
The Ecological Roles of Medium and Small Carnivores in the Terrestrial Animal Community in Liancheng National Nature Reserve, China

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Data Collection
2.3. Data Analysis
2.3.1. Analysis of Species Spatial Associations
2.3.2. Characterization of Species Spatial Associations
2.3.3. Species Contribution to Spatial Network Complexity
3. Results
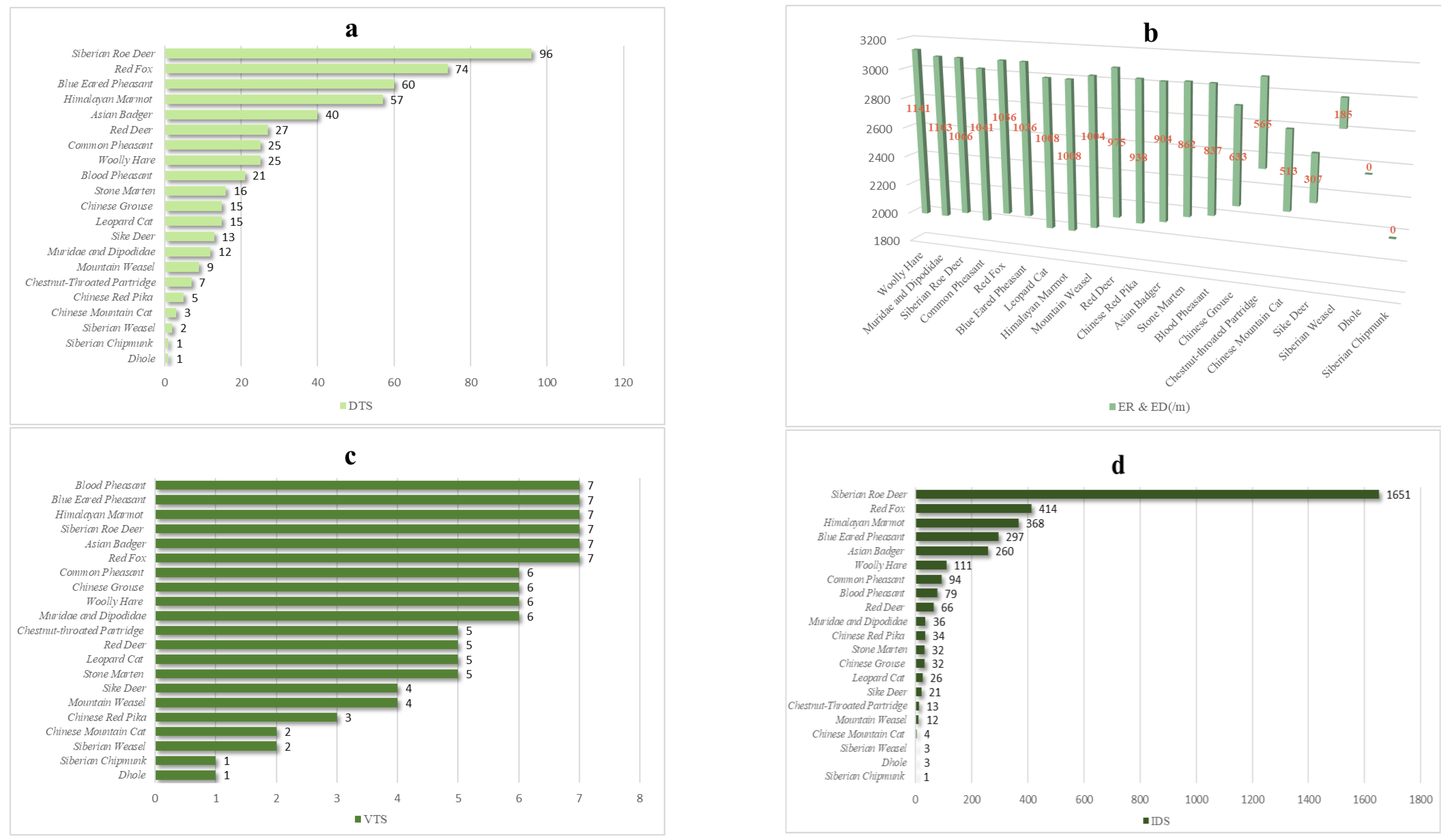
3.1. Diversity of Species
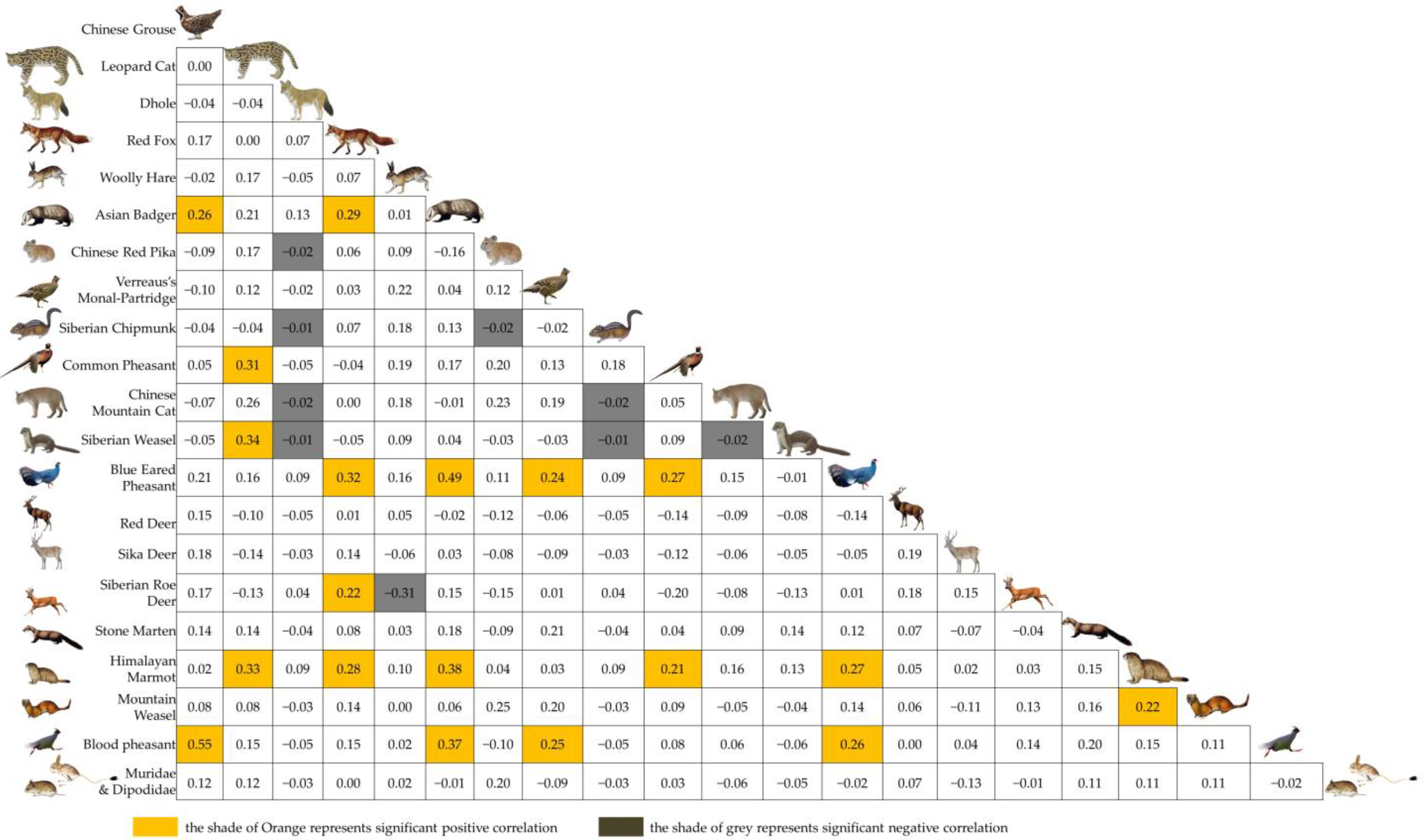
3.2. Spatial Associations of Medium and Small Carnivores
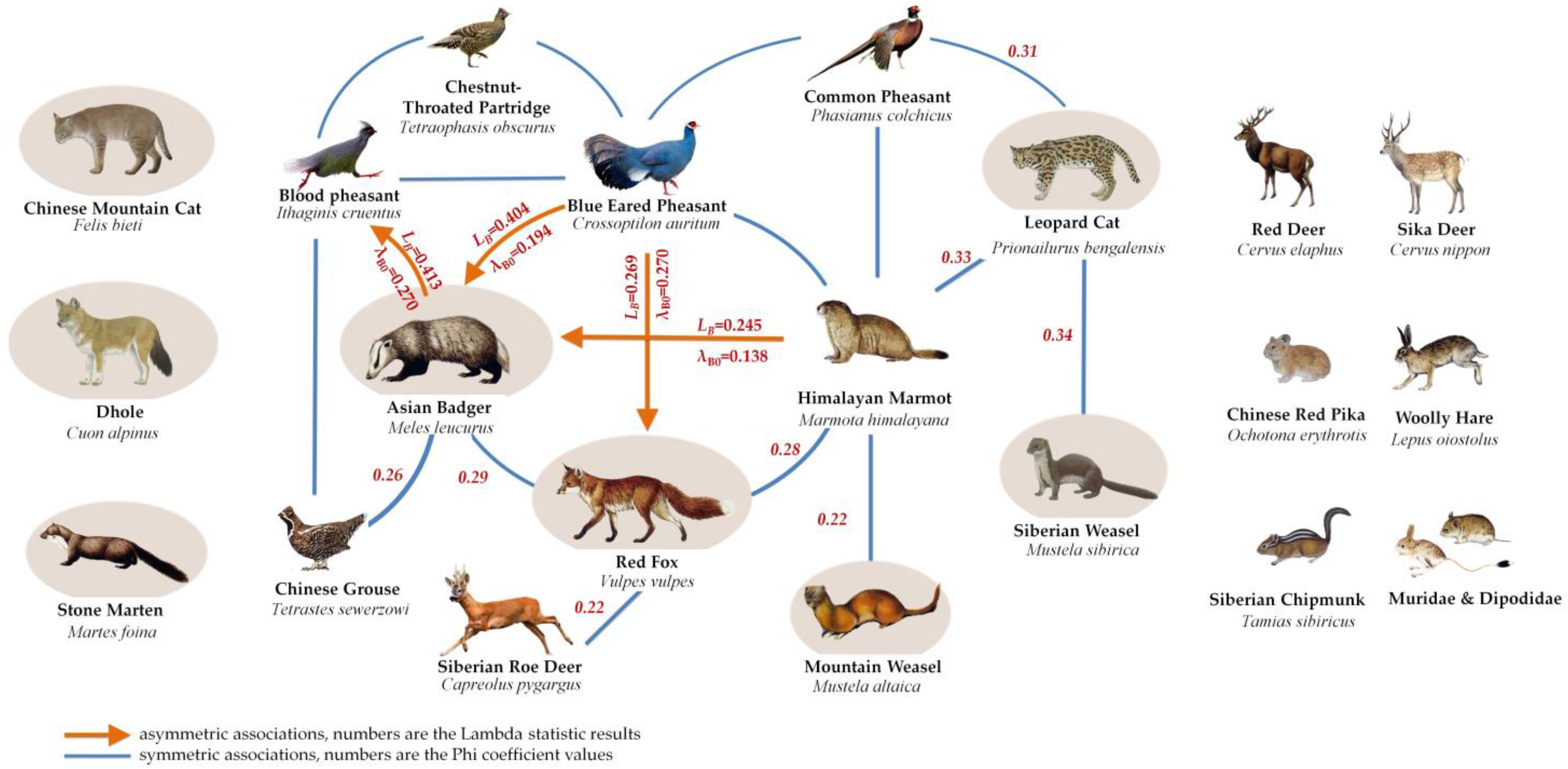
3.3. Characterization of Spatial Associations of Medium and Small Carnivores
3.4. Contribution of Medium and Small Carnivores to Network Maintenance
4. Discussion
4.1. Status of Species Diversity in the LNNR
4.2. Medium and Small Carnivores Cannot Replace the Ecological Roles of Large Carnivores
4.3. Ecological Role of Medium and Small Carnivores
4.4. Community Development Trends
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fryxell, J.M.; Mosser, A.; Sinclair, A.R.E.; Packer, C. Group Formation Stabilizes Predator-Prey Dynamics. Nature 2007, 449, 1041–1043. [Google Scholar] [CrossRef] [PubMed]
- Lyngdoh, S.; Shrotriya, S.; Goyal, S.P.; Clements, H.; Hayward, M.W.; Habib, B. Prey Preferences of the Snow Leopard (Panthera uncia): Regional Diet Specificity Holds Global Significance for Conservation. PLoS ONE 2014, 9, e100071. [Google Scholar] [CrossRef] [PubMed]
- Ripple, W.J.; Beschta, R.L. Large Predators Limit Herbivore Densities in Northern Forest Ecosystems. Eur. J. Wildl. Res. 2012, 58, 733–742. [Google Scholar] [CrossRef]
- Ritchie, E.G.; Elmhagen, B.; Glen, A.S.; Letnic, M.; Ludwig, G.; Mcdonald, R.A. Ecosystem Restoration with Teeth: What Role for Predators ? Trends Ecol. Evol. 2012, 27, 265–271. [Google Scholar] [CrossRef]
- Suraci, J.P.; Clinchy, M.; Dill, L.M.; Roberts, D.; Zanette, L.Y. Fear of Large Carnivores Causes a Trophic Cascade. Nat. Commun. 2016, 7, 10698. [Google Scholar] [CrossRef] [Green Version]
- Sarasola, J.H.; Zanón-Martínez, J.I.; Costán, A.S.; Ripple, W.J. Hypercarnivorous Apex Predator Could Provide Ecosystem Services by Dispersing Seeds. Sci. Rep. 2016, 6, 19647. [Google Scholar] [CrossRef] [Green Version]
- Ripple, W.J.; Estes, J.A.; Beschta, R.L.; Wilmers, C.C.; Ritchie, E.G.; Hebblewhite, M.; Berger, J.; Elmhagen, B.; Letnic, M.; Nelson, M.P.; et al. Status and Ecological Effects of the World’s Largest Carnivores. Science 2014, 343, 1241484. [Google Scholar] [CrossRef] [Green Version]
- Prugh, L.R.; Stoner, C.J.; Epps, C.W.; Bean, W.T.; Ripple, W.J.; Laliberte, A.S.; Brashares, J.S. The Rise of the Mesopredator. Bioscience 2009, 59, 779–791. [Google Scholar] [CrossRef]
- Roemer, G.W.; Gompper, M.E.; Valkenburgh, B. Van The Ecological Role of the Mammalian Mesocarnivore. Bioscience 2009, 59, 165–173. [Google Scholar] [CrossRef]
- Wang, F.; Winkler, J.; Viña, A.; McShea, W.J.; Li, S.; Connor, T.; Zhao, Z.; Wang, D.; Yang, H.; Tang, Y.; et al. The Hidden Risk of Using Umbrella Species as Conservation Surrogates: A Spatio-Temporal Approach. Biol. Conserv. 2021, 253, 108913. [Google Scholar] [CrossRef]
- Putman, R.J. Community Ecology; Chapman & Hall: London, UK; New York, NY, USA, 1994. [Google Scholar]
- Tylianakis, J.M.; Laliberté, E.; Nielsen, A.; Bascompte, J. Conservation of Species Interaction Networks. Biol. Conserv. 2010, 143, 2270–2279. [Google Scholar] [CrossRef]
- Forbes, S.A. On the Local Distribution of Certain Illinois Fishes: An Essay in Statistical Ecology. Ill. Nat. Hist. Surv. Bull. 1907, 7, 273–297. [Google Scholar] [CrossRef]
- Yang, H.; Li, J.; Jiang, N.; Shi, X.; Wang, P.; Li, Z. Interspecific Associations Between Takins (Budorcas taxicolor) and Sympatric Terrestrial Mammals at Wolong National Nature Reserve, China. Chin. J. Wildl. 2021, 42, 654–662. [Google Scholar] [CrossRef]
- Zhou, H.; Jiang, N.; Li, J.; Yang, H.; Wu, Y.; Wang, P.; Shi, X.; LI, Z. Study on the Terrestrial Animal Community That Panthera Uncia Lived in Wolong National Nature Reserve, Sichuan Province, China. Chin. J. Wildl. 2021, 42, 645–653. [Google Scholar] [CrossRef]
- Li, Q.; Zhong, Y.; Zhou, H.; Liu, Z.; Wang, X.; Shi, X.; Li, Z. Analysis on Interspecific Association of Sambars Community in Wolong National Nature Reserve, Sichuan, China. Chin. J. Wildl. 2022, 43, 314–322. [Google Scholar] [CrossRef]
- Liu, Z.; Zhong, Y.; Wang, X.; Li, Q.; Zhou, H.; Shi, X.; Li, Z. Analysis on the Community Environment of Sichuan Snub-Nosed Monkeys (Rhinopithecus roxellana) in Wolong, China. Chin. J. Wildl. 2022, 43, 614–622. [Google Scholar] [CrossRef]
- Wang, X.; Man, Z.; Li, W.; Yang, J.; Zhao, Y.; Zhang, L.; Zhao, W. Camera Trapping Survey on Ground Dwelling Birds and Mammals in Liancheng National Nature Reserve, Gansu Province. Chin. J. Wildl. 2019, 40, 322–331. [Google Scholar] [CrossRef]
- O’Brien, T.G.; Kinnaird, M.F.; Wibisono, H.T. Crouching Tigers, Hidden Prey: Sumatran Tiger and Prey Populations in a Tropical Forest Landscape. Anim. Conserv. 2003, 6, 131–139. [Google Scholar] [CrossRef] [Green Version]
- Siegel, S.; John Castellan, J. Nonparametric Statistics for the Behavioral Sciences, 2nd ed.; McGraw-Hill International Editions: New York, NY, USA, 1988; ISBN 978-0070573574. [Google Scholar]
- Jun, C. Preliminary Investigation of Mammals and Their Habitats along Datong River in Yongdeng District, Lanzhou. Chin. J. Zool. 1960, 4, 104–107. [Google Scholar] [CrossRef]
- Xue, Y.; Li, J.; Hu, Y.; Ma, L.; Qian, W.; Yan, P.; Yang, M.; Chen, D.; Wu, B.; Li, D. Camera-Trapping Survey of the Mammals and Birds in the Qilian Moun_tain National Park (Qinghai Area), China. Acta Theriol. Sin. 2019, 39, 466–475. [Google Scholar] [CrossRef]
- Hu, D.; Xu, K.; Zhang, J.; Ma, D.; Ni, Z.; Xiao, Z. Wildlife Monitoring by Camera Trapping in the Qilianshan National Nature Reserve, Gansu Province. Chin. J. Wildl. 2022, 43, 692–703. [Google Scholar] [CrossRef]
- Xue, Y.; Li, D.; Xiao, W.; Zhang, Y.; Feng, B.; Jia, H. Records of the Dhole (Cuon alpinus) in an Arid Region of the Altun Mountains in Western China. Eur. J. Wildl. Res. 2015, 61, 903–907. [Google Scholar] [CrossRef]
- Liao, Y.F. Some Biologicai Informations of Desert Cat in Qinghai. Acta Theriol. Sin. 1988, 8, 128–131. [Google Scholar] [CrossRef]
- Wei, Z.; Liang, G.; Guangshuang, S. Winter Habitat Selection of Siberian Weasels in Changbaishan Area. Chin. J. Wildl. 2008, 29, 192–194. [Google Scholar] [CrossRef]
- Yang, H.; Rong, K. Wintering Habitat Selection of Siberian Chipmunk. Chin. J. Wildl. 2013, 34, 131–134. [Google Scholar] [CrossRef]
- Estes, J.A.; Terborgh, J.; Brashares, J.S.; Power, M.E.; Berger, J.; Bond, W.J.; Carpenter, S.R.; Essington, T.E.; Holt, R.D.; Jackson, J.B.C.; et al. Trophic Downgrading of Planet Earth. Science 2011, 333, 301–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beschta, R.L.; Ripple, W.J. Large Predators and Trophic Cascades in Terrestrial Ecosystems of the Western United States. Biol. Conserv. 2009, 142, 2401–2414. [Google Scholar] [CrossRef]
- Ripple, W.J.; Wirsing, A.J.; Wilmers, C.C.; Letnic, M. Widespread Mesopredator Effects after Wolf Extirpation. Biol. Conserv. 2013, 160, 70–79. [Google Scholar] [CrossRef] [Green Version]
- Ritchie, E.G.; Johnson, C.N. Predator Interactions, Mesopredator Release and Biodiversity Conservation. Ecol. Lett. 2009, 12, 982–998. [Google Scholar] [CrossRef]
- Capitani, C.; Bertelli, I.; Varuzza, P.; Scandura, M.; Apollonio, M. A Comparative Analysis of Wolf (Canis lupus) Diet in Three Different Italian Ecosystems. Mamm. Biol. 2004, 69, 1–10. [Google Scholar] [CrossRef]
- Xu, A.C.; Jiang, Z.G.; Li, C.W.; Cai, P. Food Habits and Hunting Patterns of Tibetan Brown Bear during Warm Seasons in Kekexili Region on Qinghai-Tibetan Plateau. Zool. Res. 2010, 31, 670–674. [Google Scholar] [CrossRef]
- Grassman, L.I.; Tewes, M.E.; Silvy, N.J.; Kreetiyutanont, K. Spatial ecology and diet of the dhole Cuon alpinus (Canidae, Carnivora) in north central Thailandt, K. Spatial Ecology and Diet of the Dhole Cuon alpinus (Canidae, Carnivora) in North Central Thailand. Mammalia 2005, 69, 11–20. [Google Scholar] [CrossRef]
- Thinley, P.; Kamler, J.F.; Wang, S.W.; Lham, K.; Stenkewitz, U.; Macdonald, D.W. Seasonal Diet of Dholes (Cuon alpinus) in Northwestern Bhutan. Mamm. Biol. 2011, 76, 518–520. [Google Scholar] [CrossRef]
- Krofel, M.; Huber, D.; Kos, I. Diet of Eurasian Lynx Lynx Lynx in the Northern Dinaric Mountains (Slovenia and Croatia). Acta Theriol. Warsz. 2011, 56, 315–322. [Google Scholar] [CrossRef]
- Kamler, J.F.; Johnson, A.; Vongkhamheng, C.; Bousa, A. The Diet, Prey Selection, and Activity of Dholes (Cuon alpinus) in Northern Laos. J. Mammal. 2012, 93, 627–633. [Google Scholar] [CrossRef] [Green Version]
- Mengüllüoğlu, D.; Ambarlı, H.; Berger, A.; Hofer, H. Foraging Ecology of Eurasian Lynx Populations in Southwest Asia: Conservation Implications for a Diet Specialist. Ecol. Evol. 2018, 8, 9451–9463. [Google Scholar] [CrossRef]
- Xiong, M.; Shao, X.; Long, Y.; Bu, H.; Zhang, D.; Wang, D.; Li, S.; Wang, R.; Yao, M. Molecular Analysis of Vertebrates and Plants in Scats of Leopard Cats (Prionailurus bengalensis) in Southwest China. J. Mammal. 2016, 97, 1054–1064. [Google Scholar] [CrossRef]
- Sanderson, J.; Yin, Y.; Drubgyal, N. Of the only endemic cat species in China: The Chinese mountain cat. CAT News Spec. Issue 2010, 2007, 18–21. Available online: http://www.catsg.org/index.php?id=197 (accessed on 4 October 2022).
- Wei, W.; Zhou, W.; Fan, N. Habitat Selection, Feeding and Caring for The Young of Alpine Weasel. Acta Theriol. Sin. 1994, 14, 184–188. [Google Scholar] [CrossRef]
- Law, C.J. Mustela Sibirica (Carnivora: Mustelidae). Mamm. Species 2018, 50, 109–118. [Google Scholar] [CrossRef] [Green Version]
- Hartová-Nentvichová, M.; Šálek, M.; Červený, J.; Koubek, P. Variation in the Diet of the Red Fox (Vulpes Vulpes) in Mountain Habitats: Effects of Altitude and Season. Mamm. Biol. 2010, 75, 334–340. [Google Scholar] [CrossRef]
- Polis, G.A.R.; Strong, D. Food Web Complexity and Community Dynamics. Am. Nat. 1996, 147, 813–846. [Google Scholar] [CrossRef]
- Letnic, M.; Ritchie, E.G.; Dickman, C.R. Top Predators as Biodiversity Regulators: The Dingo Canis Lupus Dingo as a Case Study. Biol. Rev. 2012, 87, 390–413. [Google Scholar] [CrossRef] [PubMed]
- Brodie, J.F.; Giordano, A. Lack of Trophic Release with Large Mammal Predators and Prey in Borneo. Biol. Conserv. 2013, 163, 58–67. [Google Scholar] [CrossRef]
- Pepper, S.; Barbour, A.; Glass, J. The Management of Wild Deer in Scotland; Scottish Government, Environment and Forestry Directorate: Edinburgh, UK, 2020. Available online: https://www.gov.scot/binaries/content/documents/govscot/publications/independent-report/2020/02/management-wild-deer-scotland/documents/management-wild-deer-scotland-report-deer-working-group/management-wild-deer-scotland-report-deer-working-group/govscot%3Adocument/management-wild-deer-scotland-report-deer-working-group.pdf (accessed on 4 October 2022).
- Ilic, T.; Stojanov, I.; Dimitrijevic, S. Parasitic Infections in Wild Ruminants and Wild Boar. Vet. Glas. 2011, 65, 419–431. [Google Scholar] [CrossRef]
- Yakobson, B.A.; King, R.; Amir, S.; Devers, N.; Sheichat, N.; Rutenberg, D.; Mildenberg, Z.; David, D. Rabies Vaccination Programme for Red Foxes (Vulpes vulpes) and Golden Jackals (Canis aureus) in Israel (1999–2004). Dev. Biol. 2006, 125, 133–140. [Google Scholar]
- Ferreira, A.; Seco, S.; Figueiredo, J.P.; Loureiro, A.; Gomes, A. Scabies in Wildlife Animals, Pets and Humans: A Re-Emerging Zoonosis? Eur. J. Public Health 2021, 31, 130. [Google Scholar] [CrossRef]
- Luo, X.; Hai, R. An Overview of the Epidemiological Characteristics of Marmota Himalayana Plague on Qinghai-Tibet Plateau. Chin. J. Vector Biol. Control 2010, 21, 394–398. Available online: http://www.bmsw.net.cn/CN/Y2010/V21/I4/394 (accessed on 4 October 2022).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Y | Species X | ||
|---|---|---|---|
| Presence | Absence | Total | |
| Presence | A | B | A + B |
| Absence | C | D | C + D |
| Total | A + C | B + D | N |
| Orde | Family | Species |
|---|---|---|
| Carnivora | Canidae | Red Fox (Vulpes vulpes) |
| Dhole (Cuon alpinus) | ||
| Mustelidae | Siberian Weasel (Mustela sibirica) | |
| Mountain Weasel (Mustela altaica) | ||
| Asian Badger (Meles leucurus) | ||
| Stone Marten (Martes foina) | ||
| Felidae | Chinese Mountain Cat (Felis bieti) | |
| Leopard Cat (Prionailurus bengalensis) | ||
| Cetartiodactyla | Cervidae | Sike Deer (Cervus nippon) |
| Red Deer (Cervus elaphus) | ||
| Siberian Roe Deer (Capreolus pygargus) | ||
| Rodentia | Sciuridae | Siberian Chipmunk (Tamias sibiricus) |
| Himalayan Marmot (Marmota himalayana) | ||
| Muridae and Dipodidae | ||
| Lagomorpha | Ochotonidae | Chinese Red Pika (Ochotona erythrotis) |
| Leporidae | Woolly Hare (Lepus oiostolus) | |
| Galliformes | Phasianidae | Chinese Grouse (Tetrastes sewerzowi) |
| Blue Eared Pheasant (Crossoptilon auritum) | ||
| Blood Pheasant (Ithaginis cruentus) | ||
| Chestnut-throated Partridge (Tetraophasis obscurus) | ||
| Common Pheasant (Phasianus colchicus) |
| Species | Shannon–Wiener Index (H′) | Contribution to Network Complexity |
|---|---|---|
| Himalayan marmot * | 1.770 | 15.62% |
| Blue eared pheasant * | 1.757 | 15.51% |
| Asian badger * | 1.584 | 13.98% |
| Red fox * | 1.379 | 12.17% |
| Blood pheasant * | 1.334 | 11.78% |
| Leopard cat * | 1.097 | 9.69% |
| Common pheasant * | 1.087 | 9.59% |
| Chestnut-throated partridge | 0.693 | 6.12% |
| Chinese grouse | 0.628 | 5.54% |
| Siberian roe deer | 0.000 | 0.00% |
| Mountain weasel | 0.000 | 0.00% |
| Siberian weasel | 0.000 | 0.00% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, T.; Li, Q.; Wang, X.; Cui, G.; Man, Z.; Li, W.; Zhao, M. The Ecological Roles of Medium and Small Carnivores in the Terrestrial Animal Community in Liancheng National Nature Reserve, China. Animals 2022, 12, 3518. https://doi.org/10.3390/ani12243518
Su T, Li Q, Wang X, Cui G, Man Z, Li W, Zhao M. The Ecological Roles of Medium and Small Carnivores in the Terrestrial Animal Community in Liancheng National Nature Reserve, China. Animals. 2022; 12(24):3518. https://doi.org/10.3390/ani12243518
Chicago/Turabian StyleSu, Tengwei, Qian Li, Xiaojuan Wang, Guofa Cui, Zihong Man, Wentao Li, and Minyan Zhao. 2022. "The Ecological Roles of Medium and Small Carnivores in the Terrestrial Animal Community in Liancheng National Nature Reserve, China" Animals 12, no. 24: 3518. https://doi.org/10.3390/ani12243518