
Assessment of Biostimulation Methods Based on Chemical Communication in Female Doe Reproduction

 ,
,  , and
, and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Sample Collection
2.3. Semen Processing and Artificial Insemination
2.4. Reproductive Managemenent
2.5. Experimental Design
2.6. Statistical Analysis
3. Results
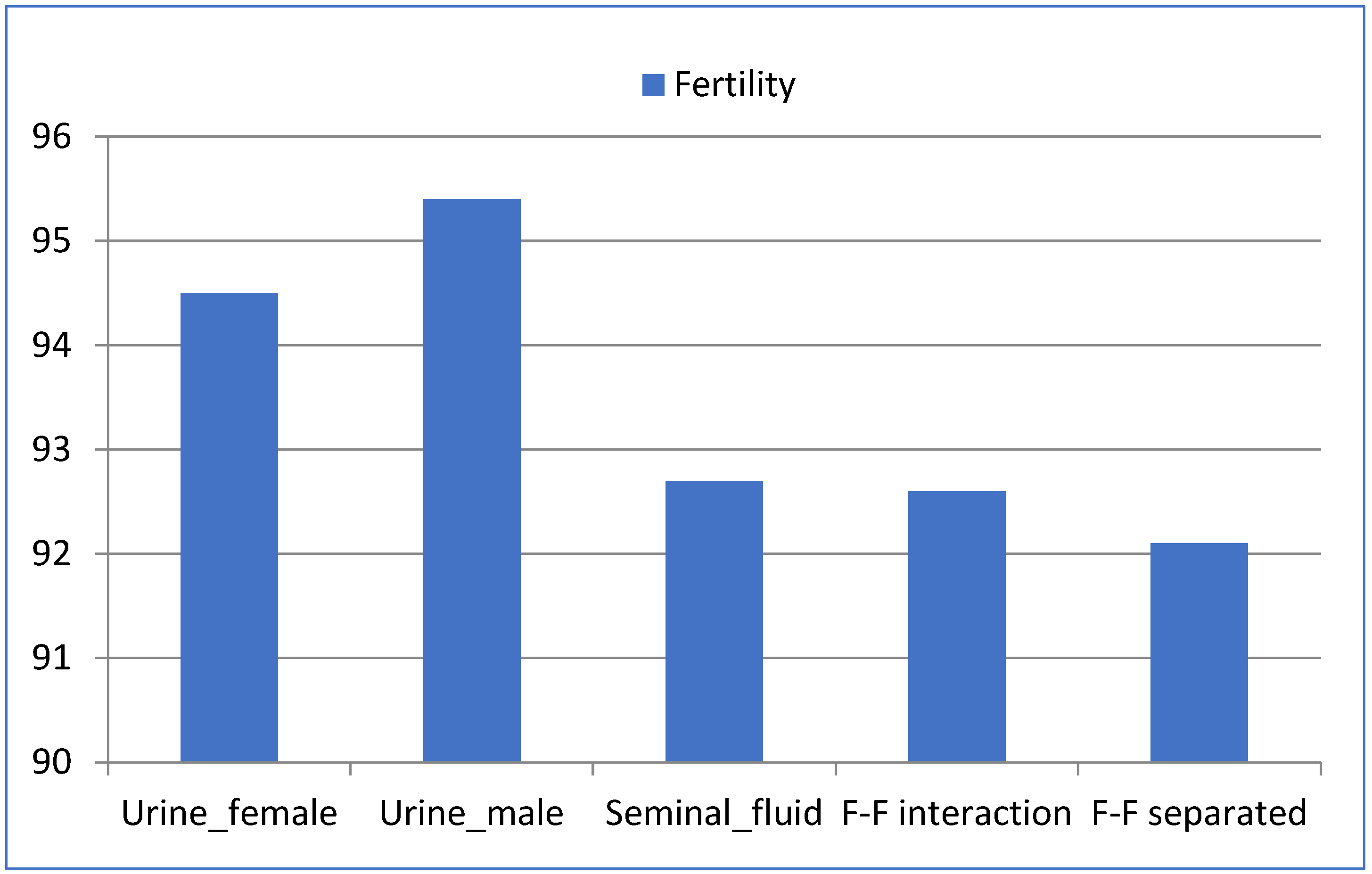
3.1. Fertility
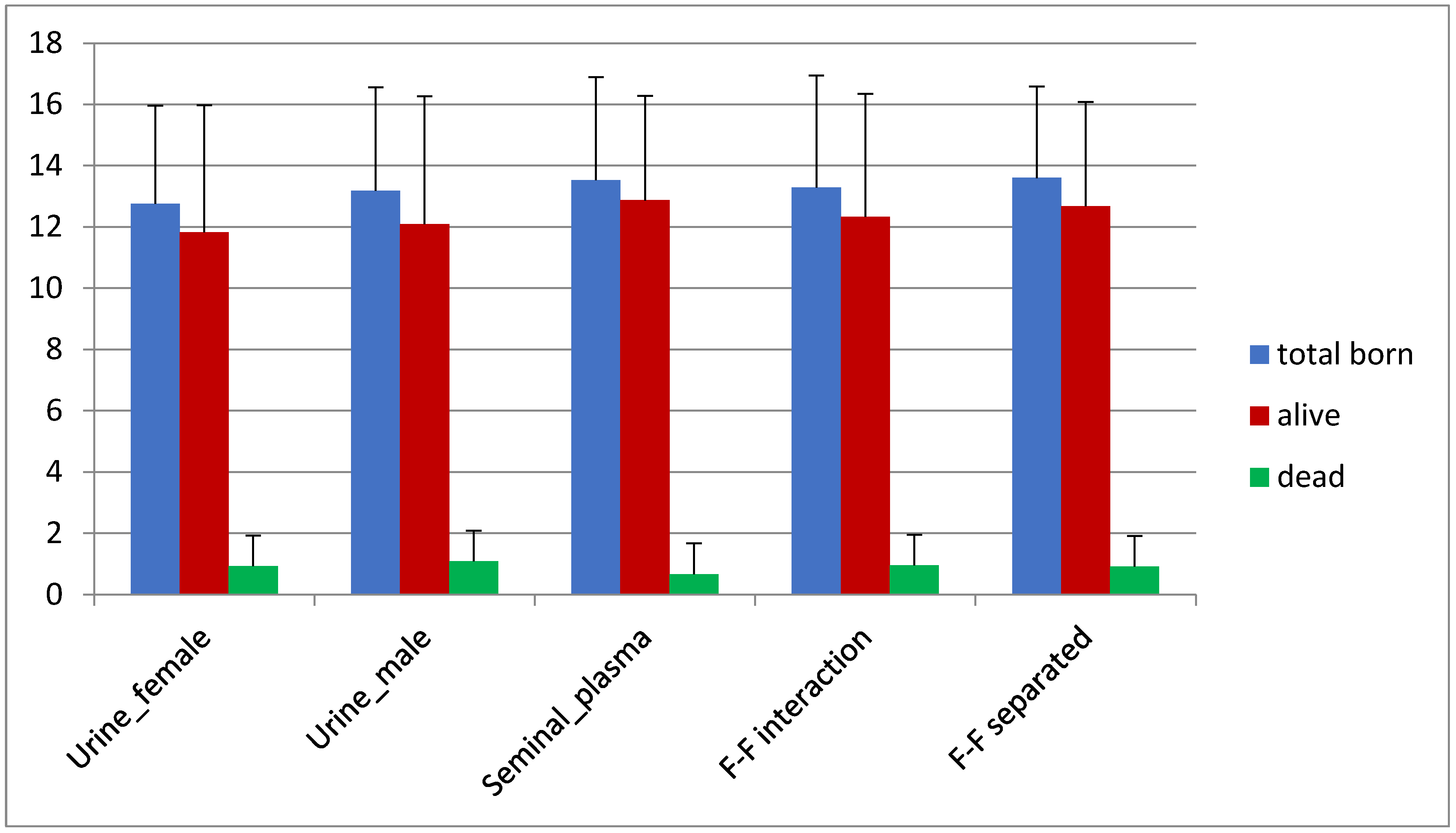
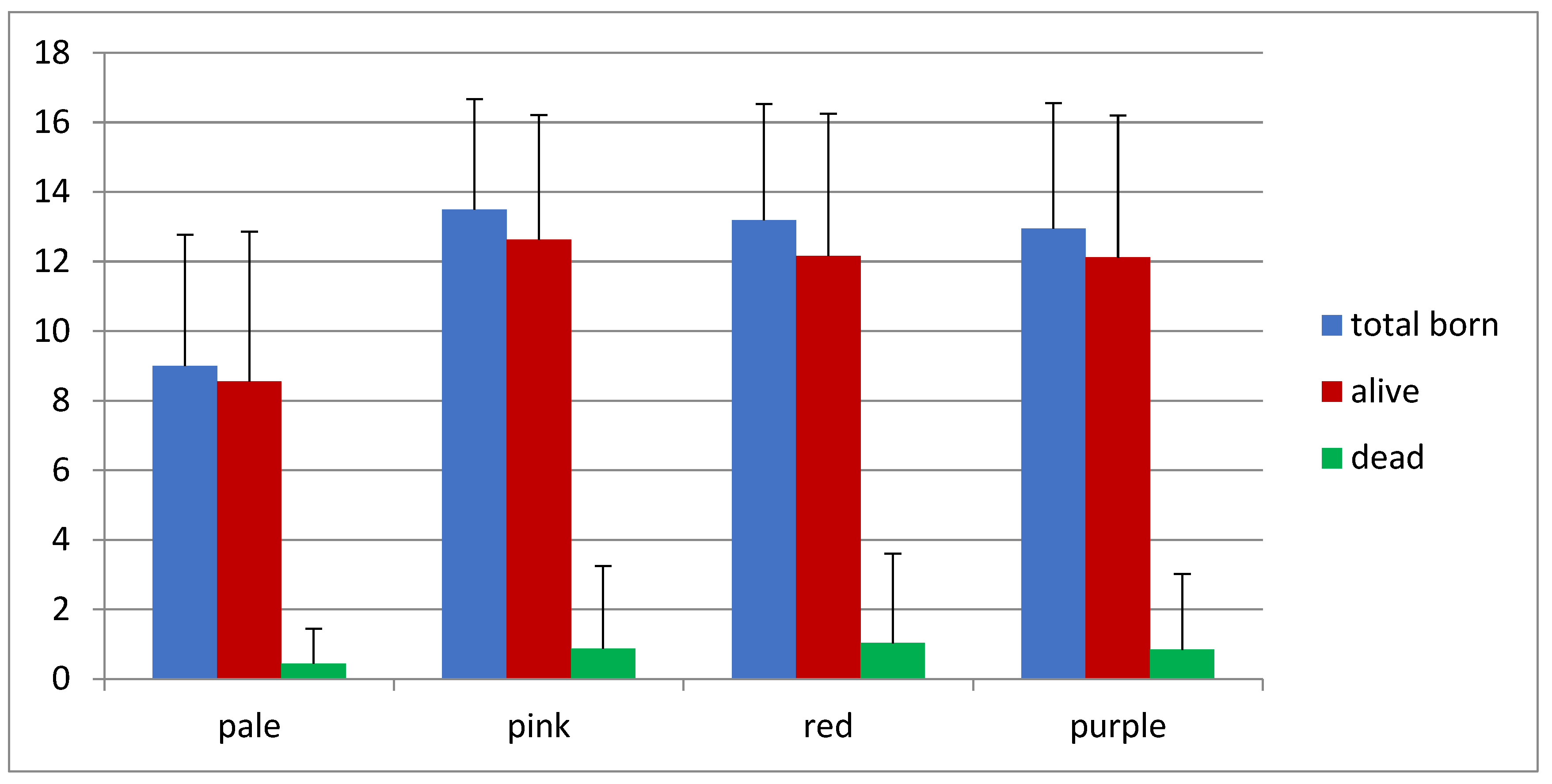
3.2. Prolificacy
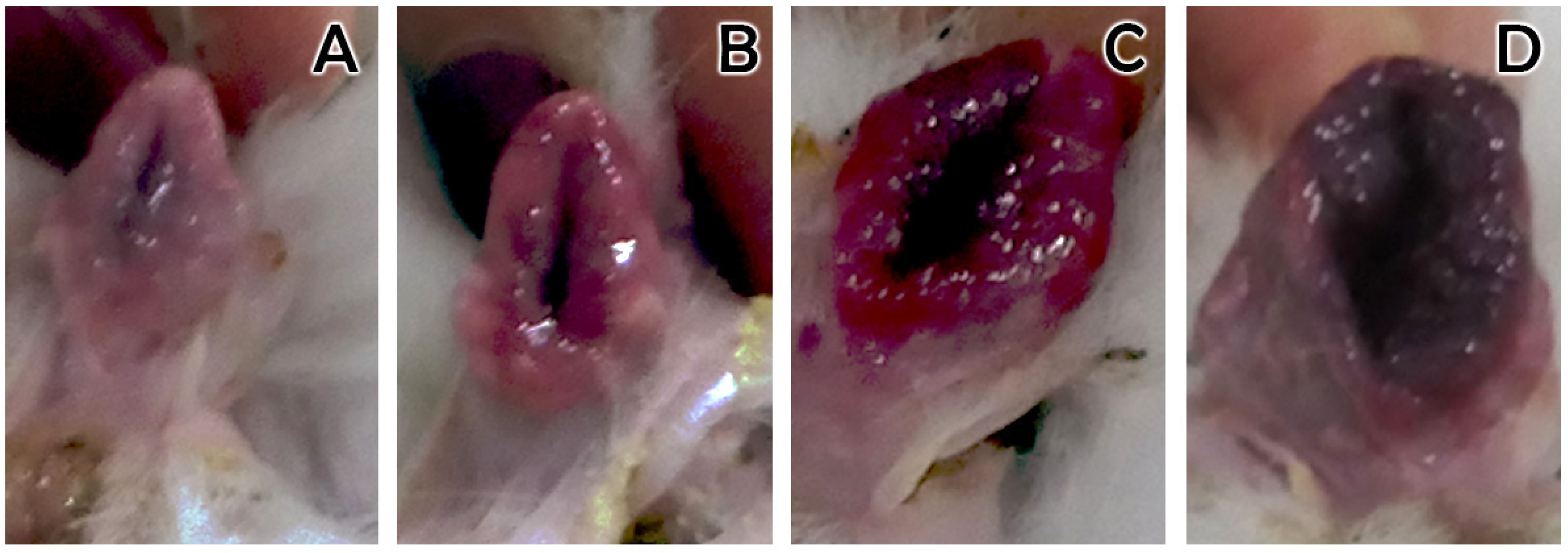
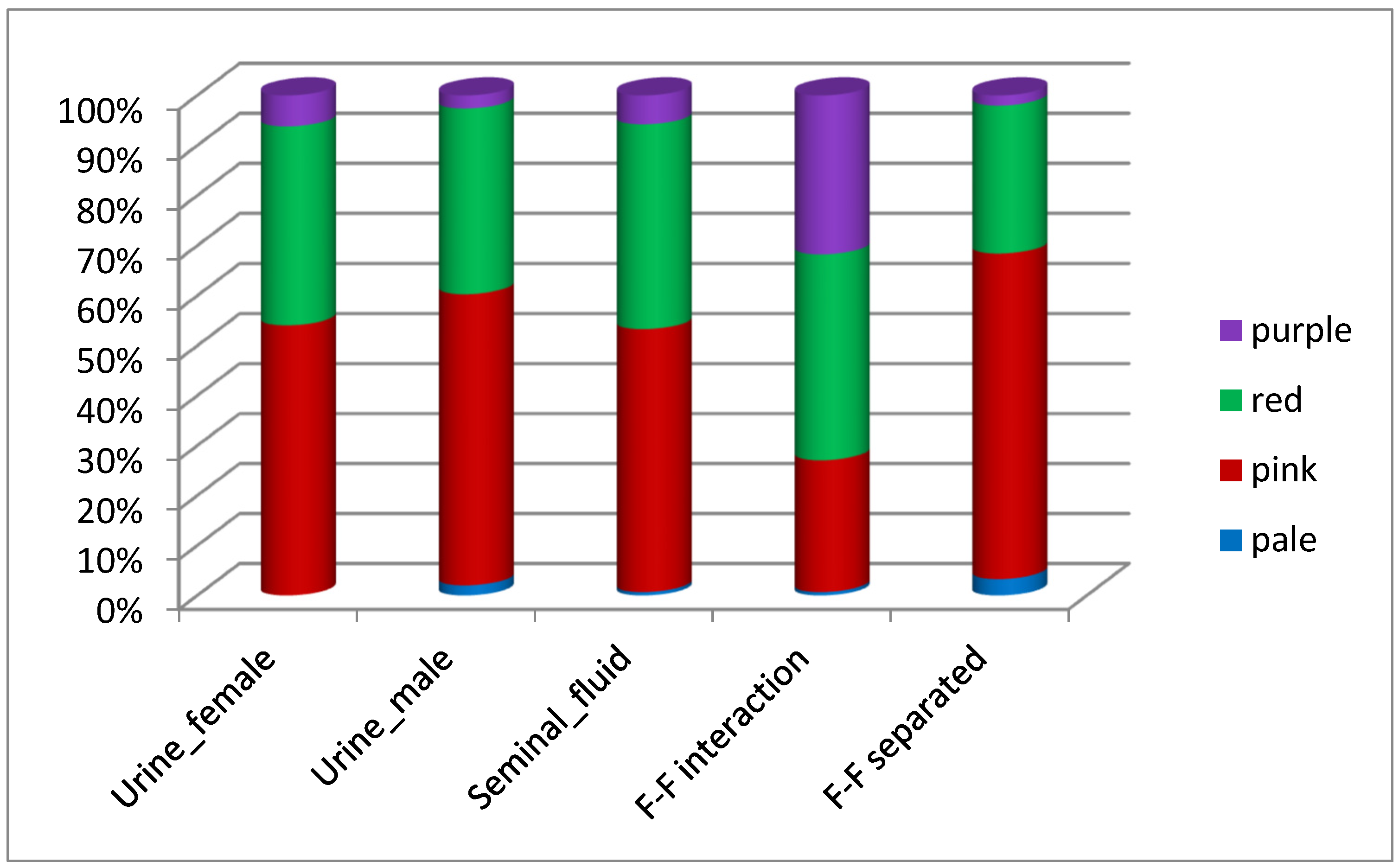
3.3. Receptivity
4. Discussion
4.1. Social Interaction Seems Not Influencing Reproductive Physiology in Farm Female Doe
4.2. Urine as a Potential Source of Sex Pheromones in Female Doe Reproduction
4.3. Seminal Plasma Might Arise as a New Source of Pheromones
4.4. Practical Considerations When Assessing Biostimulation Methods
4.5. Could Biostimulation Methods Reduce or Replace Hormonal Treatments?
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wei, D.; Talwar, V.; Lin, D. Neural circuits of social behaviours: Innate yet flexible. Neuron 2021, 109, 1600–1620. [Google Scholar] [CrossRef] [PubMed]
- Kwon, J.T.; Ryu, C.; Lee, H.; Sheffield, A.; Fan, J.; Cho, D.H.; Bigler, S.; Sullivan, H.A.; Choe, H.K.; Wickersham, I.R.; et al. An amygdala circuit that suppresses social engagement. Nature 2021, 593, 114–118. [Google Scholar] [CrossRef]
- Wyatt, T.D. Pheromones and Animal Behaviour: Chemical Signals and Signatures, 2nd ed.; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- Theau-Clément, M.; Castellini, C.; Maertens, L.; Boiti, C. Biostimulations applied to rabbit reproduction: Theory and practice. World Rabbit Sci. 1998, 6, 179–184. [Google Scholar] [CrossRef] [Green Version]
- Choudhary, S.; Lal-Kamboj, M. Effect of bull biostimulation on the oestrous behaviour of pubertal Sahiwal (Bos indicus) heifers. Anim. Reprod. Sci. 2019, 209, 106149. [Google Scholar] [CrossRef] [PubMed]
- Rekwot, P.I.; Ogwub, D.; Oyedipea, E.O.; Sekonia, V.O. The role of pheromones and biostimulation in animal reproduction. Anim. Reprod. Sci. 2001, 65, 157–170. [Google Scholar] [CrossRef]
- Novotny, M.V. Pheromones, binding proteins, and receptor responses in rodents. Biochem. Soc. Trans. 2003, 31, 117–122. [Google Scholar] [CrossRef]
- Boehm, U.; Zou, Z.; Buck, L.B. Feedback Loops Link Odor and Pheromone Signaling with Reproduction. Cell 2005, 123, 683–695. [Google Scholar] [CrossRef] [Green Version]
- Melo, A.I.; González-Mariscal, G. Communication by Olfactory Signals in Rabbits: Its Role in Reproduction. Vit. Horm. 2010, 83, 351–371. [Google Scholar] [CrossRef]
- Kerketta, S.; Singh, A.; Sahu, S.; Upadhyay, D.; Abdullah, M.; Yogi, R.; Singh, M. Biostimulation: Innovative Tool to Improve Reproductive Efficiency in Farm Animals. Int. Livest. Res. 2017, 7, 18–30. [Google Scholar] [CrossRef]
- Karlson, P.; Lüscher, M. Pheromones: A new term for a class of biologically active substances. Nature 1959, 183, 55–56. [Google Scholar] [CrossRef]
- Wyatt, T.D. Pheromones and signature mixtures: Defining species-wide signals and variable cues for identity in both invertebrates and vertebrates. J. Comp. Physiol. A Neuroethol. Sens. Neural Behav. Physiol. 2010, 196, 685–700. [Google Scholar] [CrossRef] [PubMed]
- Roberts, S.A.; Simpson, D.M.; Armstrong, S.D.; Davidson, A.J.; Robertson, D.H.; McLean, L.; Beynon, R.J.; Hurst, J.L. Darcin: A male pheromone that stimulates female memory and sexual attraction to an individual male’s odour. BMC Biol. 2010, 8, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mastrogiacomo, R.; Ambrosio, C.D.; Niccolini, A.; Serra, A.; Gazzano, A.; Scaloni, A.; Pelosi, P. An Odorant-Binding Protein Is Abundantly Expressed in the Nose and in the Seminal Fluid of the Rabbit. PLoS ONE 2014, 9, e111932. [Google Scholar]
- Scott, A.M.; Zhang, Z.; Jia, L.; Li, K.; Zhang, Q.; Dexheimer, T.; Ellsworth, E.; Ren, J.; Chung-Davidson, Y.W.; Zu, Y.; et al. Spermine in semen of male sea lamprey acts as a sex pheromone. PLoS Biol. 2019, 17, e3000332. [Google Scholar] [CrossRef]
- Demir, E.; Li, K.; Bobrowski-Khoury, N.; Sanders, J.I.; Beynon, R.J.; Hurst, J.L.; Kepecs, A.; Axel, R. The pheromone darcin drives a circuit for innate and reinforced behaviours. Nature 2020, 578, 137–159. [Google Scholar] [CrossRef]
- Luo, M.; Fee, M.S.; Katz, L.C. Encoding pheromonal signals in the accessory olfactory bulb of behaving mice. Science 2003, 299, 1196–1201. [Google Scholar] [CrossRef] [Green Version]
- Kimoto, H.; Haga, S.; Sato, K.; Touhara, K. Sex-specific peptides from exocrine glands stimulate mouse vomeronasal sensory neurons. Nature 2005, 437, 898–901. [Google Scholar] [CrossRef]
- Gelez, H.; Fabre-Nys, C. The male effect in sheep and goats: A review of the respective roles of the two olfactory systems. Horm. Behav. 2004, 46, 257–271. [Google Scholar] [CrossRef] [PubMed]
- El-Azzazi, F.E.; Hegab, I.M.; Hanafy, A.M. Biostimulation and reproductive performance of artificially inseminated rabbit does (oryctolagus cuniculus). World Rabbit Sci. 2017, 25, 313–321. [Google Scholar] [CrossRef] [Green Version]
- Bonanno, A.; Mazza, F.; Alabiso, M.; Di Grigoli, A.; Alicata, M.L. Effects of bio-stimulation induced by the presence of buck on reproductive performance of rabbit does. Ital. J. Anim. Sci. 2003, 2, 133–135. [Google Scholar]
- Rosa, H.J.D.; Bryant, M.J. The ‘ram effect’ as a way of modifying the reproductive activity in the ewe. Small Rumin. Res. 2002, 45, 1–16. [Google Scholar] [CrossRef]
- González-Urdiales, R. Bioestimulación en la coneja. ¿Alternativa a los tratamientos hormonales? Cunicultura 2005, 30, 7–17. [Google Scholar]
- Mousa-Balabel, T.M. Using light and melatonin in the management of New Zealand White rabbits. Open Vet. J. 2011, 1, 1–6. [Google Scholar] [PubMed]
- Quintela, L.; Peña, A.; Barrio, M.; Vega, M.D.; Díaz, R.; Maseda, F.; García, P. Reproductive performance of multiparous rabbit lactating does: Effect of lighting programs and PMSG use. Reprod. Nutr. Dev. 2001, 41, 247–257. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Dalmán, C.; Gonzalez-Mariscal, G. Major role of suckling stimulation for inhibition of estrous behaviours in lactating rabbits: Acute and chronic effects. Horm. Behav. 2012, 61, 108–113. [Google Scholar] [CrossRef]
- Keeling, L.; Tunón, H.; Olmos-Antillón, G.; Berg, C.; Jones, M.; Stuardo, L.; Swanson, J.; Wallenbeck, A.; Winckler, C.; Blokhuis, H. Animal Welfare and the United Nations Sustainable Development Goals. Front. Vet. Sci. 2019, 6, 336. [Google Scholar] [CrossRef]
- Quintela, L.A.; Peña, A.I.; Vega, M.D.; Gullón, J.; Prieto, M.C.; Barrio, M.; Becerra, J.J.; Maseda, F.; Herradón, P.G. Ovulation induction in rabbit does submitted to artificial insemination by adding buserelin to the seminal dose. Reprod. Nutr. Dev. 2004, 44, 79–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quintela, L.A.; Peña, A.I.; Vega, M.D.; Gullón, J.; Prieto, M.C.; Barrio, M.; Becerra, J.J.; Herradón, P.G. Reproductive Performance of Rabbit Does Artificially Inseminated via Intravaginal Administration of des-Gly 10, D-Ala6-LHRH Ethylamide as Ovulation Inductor. Reprod. Dom. Anim. 2009, 44, 829–833. [Google Scholar] [CrossRef] [PubMed]
- Vega, M.D.; Barrio, M.; Quintela, L.A.; Becerra, J.J.; Cainzos, J.; Prieto, A.; Rodríguez-Zamora, A.; Herradón, P.G. Evolución del manejo reproductivo en cunicultura. Inf. Técnica Económica Agrar. 2012, 108, 172–190. [Google Scholar]
- Cervantes, M.P.; Palomino, J.M.; Adams, G.P. In vivo imaging in the rabbit as a model for the study of ovulation-inducing factors. Lab. Anim. 2015, 49, 1–9. [Google Scholar] [CrossRef]
- Hassanein, E.M.; Hashem, N.M.; El-Azrak, K.E.D.M.; Gonzalez-Bulnes, A.; Hassan, G.A.; Salem, M.H. Efficiency of GnRH–Loaded Chitosan Nanoparticles for Inducing LH Secretion and Fertile Ovulations in Protocols for Artificial Insemination in Rabbit Does. Animals 2021, 11, 440. [Google Scholar] [CrossRef]
- Theau-Clément, M.; Lebas, F.; Boiti, C.; Brecchia, G.; Mercier, P. Influence of different eCG doses on sexual receptivity and productivity of rabbit does. World Rabbit Sci. 2008, 16, 65–72. [Google Scholar]
- Lebas, F.; Theau-Clement, M.; Remy, B.; Drion, P.J.; Beckers, F. Production of anti-PMSG antibodies and its relation to the roductivity of rabbit does. World Rabbit Sci. 1996, 4, 57–62. [Google Scholar]
- Stradaioli, G.; Verini-Sulpizi, A.; Monaci, M.; Canali, C.; Boiti, C. Effect of different doses of PMSG on ovarian response and in vitro embryo development in rabbits. World Rabbit Sci. 1997, 5, 143–148. [Google Scholar] [CrossRef]
- Castellini, C.; Dal-Bosco, A.; Arias-Álvarez, M.; Lorenzo, P.L.; Cardinali, R.; Garcia-Rebollar, P. The main factors affecting the reproductive performance of rabbit does: A review. Anim. Reprod. Sci. 2010, 122, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Rebollar, P.G.; Milanés, A.; Pereda, N.; Millán, P.; Cano, P.; Esquifino, A.I.; Villarroel, M.; Silván, G.; Lorenzo, P.L. Oestrus synchronisation of rabbit does at early post-partum by doe—Litter separation or ECG injection: Reproductive parameters and endocrine profiles. Anim. Reprod. Sci. 2006, 93, 218–230. [Google Scholar] [CrossRef] [PubMed]
- Arias-Alvarez, M.; García-García, R.M.; Torres-Rovira, L.; González-Bulnes, A.; Rebollar, P.-G.; Lorenzo, P.L. Influence of hormonal and nonhormonal estrus synchronization methods on follicular and oocyte quality in primiparous lactating does at early postpartum period. Theriogenology 2010, 73, 26–35. [Google Scholar] [CrossRef]
- Villamayor, P.R.; Cifuentes, J.M.; Fdz-de-Troconiz, P.; Sanchez-Quinteiro, P. Morphological and immunohistochemical study of the rabbit vomeronasal organ. J. Anat. 2018, 233, 814–827. [Google Scholar] [CrossRef]
- Chen, P.; Hong, W. Neural Circuit Mechanisms of Social Behaviour. Neuron 2018, 98, 16–30. [Google Scholar] [CrossRef] [Green Version]
- Walkden-Brown, S.W.; Martin, G.B.; Restall, B.J. Role of male-female interaction in regulating reproduction in sheep and goats. J. Reprod. Fertil. Suppl. 1999, 54, 243–257. [Google Scholar] [CrossRef]
- Martin, G.B.; Milton, J.T.B.; Davidson, R.H.; Banchero-Hunzicker, G.E.; Lindsay, D.R.; Blache, D. Natural methods for increasing reproductive efficiency in small ruminants. Anim. Reprod. Sci. 2004, 82, 231–246. [Google Scholar] [CrossRef]
- Chenoweth, P.J.; Landaeta-Hernández, A.J.; Flöercke, C. Reproductive and Maternal Behaviour of Livestock. Genet. Behav. Domest. Anim. 2014, 5, 158–194. [Google Scholar]
- Zalesky, D.D.; Day, M.L.; Garcia-Winder, M.; Imakawa, K.; Kittok, R.J.; D’Occhio, M.J.; Kinder, J.E. Influence of exposure to bulls on resumption of estrous cycles following parturition in beef cows. J. Ser. Nebraska Agric. Exp. Sta. Anim. Sci. 1984, 59, 1134–1139. [Google Scholar]
- Kustos, K.; Eiben, C.S.; Szendrő, Z.S.; Theau-Clement, M.; Gódor, S.-N.É.; Jovanczai, Z.S. Effect on reproductive traits of male presence among rabbit does before artificial insemination (Preliminary Results). World Rabbit Sci. 2000, 8, 161–166. [Google Scholar]
- Ola, S.I.; Oyegbade, M.O. Buck effect on rabbit oestrous: Vulva colour, vaginal lumen cells and ovarian follicle populations. World Rabbit Sci. 2012, 20, 71–79. [Google Scholar] [CrossRef]
- Coombes, H.A.; Stockley, P.; Hurst, J.L. Female Chemical Signalling Underlying Reproduction in Mammals. J. Chem. Ecol. 2018, 44, 851–873. [Google Scholar] [CrossRef] [Green Version]
- Restall, B.J.; Restall, H.; Walkden-Brown, S.W. The induction of ovulation in anovulatory goats by oestrous females. Anim. Reprod. Sci. 1995, 40, 299–303. [Google Scholar] [CrossRef]
- Delcroix, I.; Mauget, R.; Signoret, J.P. Existence of synchronization of reproduction at the level of the social group of the European wild boar (Sus scrofa). J. Reprod. Fertil. 1990, 89, 613–617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McClintock, M.K. Menstrual synchronism and suppression. Nature 1971, 229, 244–245. [Google Scholar] [CrossRef] [PubMed]
- Wright, I.A.; Rhind, S.-M.; Smith, A.J.; Whyte, T.K. Female-female influences on the duration of the post-partum anoestrous period in beef cows. Anim. Sci. 2010, 59, 49–53. [Google Scholar] [CrossRef]
- Luzynski, K.C.; Nicolakis, D.; Marconi, M.A.; Zala, S.M.; Kwak, J.; Penn, D.J. Pheromones that correlate with reproductive success in competitive conditions. Sci. Rep. 2021, 11, 21970. [Google Scholar] [CrossRef] [PubMed]
- Whitten, W.K. Modifications of the oestrus cycle of the mouse by external stimuli associated with the male: Changes in the oestrus cycle determined by vaginal smears. J. Endocrin. 1958, 11, 307. [Google Scholar] [CrossRef] [PubMed]
- DeCatanzaro, D. Sex steroids as pheromones in mammals: The exceptional role of estradiol. Horm. Behav. 2015, 68, 103–116. [Google Scholar] [CrossRef]
- Izard, M.K.; Vandenbergh, J.G. The effects of bull urine on puberty and calving date in crossbred beef heifers. J. Anim. Sci. 1982, 55, 1160–1168. [Google Scholar] [CrossRef] [Green Version]
- Knight, T.W.; Lynch, P.R. Source of ram pheromones that stimulate ovulation in the ewe. Anim. Reprod. Sci. 1980, 3, 133–136. [Google Scholar] [CrossRef]
- Mak, G.K.; Enwere, E.K.; Gregg, C.; Pakarainen, T.; Poutanen, M.; Huhtaniemi, I.; Weiss, S. Male pheromone–stimulated neurogenesis in the adult female brain: Possible role in mating behaviour. Nat. Neurosci. 2007, 10, 1003–1011. [Google Scholar] [CrossRef] [PubMed]
- Ratto, M.H.; Leduc, Y.A.; Valderrama, X.P.; van-Straaten, K.E.; Delbaere, L.T.; Pierson, R.A.; Adams, G.P. The nerve of ovulation-inducing factor in semen. Proc. Natl. Acad. Sci. USA 2012, 109, 15042–15047. [Google Scholar] [CrossRef] [Green Version]
- Adams, G.P.; Ratto, M.H.; Silva, M.E.; Carrasco, R.A. Ovulation-inducing factor (OIF/ß-NGF) in seminal plasma: A review and update. Reprod. Dom. Anim. 2016, 51, 4–17. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.E.; Smulders, J.P.; Guerra, M.; Valderrama, X.P.; Letelier, C.; Adams, G.P.; Ratto, M.H. Cetrorelix suppresses the preovulatory LH surge and ovulation induced by ovulation-inducing factor (OIF) present in llama seminal plasma. Reprod. Biol. Endocr. 2011, 9, 74. [Google Scholar] [CrossRef] [Green Version]
- El-Allali, K.; El-Bousmaki, N.; Ainani, H.; Simonneaux, V. Effect of the Camelid’s Seminal Plasma Ovulation-Inducing Factor/β-NGF: A Kisspeptin Target Hypothesis. Front. Vet. Sci. 2017, 4, 99. [Google Scholar] [CrossRef]
- Sanchez-Rodriguez, A.; Abad, P.; Arias-Alvarez, M.; Rebollar, P.G.; Bautista, J.M.; Lorenzo, P.L.; García-García, R.M. Recombinant rabbit beta nerve growth factor production and its biological effects on sperm and ovulation in rabbits. PLoS ONE 2019, 14, e0223091. [Google Scholar]
- Sanchez-Rodriguez, A.; Arias-Álvarez, M.; Millán, P.; Lorenzo, P.L.; García-García, R.M.; Rebollar, P.G. Physiological effects on rabbit sperm and reproductive response to recombinant rabbit beta nerve growth factor administered by intravaginal route in rabbit does. Theriogenology 2020, 157, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Hellier, V.; Brock, O.; Candlish, M.; Desroziers, E.; Aoki, M.; Mayer, C.; Piet, R.; Herbison, R.; Colledge, W.H.; Prévot, V.; et al. Female sexual behaviour in mice is controlled by kisspeptin neurons. Nat. Commun. 2018, 9, 400. [Google Scholar] [CrossRef] [PubMed]
- Schjenken, J.E.; Robertson, S.A. The Female Response to Seminal Fluid. Physiol. Rev. 2020, 100, 1077–1117. [Google Scholar] [CrossRef]
- Fernández, M.D.V. Optimization of the Use of Artificial Insemination in Rabbit Farming. Ph.D. Thesis, University of Santiago de Compostela, Santiago de Compostela, Spain, 2010. [Google Scholar]
- Theau-Clément, M. Preparation of the rabbit doe to insemination: A review. World Rabbit Sci. 2007, 15, 61–80. [Google Scholar] [CrossRef] [Green Version]
- Szendrő, Z.S.; Szendrő, K.; Dalle-Zotte, A. Management of Reproduction on Small, Medium and Large Rabbit Farms: A Review. Asian-Aust. J. Anim. Sci. 2012, 25, 738–748. [Google Scholar] [CrossRef] [Green Version]
- Yang, R.; Raper, K.C.; Lusk, J.L. The Impact of Hormone Use Perception on Consumer Meat Preference. In Proceedings of the Southern Agricultural Economics Association (SAEA) Annual Meeting, Mobile, AL, USA, 4–7 February 2017. [Google Scholar]
- EMA/CVMP/156095/2017. European Public MRL Assessment Report (EPMAR) for Alarelin (All Food Producing Species). Available online: https://www.ema.europa.eu/en/documents/mrl-report/alarelin-all-food-producing-species-european-public-maximum-residue-limit-assessment-report-epmar_en.pdf (accessed on 15 December 2021).
- 2016/2077(INI). Report on Minimum Standards for the Protection of Farm Rabbits. Available online: https://www.europarl.europa.eu/doceo/document/A-8-2017-0011_EN.pdf (accessed on 15 December 2021).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Definition | Measurement |
|---|---|---|
| Receptivity | Behavioral receptivity to mating, which indicates estrus | Vulvar color (pale, pink, red, purple) |
| Fertility | Average number of females that give birth successfully | Percentage of females that give birth (kindling rate) |
| Prolificacy | Average number of born kits/litter | Number of born kits/litter |
| Number of born alive kits/litter | Average number of born alive kits/litter | Number of born alive kits/litter |
| Number of born dead kits/litter | Average number of born dead kits/litter | Number of born dead kits/litter |
| Experimental Group | N (Fertility) | Fertility% | N (Prolificacy) | Prolificacy ± SD | Alive ± SD | Dead ± SD |
|---|---|---|---|---|---|---|
| Urine_female | 138/146 | 94.5 | 138 | 12.76 ± 3.2 | 11.83 ± 4.15 | 0.93 ± 2.38 |
| Urine_male | 144/151 | 95.4 | 144 | 13.18 ± 3.38 | 12.09 ± 4.18 | 1.09 ± 2.79 |
| Seminal_plasma | 127/137 | 92.7 | 127 | 13.53 ± 3.36 | 12.87 ± 3.41 | 0.67 ± 1.99 |
| F–F interaction | 137/148 | 92.6 | 137 | 13.29 ± 3.65 | 12.33 ± 4.02 | 0.96 ± 2.35 |
| F–F separated | 140/152 | 92.1 | 140 | 13.6 ± 2.99 | 12.68 ± 3.4 | 0.92 ± 2.49 |
| Insemination Number | ||||||
| 1 | 234/259 | 90.3 a | 234 | 13.46 ± 3.73 abc | 12.35 ± 4.2 | 1.11 ± 2.39 |
| 2 | 224/244 | 91.8 a | 224 | 13.55 ± 2.97 ab | 12.72 ± 3.44 | 0.83 ± 2.15 |
| 3 | 228/231 | 98.7 b | 228 | 12.78 ± 3.19 ac | 11.98 ± 3.88 | 0.81 ± 2.7 |
| Vulvar Color | ||||||
| Pale | 09/10 | 90 | 9 | 9 ± 3.77 a | 8.56 ± 4.3 ac | 0.44 ± 1.01 |
| Pink | 352/377 | 93.4 | 352 | 13.49 ± 3.18 b | 12.63 ± 3.58 b | 0.87 ± 2.38 |
| Red | 256/276 | 92.8 | 256 | 13.19 ± 3.34 b | 12.16 ± 4.09 b | 1.03 ± 2.58 |
| Purple | 69/71 | 97.2 | 69 | 12.95 ± 3.61 b | 12.12 ± 4.08 c | 0.84 ± 2.18 |
| Kindling Number | ||||||
| 3 | 52/56 | 92.9 | 52 | 14.13 ± 4.17 | 13.15 ± 4.32 | 0.98 ± 1.84 |
| 4 | 144/157 | 91.7 | 144 | 13.34 ± 3.27 | 12.58 ± 3.88 | 0.76 ± 2.26 |
| 5 | 126/138 | 91.3 | 126 | 13.53 ± 3.13 | 12.94 ± 3.46 | 0.6 ± 1.8 |
| 6 | 120/130 | 92.3 | 120 | 13.4 ± 3.48 | 11.89 ± 4.42 | 1.51 ± 3.29 |
| 7 | 120/128 | 93.8 | 120 | 12.7 ± 3.4 | 11.77 ± 3.78 | 0.93 ± 2.27 |
| 8 | 85/86 | 98.8 | 85 | 13.03 ± 2.69 | 12 ± 3.47 | 1.04 ± 2.86 |
| 9 | 39/39 | 100 | 39 | 12.82 ± 3.33 | 12.44 ± 3.36 | 0.38 ± 1.13 |
| Variable | Values | OR (Ods Ratio) | Confidential Interval for OR | p-Value |
|---|---|---|---|---|
| Insemination | first | reference | ||
| second | 1.2 | 0.646–2.215 | 0.568 | |
| third | 8.12 | 2.418–27.266 | 0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Villamayor, P.R.; Gullón, J.; Yáñez, U.; Sánchez, M.; Sánchez-Quinteiro, P.; Martínez, P.; Quintela, L. Assessment of Biostimulation Methods Based on Chemical Communication in Female Doe Reproduction. Animals 2022, 12, 308. https://doi.org/10.3390/ani12030308
Villamayor PR, Gullón J, Yáñez U, Sánchez M, Sánchez-Quinteiro P, Martínez P, Quintela L. Assessment of Biostimulation Methods Based on Chemical Communication in Female Doe Reproduction. Animals. 2022; 12(3):308. https://doi.org/10.3390/ani12030308
Chicago/Turabian StyleVillamayor, Paula R., Julián Gullón, Uxía Yáñez, María Sánchez, Pablo Sánchez-Quinteiro, Paulino Martínez, and Luis Quintela. 2022. "Assessment of Biostimulation Methods Based on Chemical Communication in Female Doe Reproduction" Animals 12, no. 3: 308. https://doi.org/10.3390/ani12030308