
Changes in Cat Facial Morphology Are Related to Interaction with Humans

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
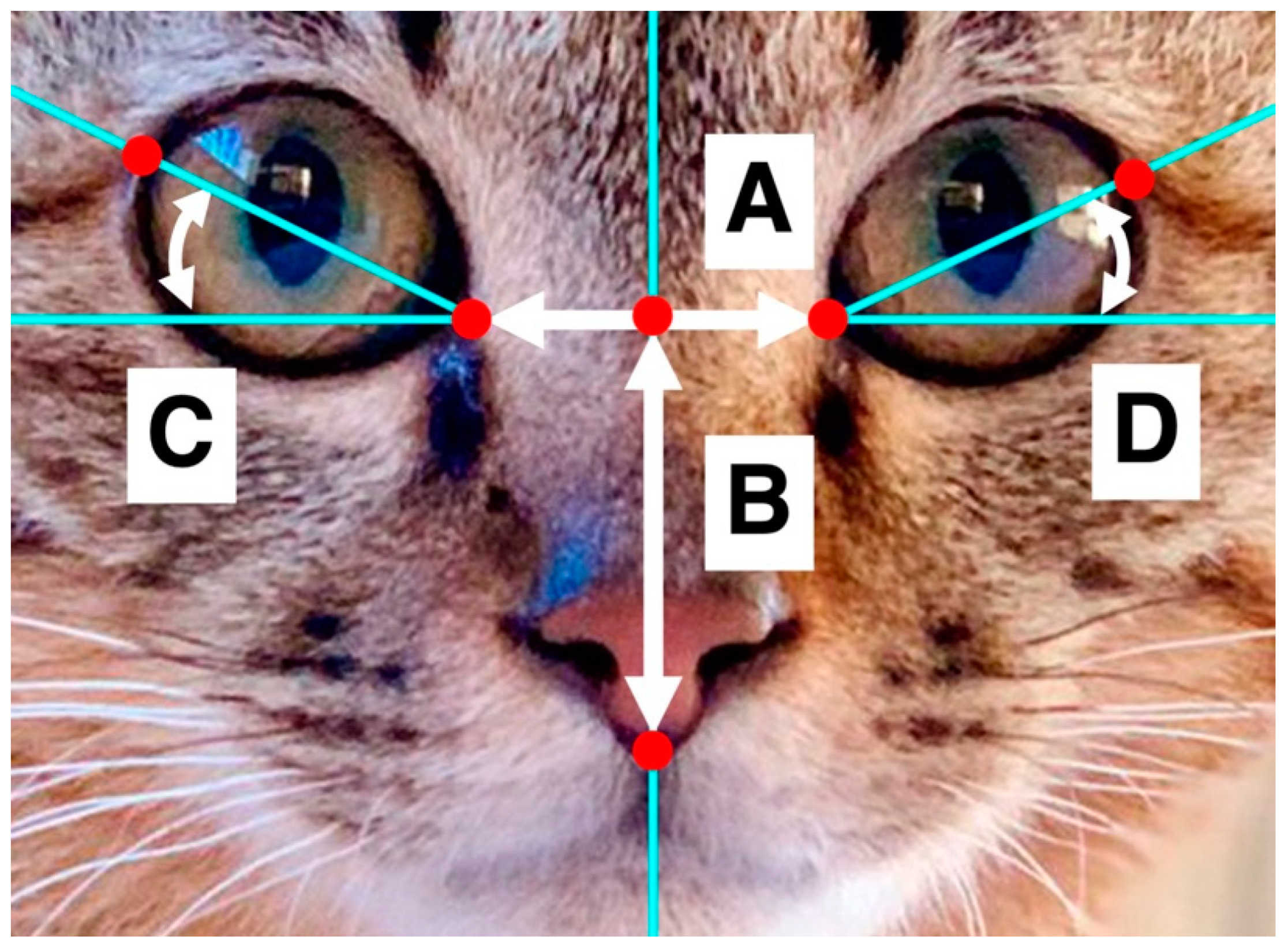
2. Materials and Methods
2.1. Study 1
2.2. Study 2
2.3. Study 3
3. Results
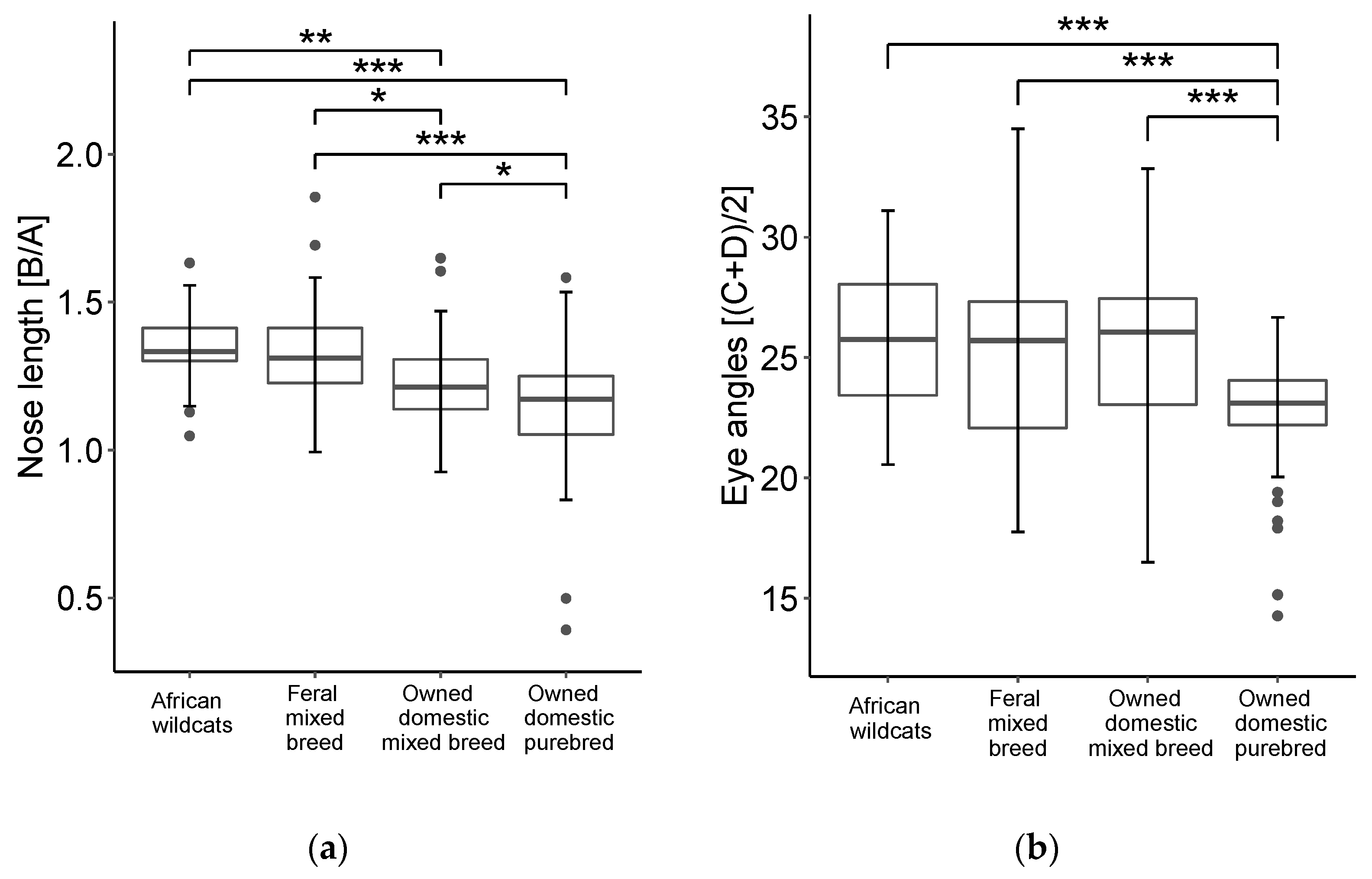
3.1. Study 1
3.2. Study 2
3.3. Study 3
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bruce, V. Recognising Faces; Routledge: Nottingham, UK, 2017; Available online: https://play.google.com/store/books/details?id=5ciRDgAAQBAJ (accessed on 20 October 2022).
- Lorenz, F.W. Fattening cockerels by stilbestrol administration. Poult. Sci. 1943, 22, 190–191. [Google Scholar] [CrossRef]
- Alley, T.R. Head shape and the perception of cuteness. Dev. Psychol. 1981, 17, 650–654. [Google Scholar] [CrossRef]
- Almanza-Sepúlveda, M.L.; Dudin, A.; Wonch, K.E.; Steiner, M.; Feinberg, D.R.; Fleming, A.S.; Hall, G.B. Exploring the morphological and emotional correlates of infant cuteness. Infant Behav. Dev. 2018, 53, 90–100. [Google Scholar] [CrossRef]
- Hildebrandt, K.A.; Fitzgerald, H.E. Facial feature determinants of perceived infant attractiveness. Infant Behav. Dev. 1979, 2, 329–339. [Google Scholar] [CrossRef]
- Sternglanz, S.H.; Gray, J.L.; Murakami, M. Adult preferences for infantile facial features: An ethological approach. Anim. Behav. 1977, 25, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Glocker, M.L.; Langleben, D.D.; Ruparel, K.; Loughead, J.W.; Gur, R.C.; Sachser, N. Baby schema in infant faces induces cuteness perception and motivation for caretaking in adults. Ethology 2009, 115, 257–263. [Google Scholar] [CrossRef] [Green Version]
- Glocker, M.L.; Langleben, D.D.; Ruparel, K.; Loughead, J.W.; Valdez, J.N.; Griffin, M.D.; Sachser, N.; Gur, R.C. Baby schema modulates the brain reward system in nulliparous women. Proc. Natl. Acad. Sci. USA 2009, 106, 9115–9119. [Google Scholar] [CrossRef] [Green Version]
- Kruger, D.J. Non-mammalian infants requiring parental care elicit greater human caregiving reactions than superprecocial infants do. Ethology 2015, 121, 769–774. [Google Scholar] [CrossRef]
- Trut, L.; Oskina, I.; Kharlamova, A. Animal evolution during domestication: The domesticated fox as a model. Bioessays 2009, 31, 349–360. [Google Scholar] [CrossRef] [Green Version]
- Kaminski, J.; Waller, B.M.; Diogo, R.; Hartstone-Rose, A.; Burrows, A.M. Evolution of facial muscle anatomy in dogs. Proc. Natl. Acad. Sci. USA 2019, 116, 14677–14681. [Google Scholar] [CrossRef]
- Waller, B.M.; Peirce, K.; Caeiro, C.C.; Scheider, L.; Burrows, A.M.; McCune, S.; Kaminski, J. Paedomorphic facial expressions give dogs a selective advantage. PLoS ONE 2013, 8, e82686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Senese, V.P.; De Falco, S.; Bornstein, M.H.; Caria, A.; Buffolino, S.; Venuti, P. Human infant faces provoke implicit positive affective responses in parents and non-parents alike. PLoS ONE 2013, 8, e80379. [Google Scholar] [CrossRef] [PubMed]
- Caria, A.; de Falco, S.; Venuti, P.; Lee, S.; Esposito, G.; Rigo, P.; Birbaumer, N.; Bornstein, M.H. Species-specific response to human infant faces in the premotor cortex. NeuroImage 2012, 60, 884–893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Little, A.C. Manipulation of infant-like traits affects perceived cuteness of infant, adult and cat faces. Ethology 2012, 118, 775–782. [Google Scholar] [CrossRef]
- Borgi, M.; Cogliati-Dezza, I.; Brelsford, V.; Meints, K.; Cirulli, F. Baby schema in human and animal faces induces cuteness perception and gaze allocation in children. Front. Psychol. 2014, 5, 411. [Google Scholar] [CrossRef] [Green Version]
- Jack, S.; Carroll, G.A. The effect of baby schema in cats on length of stay in an Irish animal shelter. Animals 2022, 12, 1461. [Google Scholar] [CrossRef]
- Diamond, J. Guns, Germs, and Steel: The Fates of Human Societies; W.W. Norton & Co.: New York, NY, USA, 1997. [Google Scholar]
- Vigne, J.D.; Guilaine, J.; Debue, K.; Haye, L.; Gérard, P. Early taming of the cat in Cyprus. Science 2004, 304, 259. [Google Scholar] [CrossRef] [Green Version]
- Driscoll, C.A.; Macdonald, D.W.; O’Brien, S.J. From wild animals to domestic pets, an evolutionary view of domestication. Proc. Natl. Acad. Sci. USA 2009, 106, 9971–9978. [Google Scholar] [CrossRef] [Green Version]
- Hemmer, H. Man’s strategy in domestication—A synthesis of new research trends. Experientia 1976, 32, 663–666. [Google Scholar] [CrossRef]
- Kruska, D. Mammalian domestication and its effect on brain structure and behavior. Intell. Evol. Biol. 1988, 17, 211–250. [Google Scholar] [CrossRef]
- Ragni, B.; Randi, E. Multivariate analysis of craniometric characters in European wild cat, domestic cat, and African wild cat (genus Felis). Z. Für Säugetierkunde 1986, 51, 243–251. Available online: https://pascal-francis.inist.fr/vibad/index.php?action=getRecordDetail&idt=7886176 (accessed on 20 October 2022).
- Lesch, R.; Kitchener, A.C.; Hantke, G.; Kotrschal, K.; Fitch, W.T. Cranial volume and palate length of cats, Felis spp., under domestication, hybridization and in wild populations. R. Soc. Open Sci. 2022, 9, 210477. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, A.S.; Wrangham, R.W.; Fitch, W.T. The ‘domestication syndrome’ in mammals: A unified explanation based on neural crest cell behavior and genetics. Genetics 2014, 197, 795–808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hare, B. Survival of the friendliest: Homo sapiens Evolved via Selection for Prosociality. Annu. Rev. Psychol. 2017, 68, 155–186. [Google Scholar] [CrossRef]
- Kobayashi, H.; Kohshima, S. Unique morphology of the human eye. Nature 1997, 387, 767–768. [Google Scholar] [CrossRef]
- Komori, M.; Nittono, H. Influence of age-independent facial traits on adult judgments of cuteness and infantility of a child’s face. Procedia Soc. Behav. Sci. 2013, 97, 285–291. [Google Scholar] [CrossRef] [Green Version]
- Kuze, N.; Malim, T.P.; Kohshima, S. Developmental changes in the facial morphology of the Borneo orangutan (Pongo pygmaeus): Possible signals in visual communication. Am. J. Primatol. 2005, 65, 353–376. [Google Scholar] [CrossRef]
- Borgi, M.; Majolo, B. Facial width-to-height ratio relates to dominance style in the genus Macaca. PeerJ 2016, 4, e1775. [Google Scholar] [CrossRef] [Green Version]
- Wilson, V.; Weiss, A.; Lefevre, C.E.; Ochiai, T.; Matsuzawa, T.; Inoue-Murayama, M.; Freeman, H.; Herrelko, E.S.; Altschul, D. Facial width-to-height ratio in chimpanzees: Links to age, sex and personality. Evol. Hum. Behav. Off. J. Hum. Behav. Evol. Soc. 2020, 41, 226–234. [Google Scholar] [CrossRef]
- Geniole, S.N.; Denson, T.F.; Dixson, B.J.; Carré, J.M.; McCormick, C.M. Evidence from meta-analyses of the facial width-to-height ratio as an evolved cue of threat. PLoS ONE 2015, 10, e0132726. [Google Scholar] [CrossRef] [Green Version]
- Malatesta, C.Z.; Fiore, M.J.; Messina, J.J. Affect, personality, and facial expressive characteristics of older people. Psychol. Aging 1987, 2, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Gründl, M.; Klein, S.; Horczakiwskyj, R.; Feser, D.; Jung, M.; Eisenmann-Klein, M.; Prantl, L. The ‘jaguar’s eye’ as a new beauty trend? Age-related effects in judging the attractiveness of the oblique eye axis. Aesthetic Plast. Surg. 2008, 32, 915–919. [Google Scholar] [CrossRef] [PubMed]
- Duffy, D.L.; de Moura, R.T.D.; Serpell, J.A. Development and evaluation of the Fe-BARQ: A new survey instrument for measuring behavior in domestic cats (Felis s. catus). Behav. Process. 2017, 141, 329–341. [Google Scholar] [CrossRef] [PubMed]
- David, H.A. Ranking from unbalanced paired-comparison data. Biometrika 1987, 74, 432–436. [Google Scholar] [CrossRef]
- Kruska, D.C.T. On the evolutionary significance of encephalization in some eutherian mammals: Effects of adaptive radiation, domestication, and feralization. Brain Behav. Evol. 2005, 65, 73–108. [Google Scholar] [CrossRef] [PubMed]
- Derenne, P. Donnees craniometriques sur le chat haret (Felis catus) de l’Archipel de Kerguelen. Mammalia 1972, 36, 459–481. [Google Scholar] [CrossRef]
- Smith, B.P.; Lucas, T.A.; Norris, R.M.; Henneberg, M. Brain size/body weight in the dingo (Canis dingo): Comparisons with domestic and wild canids. Aust. J. Zool. 2017, 65, 292–301. [Google Scholar] [CrossRef]
- Spencer, P.B.S.; Yurchenko, A.A.; David, V.A.; Scott, R.; Koepfli, K.P.; Driscoll, C.; O’Brien, S.J.; Menotti-Raymond, M. The population origins and expansion of feral cats in Australia. J. Hered. 2016, 107, 104–114. [Google Scholar] [CrossRef] [Green Version]
- Nittono, H. The two-layer model of ‘kawaii’: A behavioural science framework for understanding kawaii and cuteness. East Asian J. Popul. Cult. 2016, 2, 79–95. [Google Scholar] [CrossRef]
- Shionosaki, K.; Yamada, F.; Ishikawa, T.; Shibata, S. Feral cat diet and predation on endangered endemic mammals on a biodiversity hot spot (Amami–Ohshima Island, Japan). Wildl. Res. 2015, 42, 343–352. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor | Estimate | Std. Error | t Value | Pr (>t) |
|---|---|---|---|---|
| Sex | −0.01 | 0.01 | −0.42 | 0.67 |
| Years with the owner | −0.00 | 0.00 | −0.50 | 0.62 |
| Sociability with people | −0.00 | 0.01 | −0.37 | 0.71 |
| Stranger-directed aggression | −0.00 | 0.01 | −0.14 | 0.89 |
| Factor | Estimate | Std. Error | t Value | Pr (>t) |
|---|---|---|---|---|
| Sex | −0.48 | 0.32 | −1.49 | 0.14 |
| Years with the owner | −0.08 | 0.04 | −2.06 | 0.04 * |
| Sociability with people | −0.24 | 0.13 | −1.83 | 0.06 |
| Stranger-directed aggression | −0.03 | 0.16 | −0.19 | 0.85 |
| African Wildcats | Feral Mixed Breed | Owned Domestic Mixed Breed | Owned Domestic Purebred | David‘s Score | normDS | Rank | |
|---|---|---|---|---|---|---|---|
| African wildcats | - | 48 | 25 | 44 | −4.6 | 0.33 | 4 |
| Feral mixed breed | 307 | - | 110 | 183 | 0.78 | 1.69 | 2 |
| Owned domestic mixed breed | 330 | 245 | - | 236 | 3.12 | 2.28 | 1 |
| Owned domestic purebreds | 311 | 172 | 119 | - | 0.75 | 1.69 | 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hattori, M.; Saito, A.; Nagasawa, M.; Kikusui, T.; Yamamoto, S. Changes in Cat Facial Morphology Are Related to Interaction with Humans. Animals 2022, 12, 3493. https://doi.org/10.3390/ani12243493
Hattori M, Saito A, Nagasawa M, Kikusui T, Yamamoto S. Changes in Cat Facial Morphology Are Related to Interaction with Humans. Animals. 2022; 12(24):3493. https://doi.org/10.3390/ani12243493
Chicago/Turabian StyleHattori, Madoka, Atsuko Saito, Miho Nagasawa, Takefumi Kikusui, and Shinya Yamamoto. 2022. "Changes in Cat Facial Morphology Are Related to Interaction with Humans" Animals 12, no. 24: 3493. https://doi.org/10.3390/ani12243493