
Neuroanatomical and Immunohistological Study of the Main and Accessory Olfactory Bulbs of the Meerkat (Suricata suricatta)




Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Extraction
2.2. Sample Processing for Histological Study
2.3. Immunohistochemical Labelling (IHQ)
2.4. Histochemical Labelling (HQ) with Lectins
2.4.1. LEA and BSI-B4 HQ Protocol
2.4.2. UEA-I HQ Protocol
2.5. Acquisition of Images and Digital Treatment
3. Results
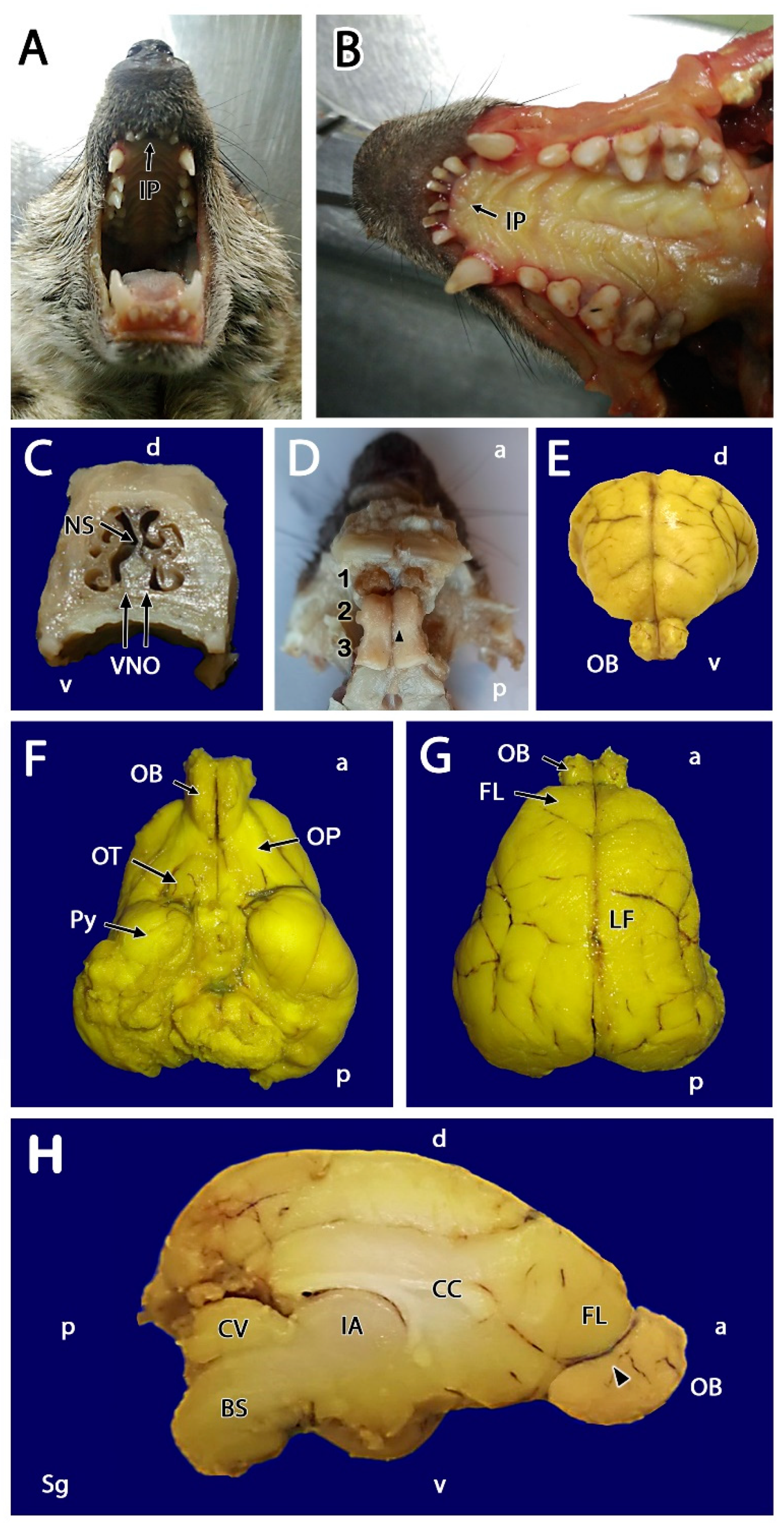
3.1. Macroscopic Study
3.2. Microscopical Study
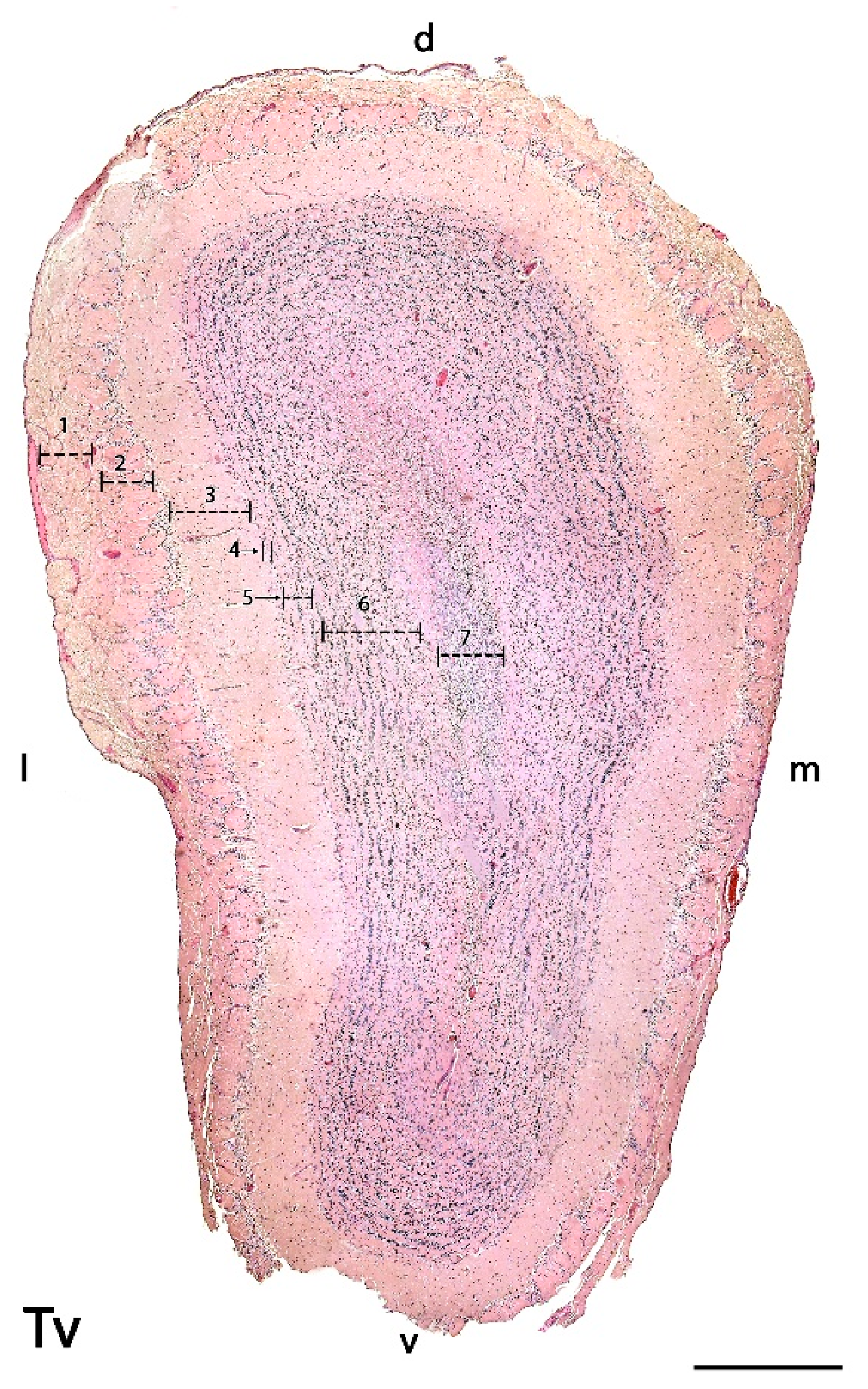
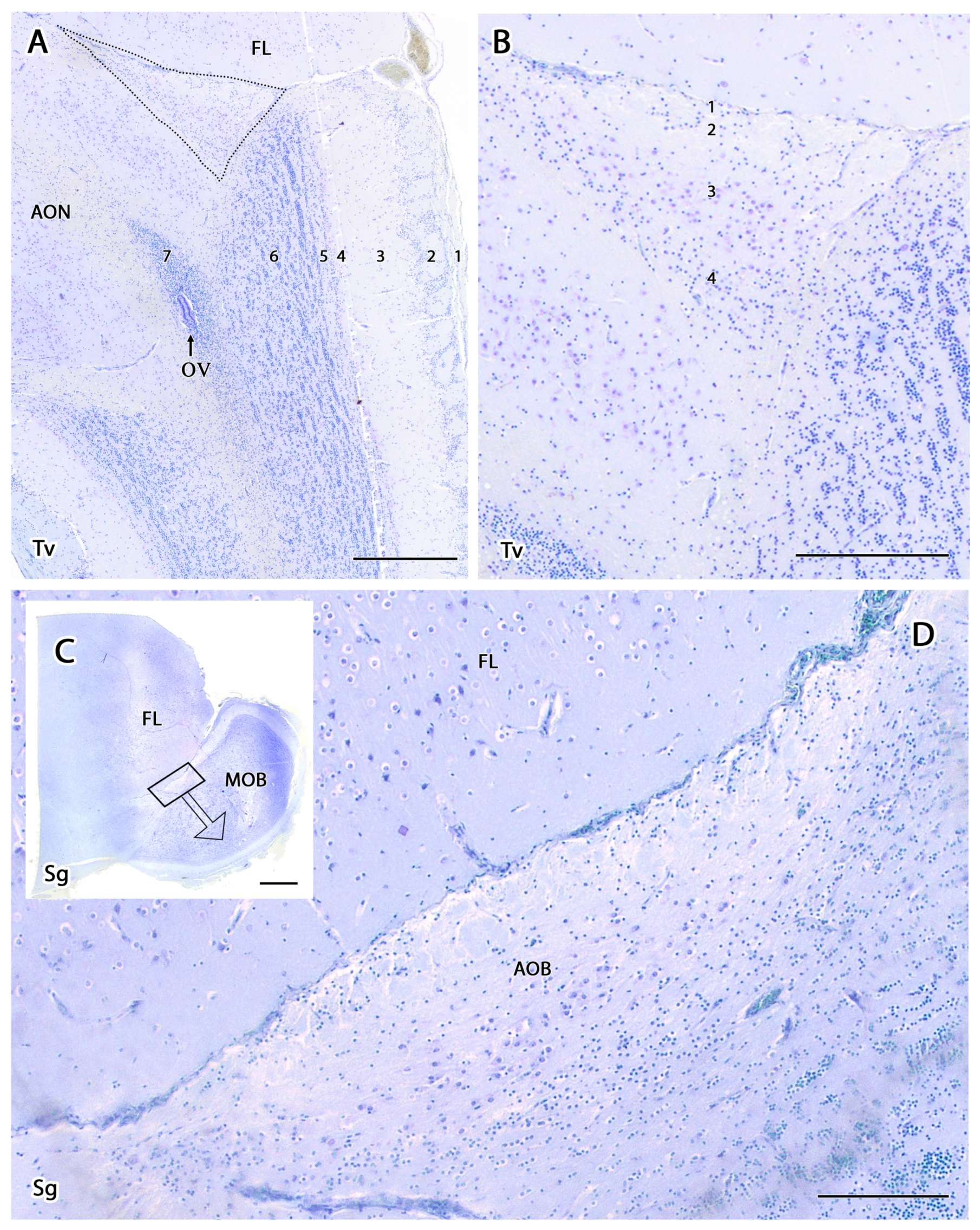
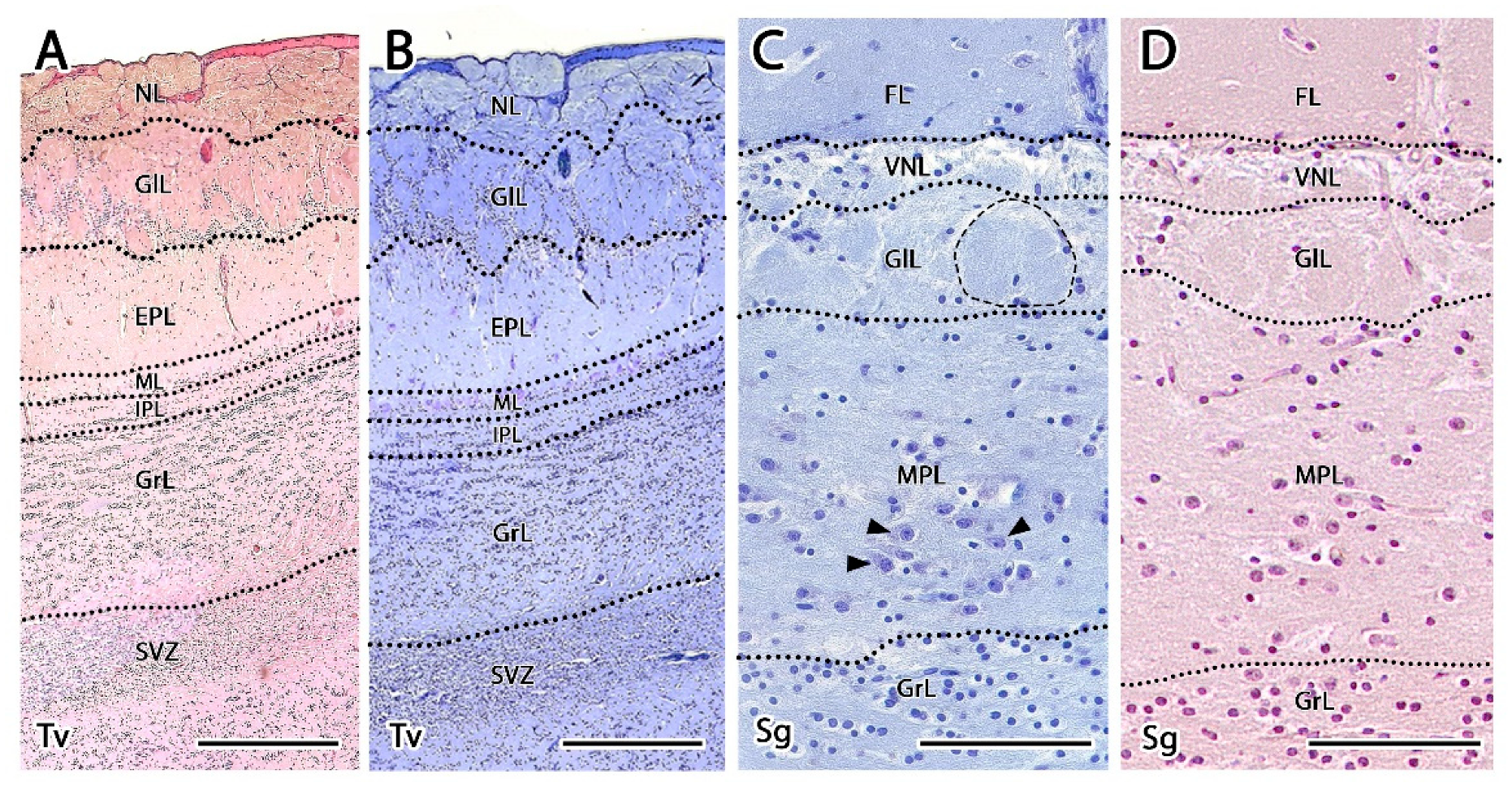
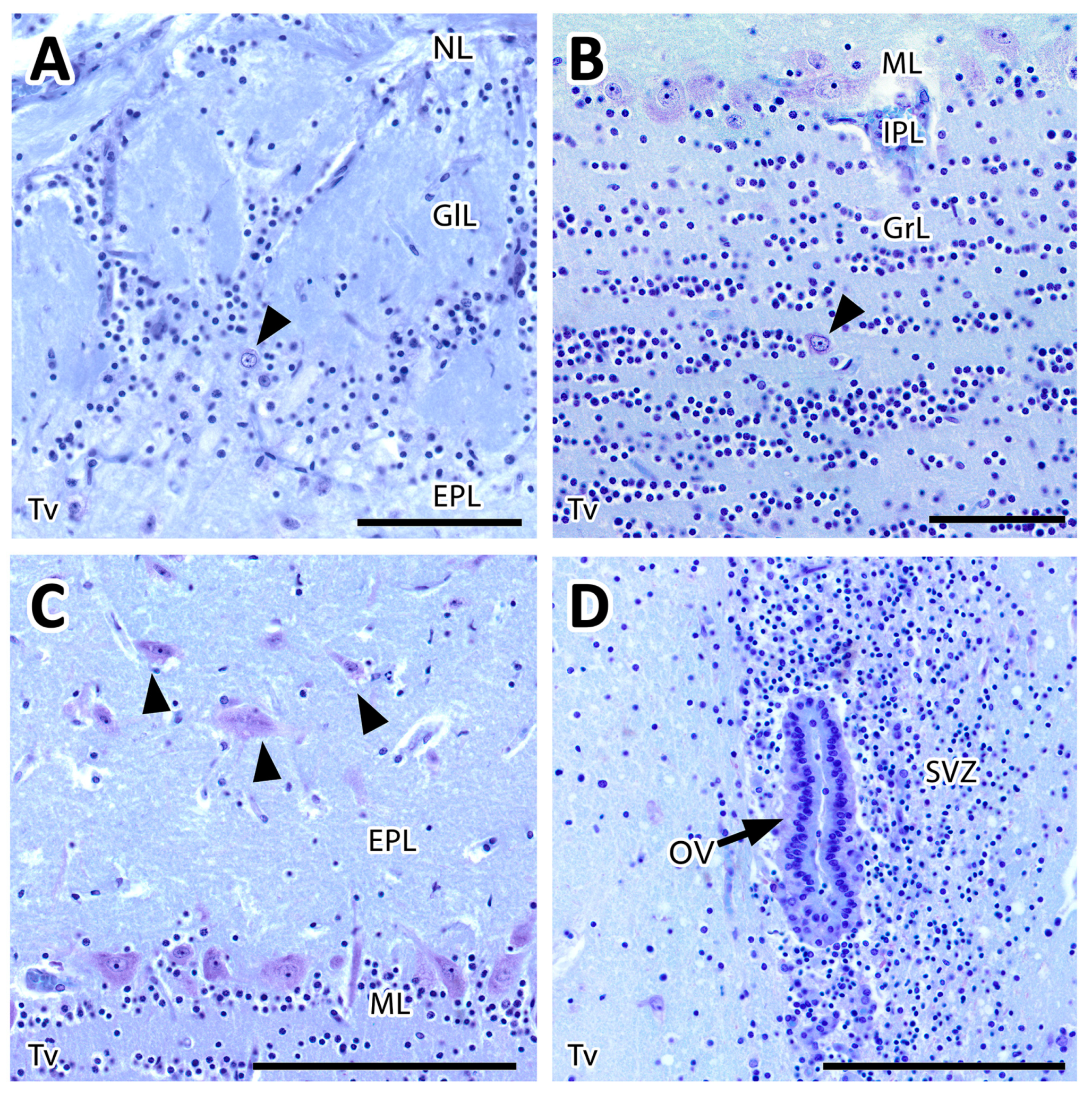
3.2.1. Histological Study
- Olfactory nerve layer (ONL): Formed by the axons of the olfactory nerves that reach the MOB.
- Glomerular layer (GlL): Composed of the glomeruli, which are spherical structures delimited by periglomerular (PG) cells that originate from the confluence of olfactory nerve axons synapsing with the dendrites of mitral cells, which represent the second neurons in olfactory pathways.
- External plexiform layer (EPL): A nerve plexus with a low cell density, primarily occupied by the dendrites of mitral cells.
- Mitral cell layer (MCL): Formed by the somas of mitral cells.
- Internal plexiform layer (IPL): A thin band of white matter interposed between the mitral and granular layers that is very reduced in the meerkat.
- Granular layer (GrL): Contains large clusters of granule cells, which serve as inhibitory neurons in the neural circuit of the MOB.
- Subventricular zone (SVZ): Nervous tissue arranged around the rostral horn of the encephalic lateral ventricles, primarily composed of neuronal precursor cells involved in adult neurogenesis. The SVZ in meerkats is highly and strikingly developed, presenting a large cellular infiltrate.
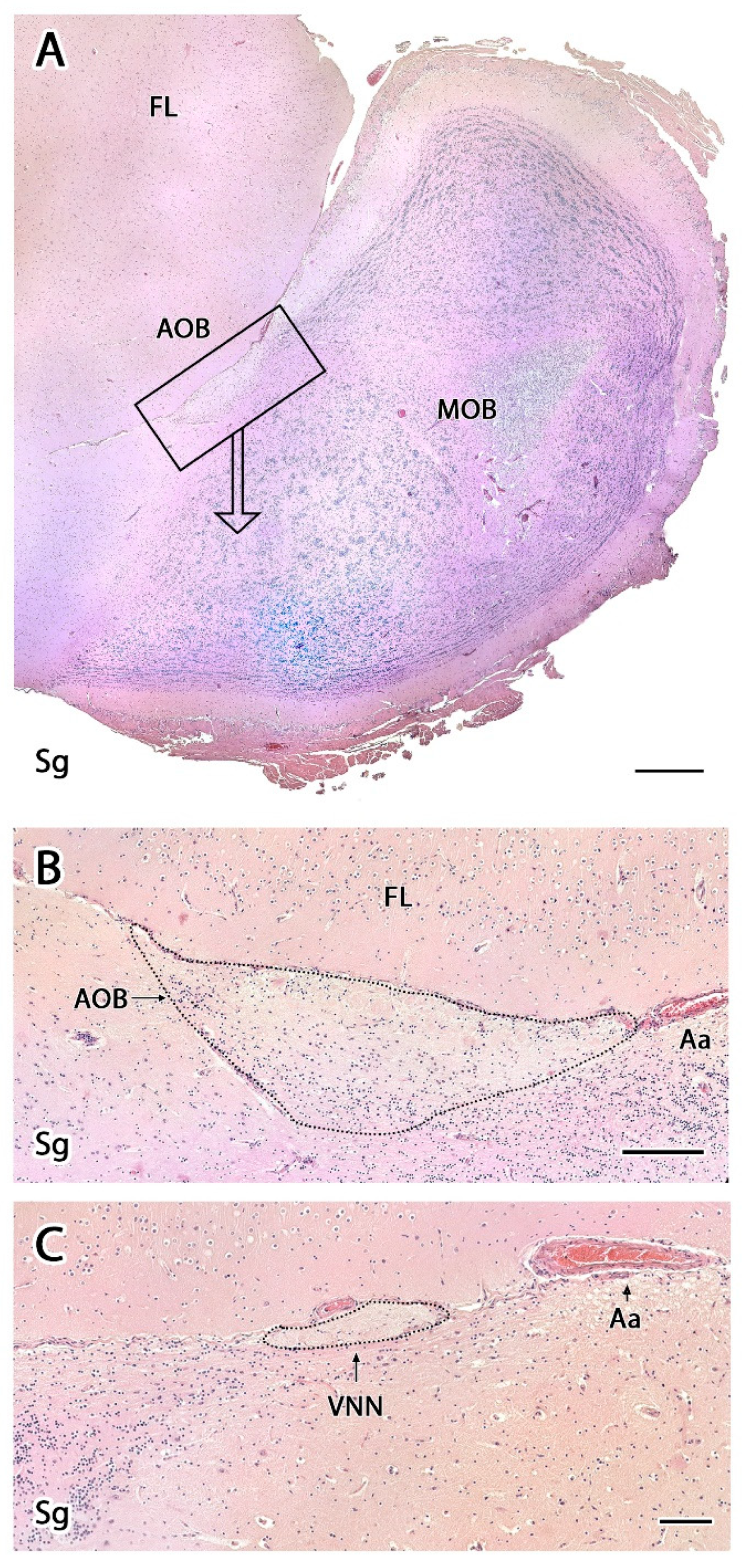
- Vomeronasal nerve layer (VNL): formed by the arrival of VNN axons.
- Glomerular layer (GlL): glomeruli in the AOB show poorer definition and fewer PG cells than their MOB equivalents (Figure 5C,D).
- Mitral-plexiform layer (MPL): the cells of the mitral layer are not organized in a monolayer, as observed in the MOB (Figure 4A,B), but are diffusely distributed, occupying a large neuropil due to the fusion of the external and internal plexiform layers with the mitral layer, forming what we term the MPL. The main cells found in this layer present an ovoidal morphology, reminiscent of the morphology of mitral cells in the MOB (Figure 5C,D).
- Granular layer (GrL): presenting with a higher cell density, the neurons in this layer have small nuclei and compact cytoplasms (Figure 5C,D).
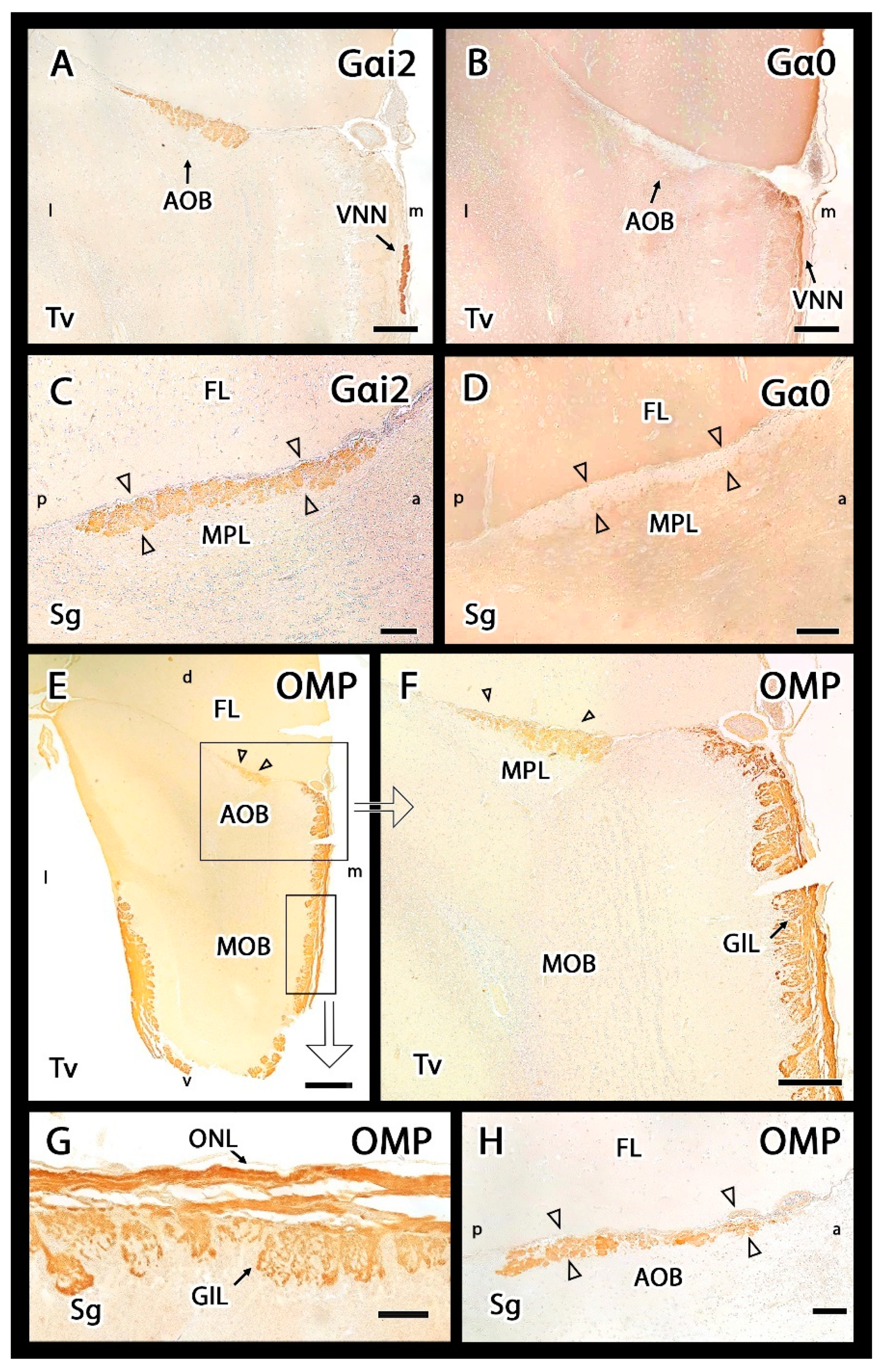
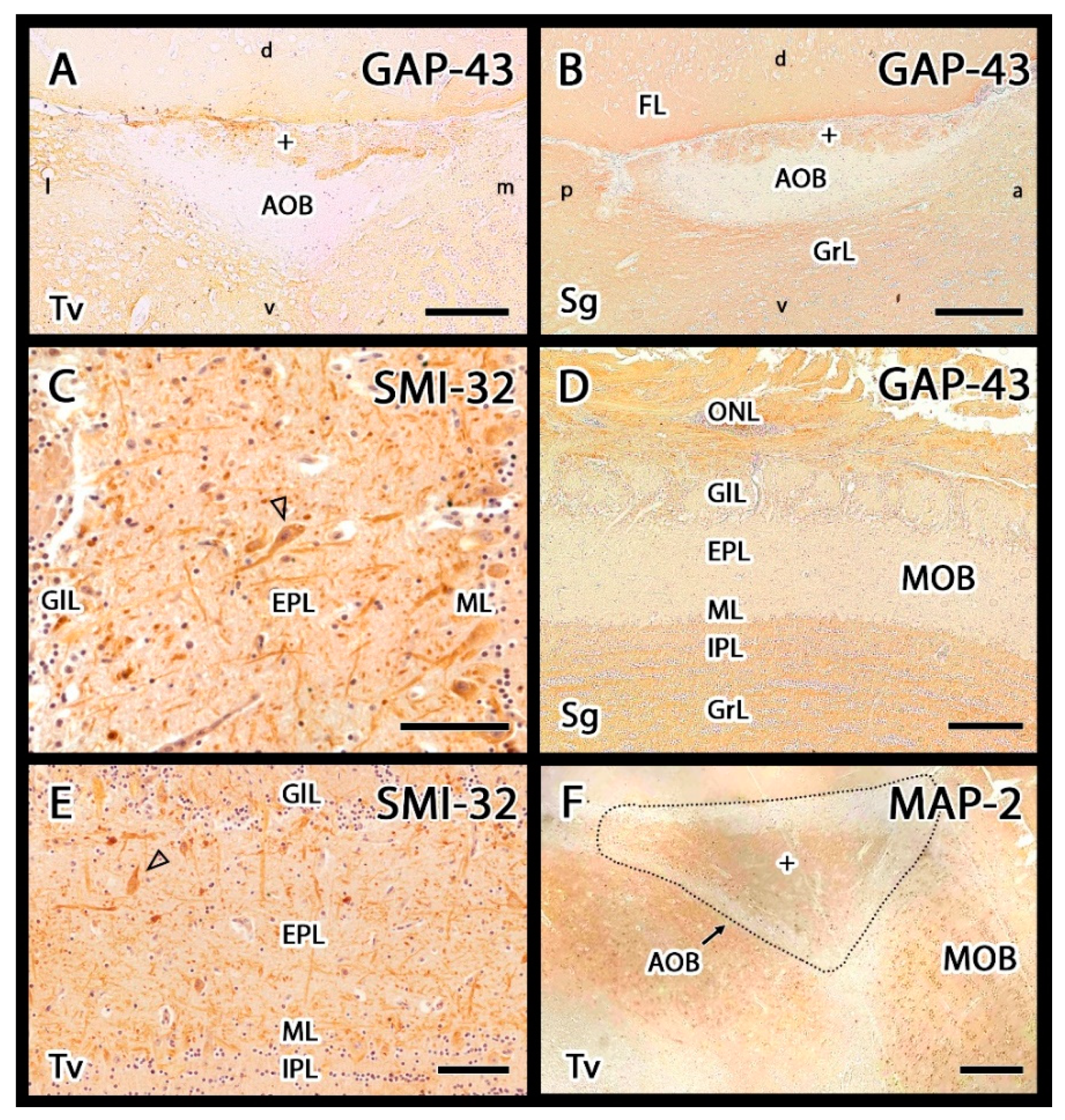
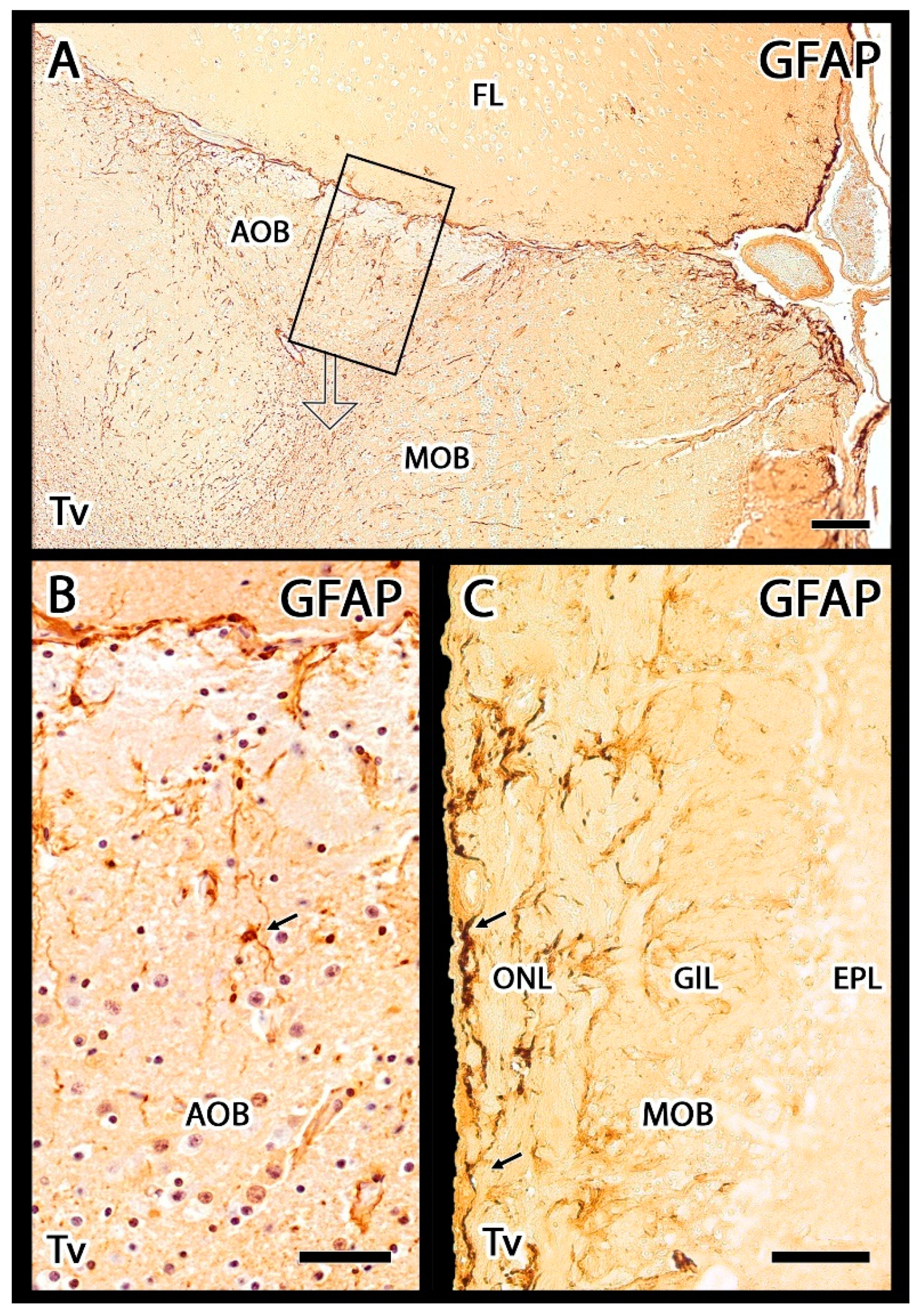
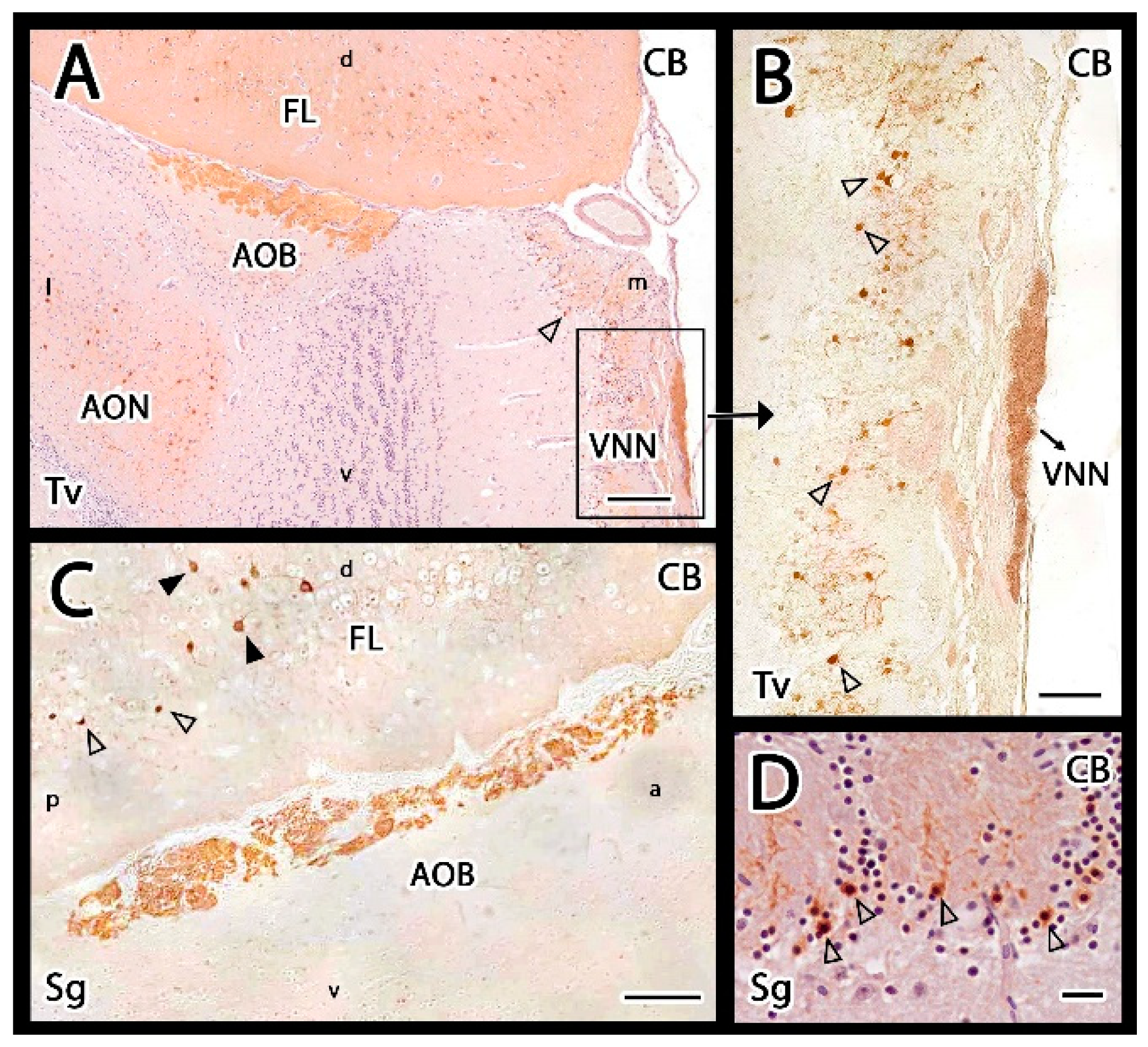
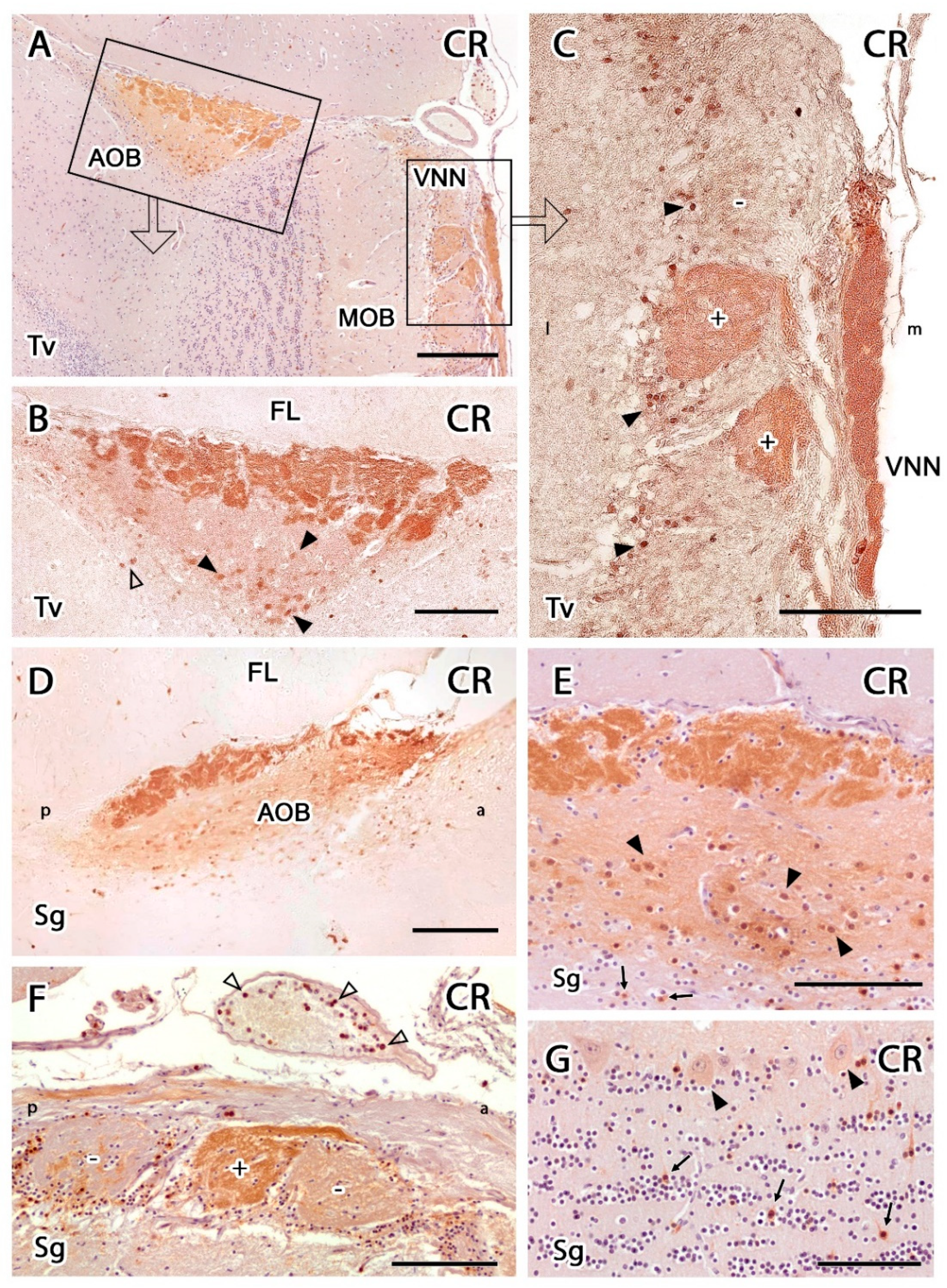
3.2.2. Immunohistochemical Study
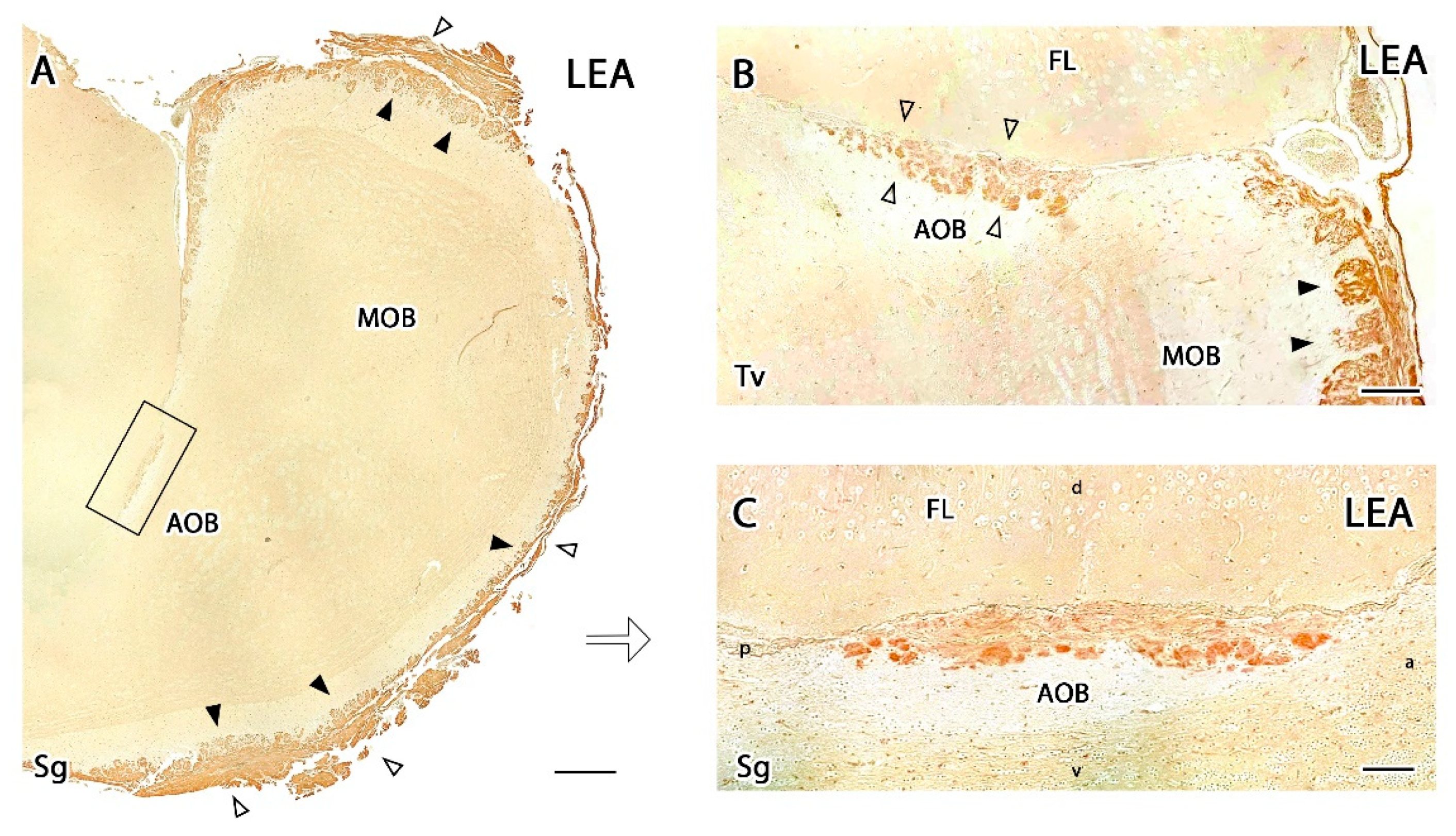
3.2.3. Lectin Histochemical Study
4. Discussion
4.1. Neuroanatomical Study
4.2. Inmunohistochemical Study
4.3. Lectin Histochemical Study
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Meister, M. On the dimensionality of odor space. Elife 2015, 4, e07865. [Google Scholar] [CrossRef] [PubMed]
- Kokocińska-Kusiak, A.; Woszczyło, M.; Zybala, M.; Maciocha, J.; Barłowska, K.; Dzięcioł, M. Canine Olfaction: Physiology, Behavior, and Possibilities for Practical Applications. Animals 2021, 11, 2463. [Google Scholar] [CrossRef] [PubMed]
- Mollo, E.; Garson, M.J.; Polese, G.; Amodeo, P.; Ghiselin, M.T. Taste and smell in aquatic and terrestrial environments. Nat. Prod. Rep. 2017, 34, 496–513. [Google Scholar] [CrossRef] [PubMed]
- Villamayor, P.R.; Arana, Á.J.; Coppel, C.; Ortiz-Leal, I.; Torres, M.V.; Sánchez-Quinteiro, P.; Sánchez, L. A comprehensive structural, lectin and immunohistochemical characterization of the zebrafish olfactory system. Sci. Rep. 2021, 11, 8865. [Google Scholar] [CrossRef] [PubMed]
- Saraiva, L.R.; Ahuja, G.; Ivandic, I.; Syed, A.S.; Marioni, J.C.; Korsching, S.I.; Logan, D.W. Molecular and neuronal homology between the olfactory systems of zebrafish and mouse. Sci. Rep. 2015, 5, 11487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breer, H.; Fleischer, J.; Strotmann, J. The sense of smell: Multiple olfactory subsystems. Cell. Mol. Life Sci. 2006, 63, 1465–1475. [Google Scholar] [CrossRef]
- Salazar, I.; Sánchez-Quinteiro, P.; Barrios, A.W.; López Amado, M.; Vega, J.A. Anatomy of the olfactory mucosa. Handb. Clin. Neurol. 2019, 164, 47–65. [Google Scholar] [CrossRef] [PubMed]
- Munger, S.D. Olfaction: Noses within noses. Nature 2009, 459, 521–522. [Google Scholar] [CrossRef]
- Firestein, S. How the olfactory system makes sense of scents. Nature 2001, 413, 211–218. [Google Scholar] [CrossRef]
- Zou, D.J.; Feinstein, P.; Rivers, A.L.; Mathews, G.A.; Kim, A.; Greer, C.A.; Mombaerts, P.; Firestein, S. Postnatal refinement of peripheral olfactory projections. Science 2004, 304, 1976–1979. [Google Scholar] [CrossRef]
- Kondoh, D.; Tanaka, Y.; Kawai, Y.K.; Mineshige, T.; Watanabe, K.; Kobayashi, Y. Morphological and histological features of the vomeronasal organ in african pygmy hedgehog (Atelerix albiventris). Animals 2021, 11, 1462. [Google Scholar] [CrossRef] [PubMed]
- Ennis, M.; Holy, T.E. Anatomy and neurobiology of the main and accessory olfactory bulbs. In Handbook of Olfaction and Gustation; Doty, R.L., Ed.; Wiley: Hoboken, NJ, USA, 2015; pp. 157–182. [Google Scholar] [CrossRef]
- Fortes-Marco, L.; Lanuza, E.; Martinez-Garcia, F. Of pheromones and kairomones: What receptors mediate innate emotional responses? Anat. Rec. 2013, 296, 1346–1363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ortiz-Leal, I.; Torres, M.V.; Villamayor, P.R.; López-Beceiro, A.; Sanchez-Quinteiro, P. The vomeronasal organ of wild canids: The fox (Vulpes vulpes) as a model. J. Anat. 2020, 237, 890–906. [Google Scholar] [CrossRef] [PubMed]
- Booth, K.K.; Katz, L.S. Role of the vomeronasal organ in neonatal offspring recognition in sheep. Biol. Reprod. 2000, 63, 953–958. [Google Scholar] [CrossRef] [Green Version]
- Pallé, A.; Montero, M.; Fernández, S.; Tezanos, P.; de Las Heras, J.A.; Luskey, V.; Birnbaumer, L.; Zufall, F.; Chamero, P.; Trejo, J.L. Gαi2(+) vomeronasal neurons govern the initial outcome of an acute social competition. Sci. Rep. 2020, 10, 894. [Google Scholar] [CrossRef]
- Villafranca-Faus, M.; Vila-Martín, M.E.; Esteve, D.; Merino, E.; Teruel-Sanchís, A.; Cervera-Ferri, A.; Martínez-Ricós, J.; Lloret, A.; Lanuza, E.; Teruel-Martí, V. Integrating pheromonal and spatial information in the amygdalo-hippocampal network. Nat. Commun. 2021, 12, 5286. [Google Scholar] [CrossRef] [PubMed]
- Del Punta, K.; Leinders-Zufall, T.; Rodriguez, I.; Jukam, D.; Wysocki, C.J.; Ogawa, S.; Zufall, F.; Mombaerts, P. Deficient pheromone responses in mice lacking a cluster of vomeronasal receptor genes. Nature 2002, 419, 70–74. [Google Scholar] [CrossRef]
- Dulac, C.; Wagner, S. Genetic analysis of brain circuits underlying pheromone signaling. Annu. Rev. Genet. 2006, 40, 449–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brennan, P.A. The vomeronasal system. Cell. Mol. Life Sci. 2001, 58, 546–555. [Google Scholar] [CrossRef]
- Brennan, P.A.; Zufall, F. Pheromonal communication in vertebrates. Nature 2006, 444, 308–315. [Google Scholar] [CrossRef]
- Zufall, F.; Leinders-Zufall, T. Mammalian pheromone sensing. Curr. Opin. Neurobiol. 2007, 17, 483–489. [Google Scholar] [CrossRef]
- Young, J.M.; Massa, H.F.; Hsu, L.; Trask, B.J. Extreme variability among mammalian V1R gene families. Genome Res. 2010, 20, 10–18. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, I.; Del Punta, K.; Rothman, A.; Ishii, T.; Mombaerts, P. Multiple new and isolated families within the mouse superfamily of V1r vomeronasal receptors. Nat. Neurosci. 2002, 5, 134–140. [Google Scholar] [CrossRef]
- Grus, W.E.; Zhang, J. Rapid turnover and species-specificity of vomeronasal pheromone receptor genes in mice and rats. Gene 2004, 340, 303–312. [Google Scholar] [CrossRef]
- Meisami, E.; Bhatnagar, K.P. Structure and diversity in mammalian accessory olfactory bulb. Microsc. Res. Tech. 1998, 43, 476–499. [Google Scholar] [CrossRef]
- Salazar, I.; Sánchez-Quinteiro, P.; Alemañ, N.; Cifuentes, J.M.; Troconiz, P.F. Diversity of the vomeronasal system in mammals: The singularities of the sheep model. Microsc. Res. Tech. 2007, 70, 752–762. [Google Scholar] [CrossRef]
- Hughes, L. Meerkats: Essential Wildlife; Character-19 ed.: Swindon, UK, 2020. [Google Scholar]
- Mausbach, J.; Goncalves, I.B.; Heistermann, M.; Ganswindt, A.; Manser, M.B. Meerkat close calling patterns are linked to sex, social category, season and wind, but not fecal glucocorticoid metabolite concentrations. PLoS ONE 2017, 12, e0175371. [Google Scholar] [CrossRef] [PubMed]
- Wadewitz, P. Olfactory Discrimination of Predators and Conspecifics in Meerkats (Suricata suricatta). Master’s Thesis, University of Zurich, Zurich, Switzerland, 2010. [Google Scholar]
- Hollén, L.I.; Manser, M.B. Persistence of alarm-call behaviour in the absence of predators: A comparison between wild and captive-born meerkats (Suricata suricatta). Ethology 2007, 113, 1038–1047. [Google Scholar] [CrossRef] [Green Version]
- Doolan, S.P.; Macdonald, D.W. Diet and foraging behaviour of group-living meerkats, Suricata suricatta, in the southern Kalahari. J. Zool. 1996, 239, 697–716. [Google Scholar] [CrossRef]
- Ewer, R.F. The behaviour of the meerkat, Suricata suricatta (Schreber). Z. Tierpsychol. 1963, 20, 570–607. [Google Scholar] [CrossRef]
- Sörensen, I.; Amundin, M.; Laska, M. Meerkats (Suricata suricatta) are able to detect hidden food using olfactory cues alone. Physiol. Behav. 2019, 202, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Coombes, H.A.; Stockley, P.; Hurst, J.L. Female chemical signalling underlying reproduction in mammals. J. Chem. Ecol. 2018, 44, 851–873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jordan, N.R.; Cherry, M.I.; Manser, M.B. Latrine distribution and patterns of use by wild meerkats: Implications for territory and mate defence. Anim. Behav. 2007, 73, 613–622. [Google Scholar] [CrossRef]
- Jordan, N.R. Scent-marking investment is determined by sex and breeding status in meerkats. Anim. Behav. 2007, 74, 531–540. [Google Scholar] [CrossRef]
- Mares, R.; Young, A.J.; Levesque, D.L.; Harrison, N.; Clutton-Brock, T.H. Responses to intruder scents in the cooperatively breeding meerkat: Sex and social status differences and temporal variation. Behav. Ecol. 2011, 22, 594–600. [Google Scholar] [CrossRef] [Green Version]
- Leclaire, S.; Jacob, S.; Greene, L.K.; Dubay, G.R.; Drea, C.M. Social odours covary with bacterial community in the anal secretions of wild meerkats. Sci. Rep. 2017, 7, 3240. [Google Scholar] [CrossRef]
- Leclaire, S.; Nielsen, J.F.; Thavarajah, N.K.; Manser, M.; Clutton-Brock, T.H. Odour-based kin discrimination in the cooperatively breeding meerkat. Biol. Lett. 2013, 9, 20121054. [Google Scholar] [CrossRef] [Green Version]
- Moran, G.; Sorensen, L. Scent marking behavior in a captive group of meerkats (Suricata suricatta). J. Mammal. 1986, 67, 120–132. [Google Scholar] [CrossRef]
- Zöttl, M.; Lienert, R.; Clutton-Brock, T.; Millesi, E.; Manser, M.B. The effects of recruitment to direct predator cues on predator responses in meerkats. Behav. Ecol. 2013, 24, 198–204. [Google Scholar] [CrossRef] [Green Version]
- Willey, T.J. The ultrastructure of the cat olfactory bulb. J. Comp. Neurol. 1973, 152, 211–232. [Google Scholar] [CrossRef] [PubMed]
- Nadi, N.S.; Hirsch, J.D.; Margolis, F.L. Laminar distribution of putative neurotransmitter amino acids and ligand binding sites in the dog olfactory bulb. J. Neurochem. 1980, 34, 138–146. [Google Scholar] [CrossRef]
- Choi, J.H.; Lee, C.H.; Yoo, K.Y.; Hwang, I.K.; Lee, I.S.; Lee, Y.L.; Shin, H.C.; Won, M.H. Age-related changes in calbindin-D28k, parvalbumin, and calretinin immunoreactivity in the dog main olfactory bulb. Cell. Mol. Neurobiol. 2010, 30, 1–12. [Google Scholar] [CrossRef]
- Chengetanai, S.; Bhagwandin, A.; Bertelsen, M.F.; Hård, T.; Hof, P.R.; Spocter, M.A.; Manger, P.R. The brain of the African wild dog. II. The olfactory system. J. Comp. Neurol. 2020, 528, 3285–3304. [Google Scholar] [CrossRef]
- Imam, A.; Bhagwandin, A.; Ajao, M.S.; Spocter, M.A.; Ihunwo, A.O.; Manger, P.R. The brain of the tree pangolin (Manis tricuspis). II. The olfactory system. J. Comp. Neurol. 2018, 526, 2548–2569. [Google Scholar] [CrossRef]
- Das, D.; Ghosh, D.; Payel, P.; Mandal, P. Handbook on Bio-Staining Procedures: A Quick Practical Approach; OrangeBooks Publication: Chhattisgarh, India, 2020; ISBN 978-93-90169-51-1. [Google Scholar]
- Shinohara, H.; Asano, T.; Kato, K. Differential localization of G-proteins Gi and Go in the accessory olfactory bulb of the rat. J. Neurosci. 1992, 12, 1275–1279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, C.; Halpern, M. Subclasses of vomeronasal receptor neurons: Differential expression of G proteins (Gi alpha 2 and G (o alpha)) and segregated projections to the accessory olfactory bulb. Brain Res. 1996, 719, 117–128. [Google Scholar] [CrossRef]
- Johnson, G.V.; Jope, R.S. The role of microtubule-associated protein 2 (MAP-2) in neuronal growth, plasticity, and degeneration. J. Neurosci. Res. 1992, 33, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Lee, V.M.-Y.; Otvos, L., Jr.; Carden, M.J.; Hollosi, M.; Dietzschold, B.; Lazzarini, R.A. Identification of the major multiphosphorylation site in mammalian neurofilaments. Proc. Natl. Acad. Sci. USA 1988, 85, 1998–2002. [Google Scholar] [CrossRef] [Green Version]
- Paxinos, G.; Kus, L.; Ashwell, K.W.S.; Watson, C.R.R. Chemoarchitectonic Atlas of the Rat Forebrain; Academic Press: San Diego, CA, USA, 2008. [Google Scholar]
- Verhaagen, J.; Oestreicher, A.B.; Gispen, W.H.; Margolis, F.L. The expression of the growth associated protein B50/GAP43 in the olfactory system of neonatal and adult rats. J. Neurosci. 1989, 9, 683–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodewald, A.; Gisder, D.; Gebhart, V.M.; Oehring, H.; Jirikowski, G.F. Distribution of olfactory marker protein in the rat vomeronasal organ. J. Chem. Neuroanat. 2016, 77, 19–23. [Google Scholar] [CrossRef]
- Kishimoto, J.; Keverne, E.B.; Emson, P.C. Calretinin, calbindin-D28k and parvalbumin-like immunoreactivity in mouse chemoreceptor neurons. Brain Res. 1993, 610, 325–329. [Google Scholar] [CrossRef]
- Bailey, M.S.; Shipley, M.T. Astrocyte subtypes in the rat olfactory bulb: Morphological heterogeneity and differential laminar distribution. J. Comp. Neurol. 1993, 328, 501–526. [Google Scholar] [CrossRef] [PubMed]
- Alroy, J.; Ucc, A.A.; Goyal, V.; Woods, W. Lectin histochemistry of glycolipid storage diseases on frozen and paraffin-embedded tissue sections. J. Histochem. Cytochem. 1986, 34, 501–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salazar, I.; Cifuentes, J.M.; Sánchez-Quinteiro, P. Morphological and immunohistochemical features of the vomeronasal system in dogs. Anat. Rec. 2013, 296, 146–155. [Google Scholar] [CrossRef] [PubMed]
- Salazar, I.; Sánchez-Quinteiro, P. Differential development of binding sites for four lectins in the vomeronasal system of juvenile mouse: From the sensory transduction site to the first relay stage. Brain Res. 2003, 979, 15–26. [Google Scholar] [CrossRef]
- Franceschini, V.; Lazzari, M.; Ciani, F. Lectin cytochemical localisation of glycoconjugates in the olfactory system of the lizards Lacerta viridis and Podarcis sicula. Anat. Embryol. 2000, 202, 49–54. [Google Scholar] [CrossRef]
- Peters, B.P.; Goldstein, I.J. The use of fluorescein-conjugated Bandeiraea simplicifolia B4-isolectin as a histochemical reagent for the detection of alpha-D-galactopyranosyl groups. Their occurrence in basement membranes. Exp. Cell Res. 1979, 120, 321–334. [Google Scholar] [CrossRef] [Green Version]
- Salazar, I.; Sánchez-Quinteiro, P. Lectin binding patterns in the vomeronasal organ and accessory olfactory bulb of the rat. Anat. Embryol. 1998, 198, 331–339. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, L.S.; Ee, P.L.; Halpern, M. Lectin histochemical identification of carbohydrate moieties in opossum chemosensory systems during development, with special emphasis on VVA-identified subdivisions in the accessory olfactory bulb. J. Morphol. 1995, 224, 331–349. [Google Scholar] [CrossRef]
- Pillay, S.; Bhagwandin, A.; Bertelsen, M.F.; Patzke, N.; Engler, G.; Engel, A.K.; Manger, P.R. Regional distribution of cholinergic, catecholaminergic, serotonergic and orexinergic neurons in the brain of two carnivore species: The feliform banded mongoose (Mungos mungo) and the caniform domestic ferret (Mustela putorius furo). J. Chem. Neuroanat. 2017, 82, 12–28. [Google Scholar] [CrossRef]
- Salazar, I.; Sánchez-Quinteiro, P. The risk of extrapolation in neuroanatomy: The case of the mammalian vomeronasal system. Front. Neuroanat. 2009, 3, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorman, M.L.; Trowbridge, B.J. The Role of Odor in the Social Lives of Carnivores. In Carnivore Behavior, Ecology, and Evolution; Gittleman, J.L., Ed.; Cornell University Press: Ithaca, NY, USA, 2019; pp. 57–88. [Google Scholar] [CrossRef]
- Alonso, J.R.; Porteros, A.; Crespo, C.; Arévalo, R.; Briñón, J.G.; Weruaga, E.; Aijón, J. Chemical anatomy of the macaque monkey olfactory bulb: NADPH-diaphorase/nitric oxide synthase activity. J. Comp. Neurol. 1998, 402, 419–434. [Google Scholar] [CrossRef]
- Kondoh, D.; Watanabe, K.; Nishihara, K.; Ono, Y.S.; Nakamura, K.G.; Yuhara, K.; Tomikawa, S.; Sugimoto, M.; Kobayashi, S.; Horiuchi, N.; et al. Histological properties of main and accessory olfactory bulbs in the common hippopotamus. Brain Behav. Evol. 2017, 90, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Kishida, T.; Thewissen, J.; Usip, S.; Suydam, R.S.; George, J.C. Organization and distribution of glomeruli in the bowhead whale olfactory bulb. PeerJ 2015, 3, e897. [Google Scholar] [CrossRef] [Green Version]
- Ngwenya, A.; Patzke, N.; Ihunwo, A.O.; Manger, P.R. Organisation and chemical neuroanatomy of the African elephant (Loxodonta africana) olfactory bulb. Brain. Struct. Funct. 2011, 216, 403–416. [Google Scholar] [CrossRef]
- Schröder, H.; Moser, N.; Huggenberger, S. The mouse olfactory system. In Neuroanatomy of the Mouse; Springer: Cham, Switzerland, 2020. [Google Scholar] [CrossRef]
- Orechio, D.; Aguiar, B.A.; Diniz, G.B.; Bittencourt, J.C.; Haemmerle, C.A.S.; Watanabe, I.S.; Miglino, M.A.; Castelucci, P. Morphological and cellular characterization of the fetal canine (Canis lupus familiaris) subventricular zone, rostral migratory stream, and olfactory bulb. Anat. Rec. 2018, 301, 1570–1584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández-Flores, F.; García-Verdugo, J.M.; Martín-Ibáñez, R.; Herranz, C.; Fondevila, D.; Canals, J.M.; Arús, C.; Pumarola, M. Characterization of the canine rostral ventricular-subventricular zone: Morphological, immunohistochemical, ultrastructural, and neurosphere assay studies. J. Comp. Neurol. 2018, 526, 721–741. [Google Scholar] [CrossRef]
- Alvarez-Buylla, A.; Garcia-Verdugo, J.M. Neurogenesis in adult subventricular zone. J. Neurosci. 2002, 22, 629–634. [Google Scholar] [CrossRef] [Green Version]
- Sohn, J.; Orosco, L.; Guo, F.; Chung, S.H.; Bannerman, P.; Mills Ko, E.; Zarbalis, K.; Deng, W.; Pleasure, D. The subventricular zone continues to generate corpus callosum and rostral migratory stream astroglia in normal adult mice. J. Neurosci. 2015, 35, 3756–3763. [Google Scholar] [CrossRef] [Green Version]
- Doetsch, F.; Alvarez-Buylla, A. Network of tangential pathways for neuronal migration in adult mammalian brain. Proc. Natl. Acad. Sci. USA 1996, 93, 14895–14900. [Google Scholar] [CrossRef] [Green Version]
- Cayre, M.; Canoll, P.; Goldman, J.E. Cell migration in the normal and pathological postnatal mammalian brain. Prog. Neurobiol. 2009, 88, 41–63. [Google Scholar] [CrossRef] [Green Version]
- Kondoh, D.; Sasaki, M.; Kitamura, N. Age-dependent decrease in glomeruli and receptor cells containing α1-2 fucose glycan in the mouse main olfactory system but not in the vomeronasal system. Cell Tissue Res. 2018, 373, 361–366. [Google Scholar] [CrossRef] [PubMed]
- Jia, C.; Halpern, M. Calbindin D28k, parvalbumin, and calretinin immunoreactivity in the main and accessory olfactory bulbs of the gray short-tailed opossum, Monodelphis domestica. J. Morphol. 2004, 259, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Schneider, N.Y.; Fletcher, T.P.; Shaw, G.; Renfree, M.B. Goα expression in the vomeronasal organ and olfactory bulb of the tammar wallaby. Chem. Senses 2012, 37, 567–577. [Google Scholar] [CrossRef] [Green Version]
- Torres, M.V.; Ortiz-Leal, I.; Villamayor, P.R.; Ferreiro, A.; Rois, J.L.; Sánchez-Quinteiro, P. Does a third intermediate model for the vomeronasal processing of information exist? Insights from the macropodid neuroanatomy. Brain Struct. Funct. 2021. [Google Scholar] [CrossRef]
- Skeen, L.C.; Hall, W.C. Efferent projections of the main and the accessory olfactory bulb in the tree shrew (Tupaia glis). J. Comp. Neurol. 1977, 172, 1–35. [Google Scholar] [CrossRef] [PubMed]
- Park, C.; Ahn, M.; Lee, J.Y.; Lee, S.; Yun, Y.; Lim, Y.K.; Taniguchi, K.; Shin, T. A morphological study of the vomeronasal organ and the accessory olfactory bulb in the Korean roe deer, Capreolus pygargus. Acta Histochem. 2014, 116, 258–264. [Google Scholar] [CrossRef]
- Mackay-Sim, A.; Duvall, D.; Graves, B.M. The West Indian manatee (Trichechus manatus) lacks a vomeronasal organ. Brain Behav. Evol. 1985, 27, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Trotier, D.; Eloit, C.; Wassef, M.; Talmain, G.; Bensimon, J.L.; Døving, K.B.; Ferrand, J. The vomeronasal cavity in adult humans. Chem. Senses 2000, 25, 369–380. [Google Scholar] [CrossRef] [Green Version]
- Salazar, I.; Sánchez-Quinteiro, P.; Cifuentes, J.M.; Lombardero, M. The accessory olfactory bulb of the mink, Mustela vison: A morphological and lectin histochemical study. Anat. Histol. Embryol. 1998, 27, 297–300. [Google Scholar] [CrossRef] [PubMed]
- Kelliher, K.R.; Baum, M.J.; Meredith, M. The ferret’s vomeronasal organ and accessory olfactory bulb: Effect of hormone manipulation in adult males and females. Anat. Rec. 2001, 263, 280–288. [Google Scholar] [CrossRef]
- Frahm, H.D.; Bhatnagar, K.P. Comparative morphology of the accessory olfactory bulb in bats. J. Anat. 1980, 130, 349–365. [Google Scholar]
- Salazar, I.; Sánchez-Quinteiro, P. A detailed morphological study of the vomeronasal organ and the accessory olfactory bulb of cats. Microsc. Res. Tech. 2011, 74, 1109–1120. [Google Scholar] [CrossRef] [PubMed]
- Salazar, I.; Cifuentes, J.M.; Sánchez-Quinteiro, P.; García Caballero, T. Structural, morphometric, and immunohistological study of the accessory olfactory bulb in the dog. Anat. Rec. 1994, 240, 277–285. [Google Scholar] [CrossRef]
- Nakajima, T.; Sakaue, M.; Kato, M.; Saito, S.; Ogawa, K.; Taniguchi, K. Immunohistochemical and enzyme-histochemical study on the accessory olfactory bulb of the dog. Anat. Rec. 1998, 252, 393–402. [Google Scholar] [CrossRef]
- Riddell, P.; Paris, M.C.J.; Joonè, C.J.; Pageat, P.; Paris, D.B.B.P. Appeasing pheromones for the management of stress and aggression during conservation of wild canids: Could the solution be right under our nose? Animals 2021, 11, 1574. [Google Scholar] [CrossRef]
- Pageat, P.; Gaultier, E. Current research in canine and feline pheromones. Vet. Clin. North Am. Small Anim. Pract. 2003, 33, 187–211. [Google Scholar] [CrossRef]
- Kaminski, J.; Marshall-Pescini, S. The Social Dog: Behaviour and Cognition; Academic Press: San Diego, CA, USA, 2014. [Google Scholar]
- Cajal, S.R. Textura del lobulo olfativo accesorio. Rev. Micros. 1902, 1, 141–150. [Google Scholar]
- Jawlowski, H. On the bulbus olfactorius and bulbus olfactorius accessorius of some mammals. Lub. Uniw. Marii Curie-Sklodowskiej Roczniki. Annls Dzialc Nauk. Biol. 1956, 10, 67–86. [Google Scholar]
- Miodonski, R. Bulbus olfactorius of the dog (Canis familiaris). Acta Biol. Cracov. 1968, 11, 65–75. [Google Scholar]
- Salazar, I.; Barber, P.C.; Cifuentes, J.M. Anatomical and immunohistological demonstration of the primary neural connections of the vomeronasal organ in the dog. Anat. Rec. 1992, 233, 309–313. [Google Scholar] [CrossRef]
- Broom, R. A contribution to the comparative anatomy of the mammalian organ of Jacobson. Trans. R. Soc. Edinb. 1897, 39, 231–255. [Google Scholar] [CrossRef] [Green Version]
- Wöhrmann-Repenning, A. Functional aspects of the vomeronasal complex in mammals. Zool. Jb. Anat. 1991, 121, 71–80. [Google Scholar]
- Sanchez-Villagra, M.R. Ontogenetic and phylogenetic transformations of the vomeronasal complex and nasal floor elements in marsupial mammals. Zool. J. Linnean Soc. 2001, 131, 459–479. [Google Scholar] [CrossRef]
- Dulac, C.; Torello, A.T. Molecular detection of pheromone signals in mammals: From genes to behaviour. Nat. Rev. Neurosci. 2003, 4, 551–562. [Google Scholar] [CrossRef]
- Suarez, R.; Mpodozis, J. Heterogeneities of size and sexual dimorphism between the subdomains of the lateral-innervated accessory olfactory bulb (AOB) of Octodon degus (Rodentia: Hystricognathi). Behav. Brain Res. 2009, 198, 306–312. [Google Scholar] [CrossRef]
- Suarez, R.; Santibáñez, R.; Parra, D.; Coppi, A.A.; Abrahão, L.M.B.; Sasahara, T.H.C.; Mpodozis, J. Share and differential traits in the accessory olfactory bulb of caviomorph rodents with particular reference to the semiaquatic capybara. J. Anat. 2011, 218, 558–565. [Google Scholar] [CrossRef] [Green Version]
- Torres, M.V.; Ortiz-Leal, I.; Villamayor, P.R.; Ferreiro, A.; Rois, J.L.; Sánchez-Quinteiro, P. The vomeronasal system of the newborn capybara: A morphological and immunohistochemical study. Sci. Rep. 2020, 10, 13304. [Google Scholar] [CrossRef]
- Halpern, M.; Shapiro, L.S.; Jia, C. Differential localization of G proteins in the opossum vomeronasal system. Brain Res. 1995, 677, 157–161. [Google Scholar] [CrossRef]
- Suarez, R.; Villalón, A.; Künzle, H.; Mpodozis, J. Transposition and Intermingling of Galphai2 and Galphao afferences into single vomeronasal glomeruli in the Madagascan lesser Tenrec Echinops telfairi. PLoS ONE 2009, 4, e8005. [Google Scholar] [CrossRef] [Green Version]
- Ortiz-Leal, I.; Torres, M.V.; Villamayor, P.R.; Fidalgo, L.; López-Beceiro, A.; Sanchez-Quinteiro, P. Can domestication shape canidae brain morphology? The accessory olfactory bulb of the red fox as a case in point. Ann. Anat 2021, 240, 151881. [Google Scholar] [CrossRef] [PubMed]
- Takigami, S.; Mori, Y.; Ichikawa, M. Projection pattern of vomeronasal neurons to the accessory olfactory bulb in goats. Chem. Senses 2000, 25, 387–393. [Google Scholar] [CrossRef]
- Takigami, S.; Mori, Y.; Tanioka, Y.; Ichikawa, M. Morphological evidence for two types of mammalian vomeronasal system. Chem. Senses 2004, 29, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Suarez, R.; Fernandez-Aburto, P.; Manger, P.R.; Mpodozis, J. Deterioration of the Gαo vomeronasal pathway in sexually dimorphic mammals. PLoS ONE 2011, 6, e26436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wekesa, K.S.; Anholt, R.R. Differential expression of G proteins in the mouse olfactory system. Brain Res. 1999, 837, 117–126. [Google Scholar] [CrossRef]
- Salazar, I.; Barrios, A.W.; Sánchez-Quinteiro, P. Revisiting the vomeronasal system from an integrated perspective. Anat. Rec. 2016, 299, 1488–1491. [Google Scholar] [CrossRef]
- Bock, P.; Rohn, K.; Beineke, A.; Baumgärtner, W.; Wewetzer, K. Site-specific population dynamics and variable olfactory marker protein expression in the postnatal canine olfactory epithelium. J. Anat. 2009, 215, 522–535. [Google Scholar] [CrossRef]
- Villamayor, P.R.; Cifuentes, J.M.; Quintela, L.; Barcia, R.; Sánchez-Quinteiro, P. Structural, morphometric and immunohistochemical study of the rabbit accessory olfactory bulb. Brain Struct. Funct. 2020, 225, 203–226. [Google Scholar] [CrossRef]
- Shnayder, L.; Schwanzel-Fukuda, M.; Halpern, M. Differential OMP expression in opossum accessory olfactory bulb. Neuroreport 1993, 5, 193–196. [Google Scholar] [CrossRef]
- Ramakers, G.J.; Verhaagen, J.; Oestreicher, A.B.; Margolis, F.L.; van Bergen en Henegouwen, P.M.; Gispen, W.H. Immunolocalization of B-50 (GAP-43) in the mouse olfactory bulb: Predominant presence in preterminal axons. J. Neurocytol. 1992, 21, 853–869. [Google Scholar] [CrossRef]
- Salazar, I.; Sánchez-Quinteiro, P.; Cifuentes, J.M.; Troconiz, P.F. General organization of the perinatal and adult accessory olfactory bulb in mice. Anat. Rec. 2006, 288, 1009–1025. [Google Scholar] [CrossRef] [PubMed]
- Dehmelt, L.; Halpain, S. The MAP2/Tau family of microtubule-associated proteins. Genome Biol. 2005, 6, 204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galliano, E.; Hahn, C.; Browne, L.P.; Villamayor, P.R.; Tufo, C.; Crespo, A.; Grubb, M.S. Brief sensory deprivation triggers cell type-specific structural and functional plasticity in olfactory bulb neurons. J. Neurosci. 2021, 41, 2135–2151. [Google Scholar] [CrossRef]
- Ashwell, K.W. Chemoarchitecture of the monotreme olfactory bulb. Brain Behav. Evol. 2006, 67, 69–84. [Google Scholar] [CrossRef] [PubMed]
- Allen, N.J.; Lyons, D.A. Glia as architects of central nervous system formation and function. Science 2018, 362, 181–185. [Google Scholar] [CrossRef] [Green Version]
- Lazzari, M.; Bettini, S.; Franceschini, V. Immunocytochemical characterisation of ensheathing glia in the olfactory and vomeronasal systems of Ambystoma mexicanum (Caudata: Ambystomatidae). Brain Struct. Funct. 2016, 221, 955–967. [Google Scholar] [CrossRef]
- Smithson, L.J.; Kawaja, M.D. A comparative examination of biomarkers for olfactory ensheathing cells in cats and guinea pigs. Brain Res. 2009, 1284, 41–53. [Google Scholar] [CrossRef]
- Jacobowitz, D.M.; Winsky, L. Immunocytochemical localization of calretinin in the forebrain of the rat. J. Comp. Neurol. 1991, 304, 198–218. [Google Scholar] [CrossRef]
- Porteros, A.; Arévalo, R.; Crespo, C.; García-Ojeda, E.; Briñón, J.G.; Aijón, J.; Alonso, J.R. Calbindin D-28k immunoreactivity in the rat accessory olfactory bulb. Brain Res. 1995, 689, 93–100. [Google Scholar] [CrossRef] [Green Version]
- Jia, C.; Halpern, M. Calbindin D28K immunoreactive neurons in vomeronasal organ and their projections to the accessory olfactory bulb in the rat. Brain Res. 2003, 977, 261–269. [Google Scholar] [CrossRef]
- Larriva-Sahd, J. Cytological organization of the alpha component of the anterior olfactory nucleus and olfactory limbus. Front. Neuroanat. 2012, 6, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shinoda, K.; Ohtsuki, T.; Nagano, M.; Okumura, T. A possible functional necklace formed by placental antigen X-P2-immunoreactive and intensely acetylcholinesterase-reactive (PAX/IAE) glomerular complexes in the rat olfactory bulb. Brain Res. 1993, 618, 160–166. [Google Scholar] [CrossRef]
- Zheng, L.M.; Ravel, N.; Jourdan, F. Topography of centrifugal acetylcholinesterase-positive fibres in the olfactory bulb of the rat: Evidence for original projections in atypical glomeruli. Neuroscience 1987, 23, 1083–1093. [Google Scholar] [CrossRef]
- Kinzinger, J.H.; Johnson, E.W.; Bhatnagar, K.P.; Bonar, C.J.; Burrows, A.M.; Mooney, M.P.; Siegel, M.I.; Smith, T.D. Comparative study of lectin reactivity in the vomeronasal organ of human and nonhuman primates. Anat. Rec. A Discov. Mol. Cell. Evol. Biol. 2005, 284, 550–560. [Google Scholar] [CrossRef] [PubMed]
- Salazar, I.; Sánchez-Quinteiro, P.; Lombardero, M.; Cifuentes, J.M. Histochemical identification of carbohydrate moieties in the accessory olfactory bulb of the mouse using a panel of lectins. Chem. Senses 2001, 26, 645–652. [Google Scholar] [CrossRef] [Green Version]
- Salazar, I.; Sánchez-Quinteiro, P.; Lombardero, M.; Cifuentes, J.M. A descriptive and comparative lectin histochemical study of the vomeronasal system in pigs and sheep. J. Anat. 2000, 196, 15–22. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ab/Lectin | 1st Ab Species/Dilution | 1st Ab Catalogue Number | 2nd Ab Species/Dilution (Catalogue Number) |
|---|---|---|---|
| Anti-Gαo | Rabbit 1:100 | MBL 551 | ImmPRESS VR HRP Anti-Rabbit IgG MP-6401-15 |
| Anti-Gαi2 | Rabbit 1:100 | Sta Cruz SC-7276 | ImmPRESS VR HRP Anti-Rabbit IgG MP-6401-15 |
| Anti-OMP | Goat 1:400 | Wako S44-10001 | Horse 1:250 Vector BA-9500 |
| Anti-MAP2 | Mouse 1:200 | Sigma M4403 | ImmPRESS VR HRP Anti-Mouse IgG MP-6402-15 |
| Anti-GAP43 | Mouse 1:800 | Sigma G9264 | ImmPRESS VR HRP Anti-Mouse IgG MP-6402-15 |
| Anti-GFAP | Rabbit 1:400 | Dako Z0334 | ImmPRESS VR HRP Anti-Rabbit IgG MP-6401-15 |
| Anti-Calbindin | Rabbit 1:5000 | Swant CB38 | ImmPRESS VR HRP Anti-Rabbit IgG MP-6401-15 |
| Anti-Calretinin | Rabbit 1:5000 | Swant 7697 | ImmPRESS VR HRP Anti-Rabbit IgG MP-6401-15 |
| Anti-SMI32 | Rabbit 1:20 | ENZ-ABS219-0100 | ImmPRESS VR HRP Anti-Rabbit IgG MP-6401-15 |
| UEA-I | 1:10 | Vector L-1060 | Rabbit 1:50 DAKO P289 |
| LEA | 20 μg/mL | Vector B-1175 | Vectastain ABC reagent PK-4000 |
| BSI-B4 | 100 µm/mL | Sigma L-2140 | Vectastain ABC reagent PK-4000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Torres, M.V.; Ortiz-Leal, I.; Ferreiro, A.; Rois, J.L.; Sanchez-Quinteiro, P. Neuroanatomical and Immunohistological Study of the Main and Accessory Olfactory Bulbs of the Meerkat (Suricata suricatta). Animals 2022, 12, 91. https://doi.org/10.3390/ani12010091
Torres MV, Ortiz-Leal I, Ferreiro A, Rois JL, Sanchez-Quinteiro P. Neuroanatomical and Immunohistological Study of the Main and Accessory Olfactory Bulbs of the Meerkat (Suricata suricatta). Animals. 2022; 12(1):91. https://doi.org/10.3390/ani12010091
Chicago/Turabian StyleTorres, Mateo V., Irene Ortiz-Leal, Andrea Ferreiro, José Luis Rois, and Pablo Sanchez-Quinteiro. 2022. "Neuroanatomical and Immunohistological Study of the Main and Accessory Olfactory Bulbs of the Meerkat (Suricata suricatta)" Animals 12, no. 1: 91. https://doi.org/10.3390/ani12010091