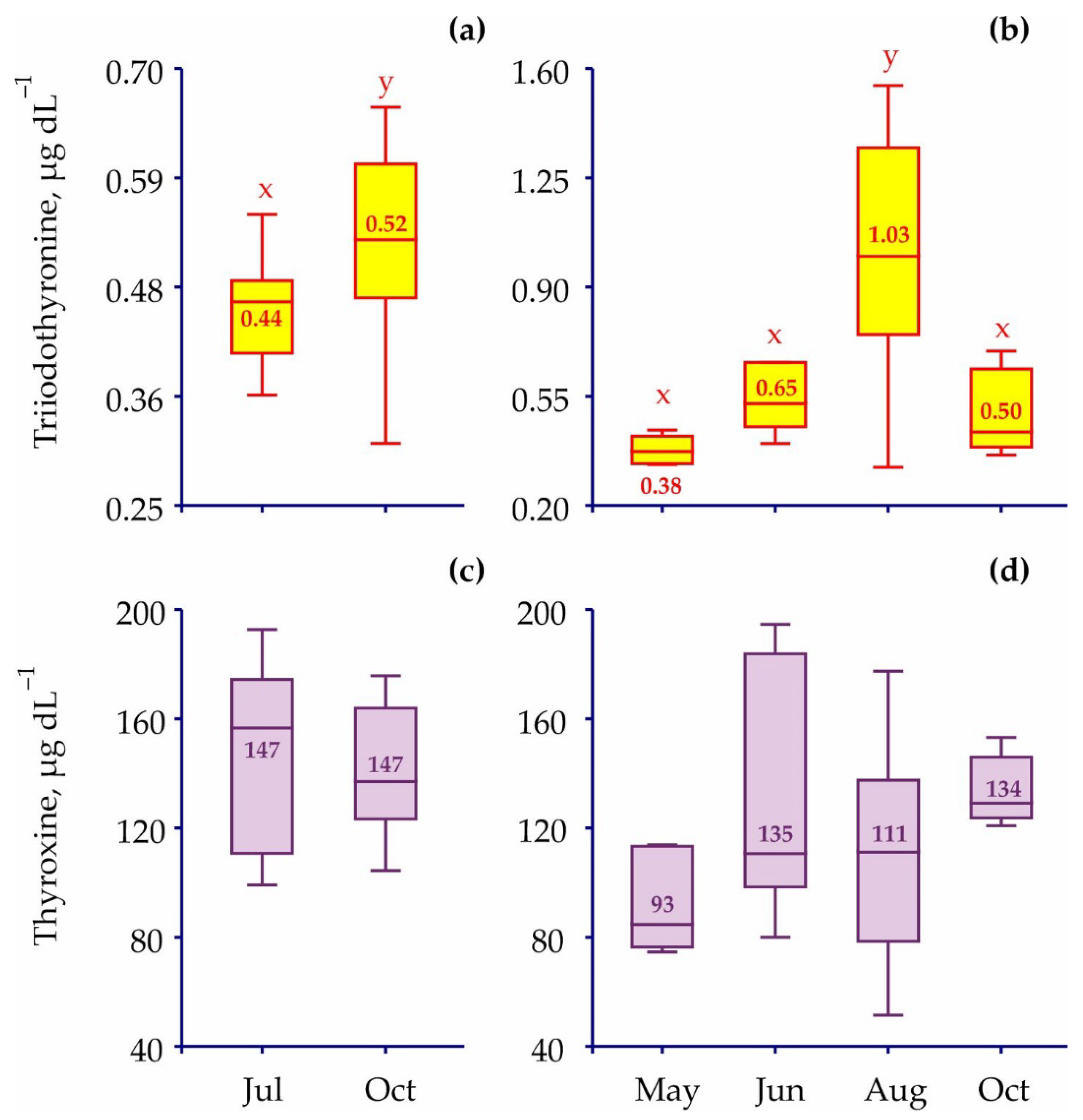
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hulbert, A.J. Thyroid hormones and their effects: A new perspective. Biol. Rev. 2000, 75, 519–631. [Google Scholar] [CrossRef] [Green Version]
- Choksi, N.Y.; Jahnke, G.D.; Hilaire, C.S.; Shelby, M. Role of thyroid hormones in human and laboratory animal reproductive health. Birth Defects Res. B 2003, 68, 479–491. [Google Scholar] [CrossRef]
- Mullur, R.; Liu, Y.-Y.; Brent, G.A. Thyroid hormone regulation of metabolism. Physiol. Rev. 2014, 94, 355–382. [Google Scholar] [CrossRef] [Green Version]
- Sainath, S.B.; André, A.; Castro, L.F.C.; Santos, M.M. The evolutionary road to invertebrate thyroid hormone signaling: Perspectives for endocrine disruption processes. Compar. Biochem. Physiol. C 2019, 223, 124–138. [Google Scholar] [CrossRef]
- Liotta, L.; Bionda, A.; Quartuccio, M.; De Nardo, F.; Visalli, R.; Fazio, E. Thyroid and lipidic profiles in nicastrese goats (Capra hircus) during pregnancy and postpartum period. Animals 2021, 11, 2386. [Google Scholar] [CrossRef]
- Lall, S.P.; Kaushik, S.J. Nutrition and metabolism of minerals in fish. Animals 2021, 11, 2711. [Google Scholar] [CrossRef]
- Crockford, S.J. Evolutionary roots of iodine and thyroid hormones in cell-cell signaling. Integr. Comp. Biol. 2009, 49, 155–166. [Google Scholar] [CrossRef] [Green Version]
- Laudet, V. The origins and evolution of vertebrate metamorphosis. Curr. Biol. 2011, 21, R726–R737. [Google Scholar] [CrossRef] [Green Version]
- Eales, J.G. Iodine metabolism and thyroid-related functions in organisms lacking thyroid follicles: Are thyroid hormones also vitamins? Proc. Soc. Exp. Biol. Med. 1997, 214, 302–317. [Google Scholar] [CrossRef]
- Heyland, A.; Price, D.A.; Bodnarova-Buganova, M.; Moroz, L.L. Thyroid hormone metabolism and peroxidase function in two non-chordate animals. J. Exp. Zool. B Mol. Dev. Evol. 2006, 306, 551–566. [Google Scholar] [CrossRef]
- Iromo, H.; Zairin, M., Jr.; Agus, M.S.; Manalu, W. Thyroxine distribution in the hemolymph, hepatopancreas, ovary, sponge, and larvae of female mud crabs (Scylla serrata) during ovarian maturation. J. Crustac. Biol. 2014, 34, 760–763. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.; Xu, F.; Qu, T.; Zhang, R.; Li, L.; Que, H.; Zhang, G. Identification of thyroid hormones and functional characterization of thyroid hormone receptor in the Pacific oyster Crassostrea gigas provide insight into evolution of the thyroid hormone system. PLoS ONE 2015, 10, e0144991. [Google Scholar] [CrossRef]
- Lustrino, D.; Silva, A.C.M.; Araujo, I.G.; Tunholi, V.M.; Tunholi-Alves, V.M.; Castro, R.N.; Carvalho, D.P.; Pinheiro, J.; Marassi, M.P. Evidence of the presence of thyroid hormones in Achatina fulica snails. An. Acad. Bras. Cienc. 2017, 89, 2181–2188. [Google Scholar] [CrossRef] [Green Version]
- Iromo, H.; Zairin, M., Jr.; Agus, M.S.; Manalu, W. Effectivy of thyroxine hormone suplementation in the ovarian maturation of females mud crab (Scylla serrata). Pak. J. Biotechnol. 2014, 11, 79–86. [Google Scholar]
- Iromo, H.; Zairin, M., Jr.; Agus, M.S.; Manalu, W. Suplementation doses thyroxine hormone of broodstock mud crab (Scylla serrata) during ovarian maturation. J. Aquac. Res. Dev. 2015, 6, 12. [Google Scholar]
- Iromo, H.; Jabarsyah, A.; Awaluddin. Reproduction of females mud crab (Scylla serrata) with thyroxine hormone supplementation in traditional ponds from north Borneo Indonesia. Int. J. Fish. Aquat. Stud. 2018, 6, 378–381. [Google Scholar]
- Britayev, T.A.; Rzhavsky, A.V.; Pavlova, L.V.; Dvoretskij, A.G. Studies on impact of the alien red king crab (Paralithodes camtschaticus) on the shallow water benthic communities of the Barents Sea. J. Appl. Ichthyol. 2010, 26, 66–73. [Google Scholar] [CrossRef]
- Oug, E.; Cochrane, S.; Sundet, J.H.; Norling, K.; Nilsson, H.C. Effects of the invasive red king crab (Paralithodes camtschaticus) on soft-bottom fauna in Varangerfjorden, Northern Norway. Mar. Biodivers. 2011, 41, 467–479. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Commercial fish and shellfish in the Barents Sea: Have introduced crab species affected the population trajectories of commercial fish? Rev. Fish Biol. Fish. 2015, 25, 297–322. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Red king crab (Paralithodes camtschaticus) fisheries in Russian waters: Historical review and present status. Rev. Fish Biol. Fish. 2018, 28, 331–353. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. New echinoderm-crab epibiotic associations from the coastal Barents Sea. Animals 2021, 11, 917. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Epibiotic communities of common crab species in the coastal Barents Sea: Biodiversity and infestation patterns. Diversity 2022, 14, 6. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Inter-annual dynamics of the Barents Sea red king crab (Paralithodes camtschaticus) stock indices in relation to environmental factors. Polar Sci. 2016, 10, 541–552. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Effects of environmental factors on the abundance, biomass, and individual weight of juvenile red king crabs in the Barents Sea. Front. Mar. Sci. 2020, 7, 726. [Google Scholar] [CrossRef]
- Bakanev, S.V.; Stesko, A.V. Red king crab. In Materials Justifying Changes to the Previously Approved Total Allowable Catch in Fishing Areas in Inland Seas of the Russian Federation, on the Continental Shelf of the Russian Federation, in the Exclusive Economical Zone of the Russian Federation, in the Azov and Caspian Seas in 2021; Sologub, D.O., Ed.; FGBUN VNIRO (Polar branch): Murmansk, Russia, 2021; pp. 2–16. (In Russian) [Google Scholar]
- Dvoretsky, A.G.; Bichkaeva, F.A.; Baranova, N.F.; Dvoretsky, V.G. Fatty acid composition of the Barents Sea red king crab (Paralithodes camtschaticus) leg meat. J. Food Compos. Anal. 2021, 98, 103826. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Cucumaria in Russian waters of the Barents Sea: Biological aspects and aquaculture potential. Front. Mar. Sci. 2021, 8, 613453. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Renewal of the recreational red king crab fishery in Russian waters of the Barents Sea: Potential benefits and costs. Mar. Policy 2022, 136, 104916. [Google Scholar] [CrossRef]
- Kuzmin, S.A.; Gudimova, E.N. Introduction of the Kamchatka (Red King) Crab in the Barents Sea. Peculiarities of Biology, Perspectives of Fishery; Publishing Kola Science Centre Russian Academy of Sciences: Apatity, Russia, 2002. (In Russian) [Google Scholar]
- Sokolov, V.I.; Milyutin, D.M. Distribution, size-sex composition, and reserves of the red king crab (Paralithodes camtschaticus) in the upper sublittoral of the Kola Peninsula (the Barents Sea). Zool. Zhurn. 2006, 85, 158–171. (In Russian) [Google Scholar]
- Pavlova, L.V. Red king crab trophic relations and its influence on bottom biocenoses. In Biology and Physiology of the Red King Crab from the Coastal Zone of the Barents Sea; Matishov, G.G., Ed.; Publishing Kola Scientific Centre Russian Academy of Sciences: Apatity, Russia, 2008; pp. 77–104. (In Russian) [Google Scholar]
- Dvoretsky, A.G.; Dvoretsky, V.G. Distribution of amphipods Ischyrocerus on the red king crab, Paralithodes camtschaticus: Possible interactions with the host in the Barents Sea. Estuar. Coast. Shelf Sci. 2009, 82, 390–396. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Fouling community of the red king crab, Paralithodes camtschaticus (Tilesius 1815), in a subarctic fjord of the Barents Sea. Polar Biol. 2009, 32, 1047–1054. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Some aspects of the biology of the amphipods Ischyrocerus anguipes associated with the red king crab, Paralithodes camtschaticus, in the Barents Sea. Polar Biol. 2009, 32, 463–469. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Population dynamics of the invasive lithodid crab, Paralithodes camtschaticus, in a typical bay of the Barents Sea. ICES J. Mar. Sci. 2013, 70, 1255–1262. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Size-at-age of juvenile red king crab (Paralithodes camtschaticus) in the coastal Barents Sea. Cah. Biol. Mar. 2014, 55, 43–48. [Google Scholar]
- Dvoretsky, A.G.; Dvoretsky, V.G. Hemolymph molting hormone concentrations in red king crabs from the Barents Sea. Polar Biol. 2010, 33, 1293–1298. [Google Scholar] [CrossRef]
- Mota, V.C.; Siikavuopio, S.I.; James, P. Physiological responses to live air transport of red king crab (Paralithodes camtschaticus). Fish. Res. 2021, 237, 105882. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Tipisova, E.V.; Elfimova, A.E.; Alikina, V.A.; Dvoretsky, V.G. Sex hormones in hemolymph of red king crabs from the Barents Sea. Animals 2021, 11, 2149. [Google Scholar] [CrossRef]
- Zenzerov, V.S.; Tipisova, E.V. Initial data on the level of thyroid hormones (Thyroxin, Triiodthyronine) and thyrotropic hormone of the hypophysis in the hemolymph of king crabs from the Barents Sea. Dokl. Biol. Sci. 2009, 428, 462–463. [Google Scholar] [CrossRef]
- Donaldson, W.E.; Byersdorfer, S.E. Biological Field Techniques for Lithodid Crabs. Fairbanks, Alaska Sea Grant College Program; University of Alaska: Fairbanks, AK, USA, 2005. [Google Scholar]
- Dvoretsky, A.G.; Dvoretsky, V.G. Size at maturity of female red king crab, Paralithodes camtschaticus, from the costal zone of Kola Peninsula (southern Barents Sea). Cah. Biol. Mar. 2015, 56, 49–54. [Google Scholar]
- Dvoretsky, A.G.; Dvoretsky, V.G. Does spine removal affect molting process in the king red crab (Paralithodes camtschaticus) in the Barents Sea? Aquaculture 2012, 326–329, 173–177. [Google Scholar] [CrossRef]
- Webster, S.G. Endocrinology of molting. In The Natural History of Crustacea: Physiology; Chang, E.S., Thiel, M., Eds.; Oxford University Press: Oxford, UK, 2015; pp. 1–35. [Google Scholar]
- Uawisetwathana, U.; Leelatanawit, R.; Klanchui, A.; Prommoon, J.; Klinbunga, S.; Karoonuthaisiri, N. Insights into eyestalk ablation mechanism to induce ovarian maturation in the black tiger shrimp. PLoS ONE 2011, 6, e24427. [Google Scholar] [CrossRef] [Green Version]
- Cheng, S.-Y.; Leonard, J.L.; Davis, P.J. Molecular aspects of thyroid hormone actions. Endocrine Rev. 2010, 31, 139–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pillai, S.M.; Verghese, P.U.; Ravichandran, P.; Roy, A.K. Effect of thyroxine on growth and moulting in Penaeus monodon Fabricius. Ind. J. Anim. Sci. 1987, 57, 241–245. [Google Scholar]
- Roustaian, P.; Gaik, L.A. Effect of thyroxine immersion on larval survival, growth and postlarvae production of freshwater prawn, Macrobrachium rosenbergii (de Man). Aquac. Res. 2006, 37, 1378–1380. [Google Scholar] [CrossRef]
- Adiputra, Y.T.; Zairin, M., Jr.; Suprayudi, M.A.; Manalu, W.; Brite, M. The effects of thyroxine hormone on gonadal maturation and growth of male spiny lobster (Panulirus homarus). Malays. J. Sci. 2020, 39, 30–40. [Google Scholar] [CrossRef] [Green Version]
- Jayaprakas, V.; Sambhu, C. Growth characteristics of white prawn Penaeus indicus (Decapoda-Crustacea) under dietary administration of protein hormones. Ind. J. Mar. Sci. 1998, 27, 389–395. [Google Scholar]
- Pinandoyo, D.H.; Windarto, S.; Herawati, V.E. The effects of addition tyroxin hormone on growth and the survival rate of giant prawn Macrobrachium rosenbergii. Int. J. Fish. Aquat. Stud. 2020, 8, 84–87. [Google Scholar]
- Dvoretsky, A.G.; Dvoretsky, V.G. Aquaculture of green sea urchin in the Barents Sea: A brief review of Russian studies. Rev. Aquac. 2020, 12, 1280–1290. [Google Scholar] [CrossRef]
- Syafaat, M.N.; Azra, M.N.; Waiho, K.; Fazhan, H.; Abol-Munafi, A.B.; Ishak, S.D.; Syahnon, M.; Ghazali, A.; Ma, H.; Ikhwanuddin, M. A Review of the nursery culture of mud crabs, genus Scylla: Current progress and future directions. Animals 2021, 11, 2034. [Google Scholar] [CrossRef]
- Hachero-Cruzado, I.; Manchado, M. Dietary phospholipids enhance growth performance and modulate cold tolerance in meagre (Argyrosomus regius) juveniles. Animals 2021, 11, 2750. [Google Scholar] [CrossRef]
- Subramoniam, T.; Kirubagaran, R. Endocrine regulation of vitellogenesis in lobsters. J. Mar. Biol. Ass. India 2010, 52, 229–236. [Google Scholar]
- Lezer, Y.; Aflalo, E.D.; Manor, R.; Sharabi, O.; Abilevich, L.K.; Sagi, A. On the safety of RNAi usage in aquaculture: The case of all-male prawn stocks generated through manipulation of the insulin-like androgenic gland hormone. Aquaculture 2015, 435, 157–166. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sex | Maturity | N | Carapace Length, mm | Weight, g | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| X | SE | Min | Max | X | SE | Min | Max | |||
| M | Juv | 7 | 66.8 | 4.3 | 54.6 | 83.1 | 239.7 | 47.7 | 131 | 432 |
| M | Ad | 29 | 130.7 | 4.3 | 85.6 | 162.7 | 2015.4 | 184.4 | 474 | 3805 |
| M | Tot | 36 | 118.3 | 5.6 | 54.6 | 162.7 | 1670.1 | 190.0 | 131 | 3805 |
| F | Juv | 16 | 71.5 | 3.8 | 49.0 | 91.7 | 293.3 | 40.5 | 97 | 532 |
| F | Ad | 11 | 120.1 | 5.1 | 99.8 | 158.0 | 1449.6 | 270.0 | 665 | 3820 |
| F | Tot | 27 | 91.3 | 5.6 | 49.0 | 158.0 | 764.4 | 156.2 | 97 | 3820 |
| M + F | Juv | 23 | 70.1 | 2.9 | 49.0 | 91.7 | 277.0 | 31.6 | 97 | 532 |
| M + F | Ad | 40 | 127.8 | 3.5 | 85.6 | 162.7 | 1859.8 | 156.4 | 474 | 3820 |
| Hormone | Comparisons | Test | df | F (H) | p |
|---|---|---|---|---|---|
| T3 | Juv-M vs. Juv-F | KWT | 1 | 1.969 | 0.161 |
| Juv-M vs. Ad-M | KWT | 1 | 5.284 | 0.022 | |
| Juv-F vs. Ad-F | ANOVA | 1 | 0.957 | 0.337 | |
| Ad-M vs. Ad-F | ANOVA | 1 | 3.393 | 0.073 | |
| Tot-M vs. Tot-F | ANOVA | 1 | 6.970 | 0.011 | |
| Juv-Tot vs. Ad-tot | ANOVA | 1 | 10.950 | 0.002 | |
| July vs. Oct (Juv) | ANOVA | 1 | 4.540 | 0.045 | |
| May vs. June vs. Aug vs. Oct (Ad) | ANOVA | 3 | 9.480 | <0.001 | |
| T4 | Juv-M vs. Juv-F | KWT | 1 | 0.755 | 0.385 |
| Juv-M vs. Ad-M | KWT | 1 | 1.122 | 0.290 | |
| Juv-F vs. Ad-F | ANOVA | 1 | 7.099 | 0.013 | |
| Ad-M vs. Ad-F | ANOVA | 1 | 0.345 | 0.560 | |
| Tot-M vs. Tot-F | KWT | 1 | 0.866 | 0.352 | |
| Juv-Tot vs. Ad-tot | KWT | 1 | 7.750 | 0.005 | |
| July vs. Oct (Juv) | ANOVA | 1 | 0.001 | 0.995 | |
| May vs. June vs. Aug vs. Oct (Ad) | ANOVA | 3 | 2.120 | 0.116 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dvoretsky, A.G.; Tipisova, E.V.; Alikina, V.A.; Elfimova, A.E.; Dvoretsky, V.G. Thyroid Hormones in Hemolymph of Red King Crabs from the Barents Sea. Animals 2022, 12, 379. https://doi.org/10.3390/ani12030379
Dvoretsky AG, Tipisova EV, Alikina VA, Elfimova AE, Dvoretsky VG. Thyroid Hormones in Hemolymph of Red King Crabs from the Barents Sea. Animals. 2022; 12(3):379. https://doi.org/10.3390/ani12030379
Chicago/Turabian StyleDvoretsky, Alexander G., Elena V. Tipisova, Viktoria A. Alikina, Aleksandra E. Elfimova, and Vladimir G. Dvoretsky. 2022. "Thyroid Hormones in Hemolymph of Red King Crabs from the Barents Sea" Animals 12, no. 3: 379. https://doi.org/10.3390/ani12030379