
Chromosome Evolution of the Liolaemus monticola (Liolaemidae) Complex: Chromosomal and Molecular Aspects
 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
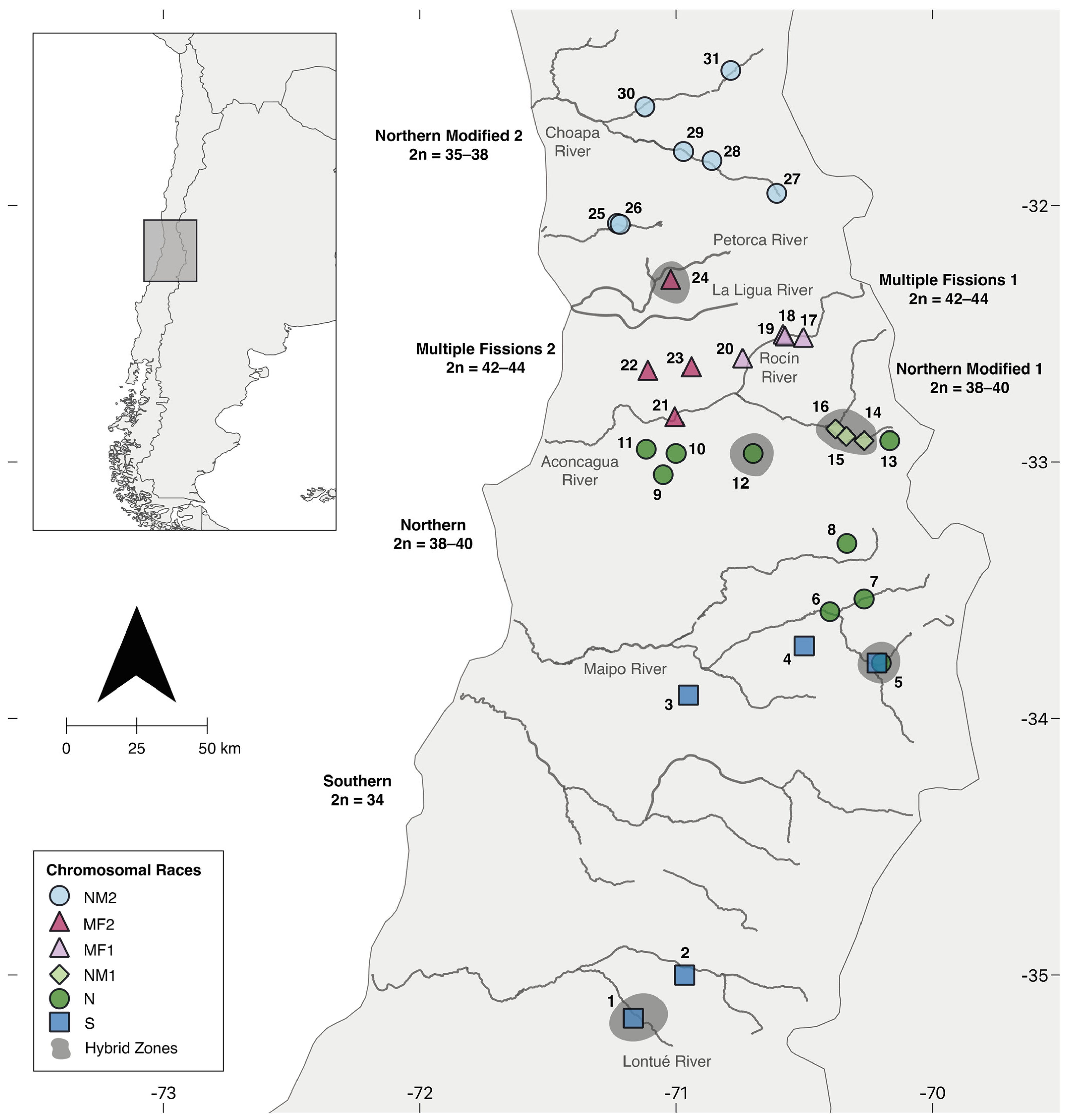
2.1. Sampling and Data Collection
2.2. Cytogenetic Analyses
2.3. Cytochrome b Sequencing and Analysis
3. Results
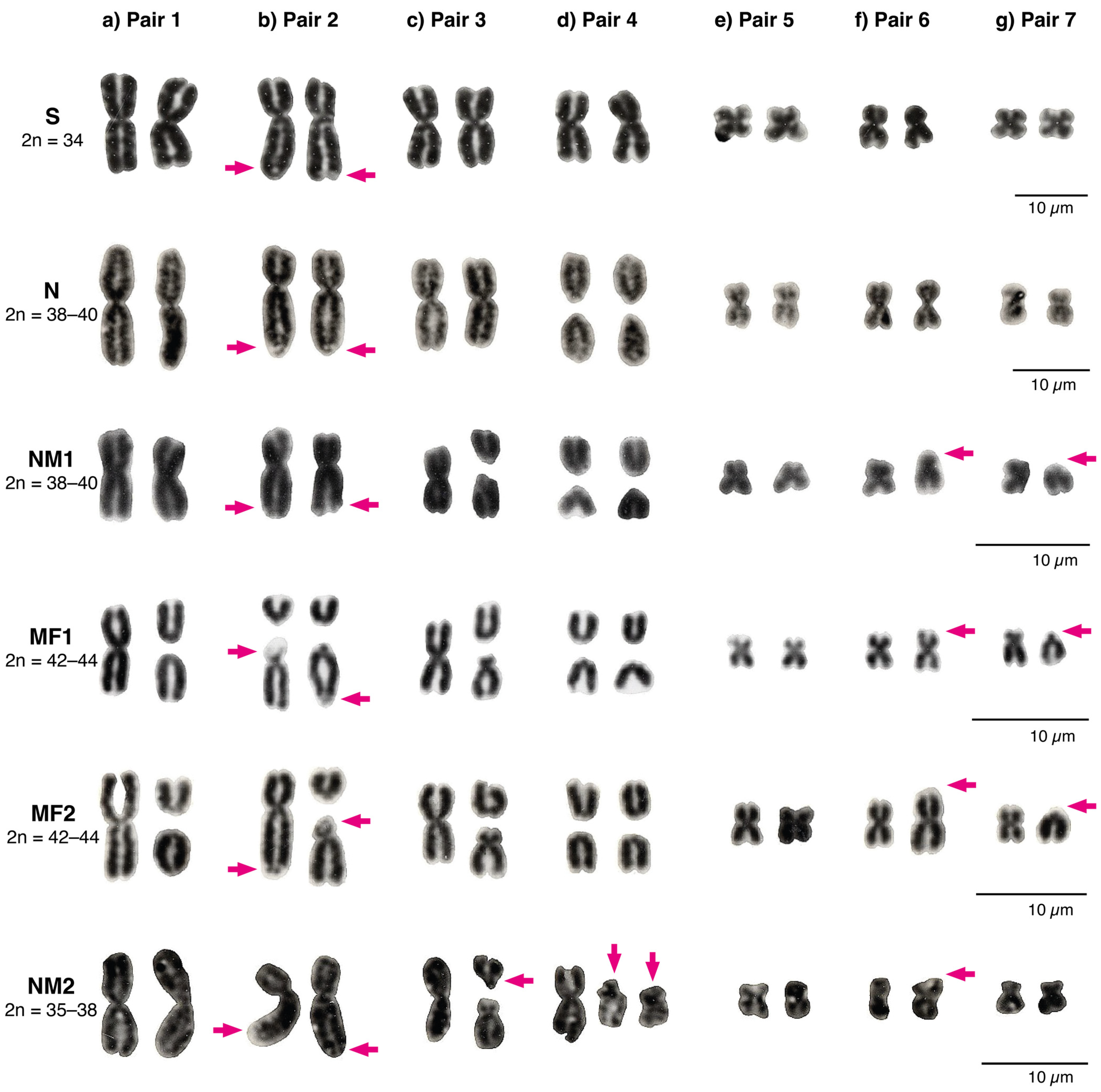
3.1. Patterns of Chromosomal Variation
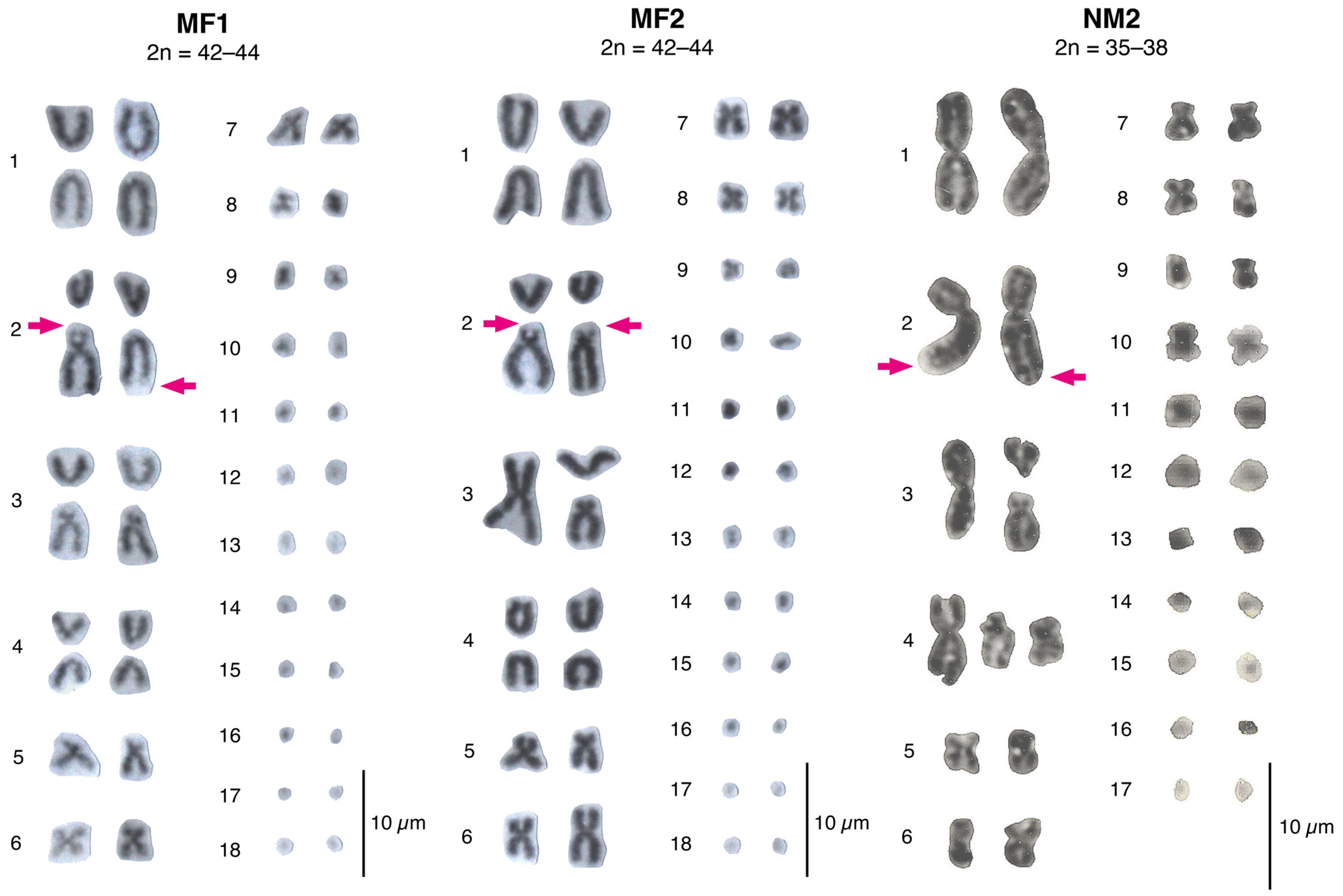
3.1.1. Andean Range between the Aconcagua River and the Petorca River: The Multiple Fission 1 (MF1) Race, 2n = 42–44
3.1.2. Coastal Range between the Aconcagua River and the Petorca River: The Multiple Fissions 2 (MF2) Race, 2n = 42–44
3.1.3. Northward from the Petorca River: The Northern Modified 2 Race, 2n = 35–38
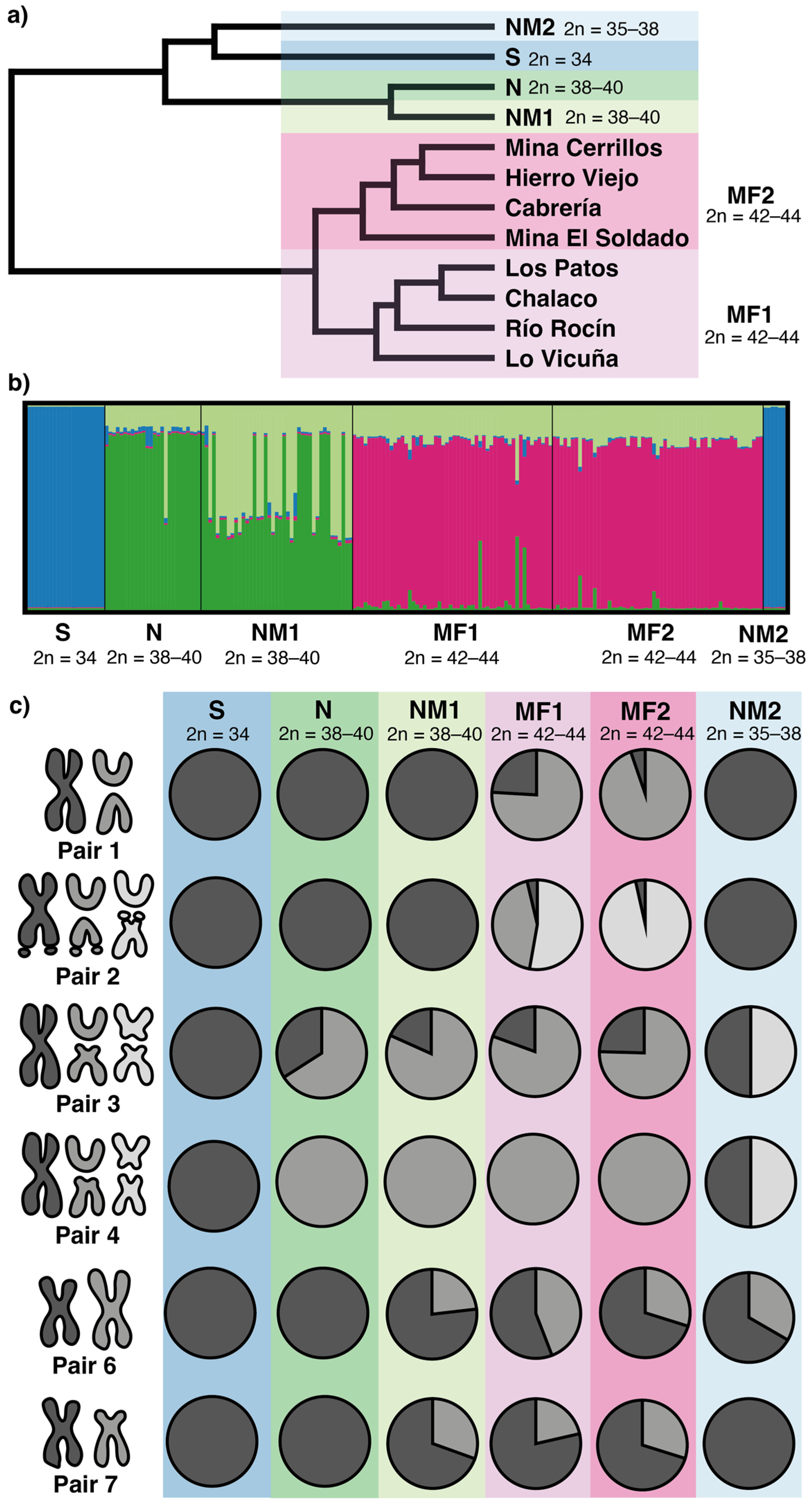
3.2. Population Cytogenetics
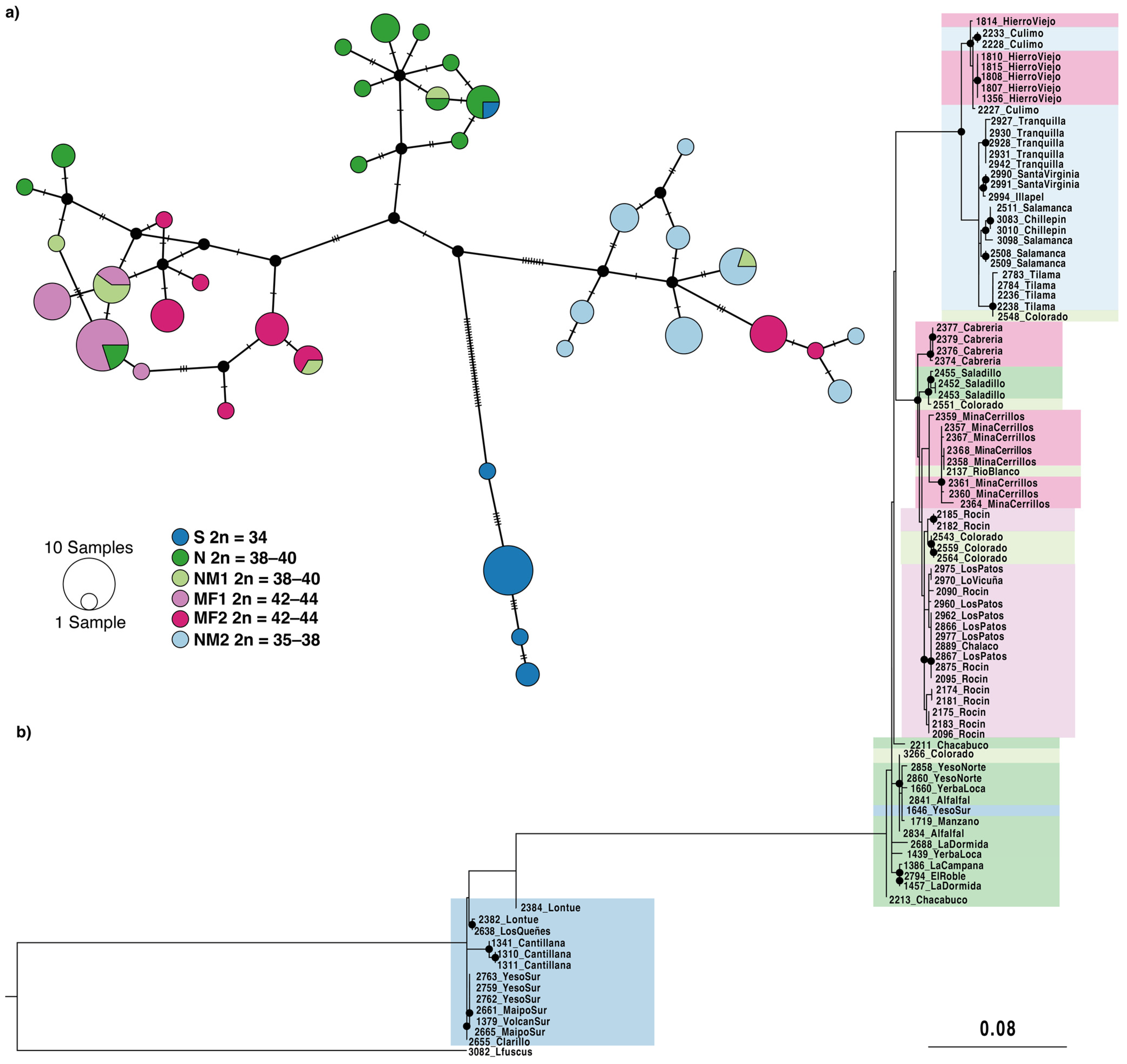
3.3. Mitochondrial cytb Sequence Analyses
4. Discussion
4.1. Patterns of Chromosomal Variation
4.1.1. The Multiple Fissions Races; MF1 and MF2
4.1.2. The Northern Modified 2 Race
4.2. Chromosome Polymorphism
4.3. Riverine Barriers and Gene Flow
4.4. Hybridization between Chromosomal Races
4.5. Model of Chromosome Evolution
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bush, G.L.; Case, S.M.; Wilson, A.C.; Patton, J.L. Rapid speciation and chromosomal evolution in mammals. Proc. Natl. Acad. Sci. USA 1977, 74, 3942–3946. [Google Scholar] [CrossRef] [Green Version]
- White, M.J.D. Chain processes in chromosomal speciation. Syst. Zool. 1978, 27, 285–298. [Google Scholar] [CrossRef]
- King, M. Karyotypic evolution in Gehyra (Gekkonidae: Reptilia) IV. Chromosome change and speciation. Genetica 1984, 64, 101–114. [Google Scholar] [CrossRef]
- King, M. Species Evolution: The Role of Chromosome Change; Cambridge University Press: New York, NY, USA, 1995; pp. X–Y. [Google Scholar]
- Hall, W.P. Comparative population cytogenetics, speciation and evolution of the crevice-using species of Sceloporus (Sauria: Iguanidae). Ph.D. Thesis, Harvard University, Cambridge, MA, USA, 1973. [Google Scholar]
- Paull, D.; Williams, E.E.; Hall, W.P. Lizard karyotypes from Galápagos Islands: Chromosomes in phylogeny and evolution. Breviora 1976, 441, 1–31. [Google Scholar]
- Capanna, E. Robertsonian numerical variation in animal speciation: Mus musculus, an emblematic model. Prog. Clin. Biol. Res. 1982, 96, 155–177. [Google Scholar] [PubMed]
- Sites, J.W., Jr.; Moritz, C. Chromosomal evolution and speciation revisited. Syst. Zool. 1987, 36, 153–174. [Google Scholar] [CrossRef]
- Britton-Davidian, J. How do chromosomal changes fit in? J. Evol. Biol. 2001, 14, 872–873. [Google Scholar] [CrossRef]
- Olmo, E. Rate of chromosome changes and speciation in reptiles. Genetica 2005, 125, 185–203. [Google Scholar] [CrossRef] [PubMed]
- Ayala, F.J.; Coluzzi, M. Chromosome speciation: Humans, Drosophila, and mosquitoes. Proc. Natl. Acad. Sci USA 2005, 102, 6535–6542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coyne, J.A.; Orr, H.A. The evolutionary genetics of speciation. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1998, 35, 287–305. [Google Scholar] [CrossRef] [PubMed]
- Rieseberg, L. Chromosomal rearrangements and speciation. Trends Ecol. Evol. 2001, 16, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Hall, W.P. Chromosome variation, genomics, speciation and evolution in Sceloporus lizards. Cytogenet. Genome Res. 2009, 127, 143–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castiglia, R.; Flores-Villela, O.; Bezerra, A.M.R.; Muñoz, A.; Gornung, E. Pattern of chromosomal changes in ‘beta’ Anolis (Norops group) (Squamata: Polychrotidae) depicted by an ancestral state analysis. Zool. Stud. 2013, 52, 60. [Google Scholar] [CrossRef] [Green Version]
- Gamble, T.; Geneva, A.J.; Glor, R.E.; Zarkower, D. Anolis sex chromosomes are derived from a single ancestral pair. Evolution 2014, 68, 1027–1041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giovannotti, M.; Trifonov, V.A.; Paoletti, A.; Kichigin, I.G.; O’Brien, P.C.M.; Kasai, F.; Giovagnoli, G.; Ng, B.L.; Ruggeri, P.; Nisi Cerioni, P.; et al. New insights into sex chromosome evolution in anole lizards (Reptilia, Dactyloidae). Chromosoma 2017, 126, 245–260. [Google Scholar] [CrossRef]
- Leaché, A.D.; Sites, J.W., Jr. Chromosome evolution and diversification in North American spiny lizards (genus Sceloporus). Cytogenet. Genome Res. 2009, 127, 166–181. [Google Scholar] [CrossRef] [PubMed]
- Leaché, A.D.; Banbury, B.L.; Linkem, C.W.; de Oca, A.N.M. Phylogenomics of a rapid radiation: Is chromosomal evolution linked to increased diversification in north american spiny lizards (Genus Sceloporus)? BMC Evol. Biol. 2016, 16, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Lamborot, M.; Espinoza, A.; Alvarez, E. Karyotypic variation in Chilean lizards of the genus Liolaemus (Iguanidae). Experientia 1979, 35, 593–595. [Google Scholar] [CrossRef]
- Lamborot, M.; Alvarez-Sarret, E. Karyotypic characterization of some Liolaemus lizards in Chile (Iguanidae). Genome 1989, 32, 393–403. [Google Scholar] [CrossRef]
- Aiassa, D.; Martori, R.; Gorla, N. Citogenética de los lagartos del género Liolaemus (Iguania: Liolaemidae) de América del Sur. Cuad. Herpetol. 2005, 18, 23–36. [Google Scholar]
- Lamborot, M. Evolución cromosómica en reptiles. In Herpetología de Chile; Vidal, M.A., Labra, A., Eds.; Science Verlag: Santiago, Chile, 2008; pp. 161–193. [Google Scholar]
- Etheridge, R. Redescription of Ctenoblepharis adspersa Tschudi, 1845, and the taxonomy of Liolaeminae (Reptilia: Squamata: Tropiduridae). Am. Mus. Novit. 1995, 3142, 34. [Google Scholar]
- Esquerré, D.; Brennan, I.G.; Catullo, R.A.; Torres-Pérez, F.; Keogh, J.S. How mountains shape biodiversity: The role of the Andes in biogeography, diversification, and reproductive biology in South America’s most species rich lizard radiation (Squamata: Liolaemidae). Evolution 2019, 73, 214–230. [Google Scholar] [CrossRef] [PubMed]
- Abdala, C.S.; Laspiur, A.; Langstroth, R.P. Las especies del género Liolaemus (Liolaemidae). Cuad. Herpetol. 2021, 35, 193–222. [Google Scholar] [CrossRef]
- Esquerré, D.; Ramírez-Álvarez, D.; Pavón-Vázquez, C.J.; Troncoso-Palacios, J.; Garín, C.F.; Keogh, J.S.; Leaché, A.D. Speciation across mountains: Phylogenomics, species delimitation and taxonomy of the Liolaemus leopardinus clade (Squamata, Liolaemidae). Mol. Phylogenet. Evol. 2019, 139, 106524. [Google Scholar] [CrossRef]
- Araya-Donoso, R.; San Juan, E.; Tamburrino, Í.; Lamborot, M.; Veloso, C.; Véliz, D. Integrating genetics, physiology and morphology to study desert adaptation in a lizard species. J. Anim. Ecol. 2022, 91, 1148–1162. [Google Scholar] [CrossRef]
- Gorman, G.C. The chromosomes of the Reptilia, a cytotaxonomic interpretation. In Cytotaxonomy and Vertebrate Evolution; Chiarelli, A.B., Capanna, E., Eds.; Academic Press: New York, NY, USA, 1972; pp. 349–424. [Google Scholar]
- Espinoza, N.D.; Formas, J.R. Karyological pattern of two Chilean lizard species of the genus Liolaemus (Sauria: Iguanidae). Experientia 1976, 32, 299–301. [Google Scholar] [CrossRef]
- Bertolotto, C.E.V.; Rodrigues, M.T.; Skuk, G.; Yonenaga-Yassuda, Y. Comparative cytogenetic analysis with differential staining in three species of Liolaemus (Squamata, Tropiduridae). Hereditas 1996, 125, 257–264. [Google Scholar] [CrossRef]
- Lamborot, M. Chromosomal evolution and speciation in some Chilean lizards. Evol. Biológica 1993, 7, 133–151. [Google Scholar]
- Lamborot, M.; Eaton, L.; Carrasco, B.A. The Aconcagua River as another barrier to Liolaemus monticola (Sauria: Iguanidae) chromosomal races of central Chile. Rev. Chil. De Hist. Nat. 2003, 76, 23–34. [Google Scholar] [CrossRef] [Green Version]
- Lamborot, M.; Alvarez, E.; Campos, I.; Espinoza, A. Karyotypic characterization of three Chilean subspecies of Liolaemus monticola. J. Hered. 1981, 72, 328–334. [Google Scholar] [CrossRef]
- Lamborot, M. Karyotypic variation among populations of Liolaemus monticola (Tropiduridae) separated by riverine barriers in the Andean Range. Copeia 1991, 4, 1044–1059. [Google Scholar] [CrossRef]
- Lamborot, M.; Alvarez-Sarret, E. Karyotypic variation within and between populations of Liolaemus monticola (Tropiduridae) separated by the Maipo River in the coastal range of central Chile. Herpetologica 1993, 49, 435–449. [Google Scholar]
- Lamborot, M.; Ossa, C.G.; Vásquez, M. Population cytogenetics of the “Northern Mod 1” chromosomal race of Liolaemus monticola Müller & Helmich (Iguanidae) from Central Chile. Gayana 2012, 76, 10–21. [Google Scholar]
- Lamborot, M. A new derived and highly polymorphic chromosomal race of Liolaemus monticola (Iguanidae) from the ‘Norte Chico’ of Chile. Chromosome Res. 1998, 6, 247–254. [Google Scholar] [CrossRef]
- Torres-Pérez, F.; Lamborot, M.; Boric-Bargetto, D.; Hernández, C.E.; Ortiz, J.C.; Palma, R.E. Plylogeography of a mountain lizard species: An ancient fragmentation process mediated by rivervine barriers in the Liolaemus monticola complex (Sauria: Liolaemidae). J. Zool. Syst. Evol. Res. 2007, 45, 72–81. [Google Scholar] [CrossRef]
- Vásquez, M.; Torres-Pérez, F.; Lamborot, M. Genetic variation within and between four chromosomal races of Liolaemus monticola in Chile. Herpetol. J. 2007, 17, 149–160. [Google Scholar]
- Lamborot, M.; Eaton, L.C. Concordance of morphological variation and chromosomal races in Liolaemus monticola (Tropiduridae) separated by riverine barriers in the Andes. J. Zool. Syst. Evol. Res. 1992, 30, 189–200. [Google Scholar] [CrossRef]
- Lamborot, M.; Eaton, L. The Maipo River as a biogeographical barrier to Liolaemus monticola (Tropiduridae) in the mountain ranges of central Chile. J. Zool. Syst. Evol. Res. 1997, 35, 105–111. [Google Scholar] [CrossRef]
- Lamborot, M. Karyotypic polymorphism and evolution within and between the Liolaemus monticola (Iguanidae) “northern 2n = 38–40″ chromosome race populations in Central Chile. Rev. Chil. Hist. Nat. 2001, 71, 121–138. [Google Scholar] [CrossRef] [Green Version]
- Aravena-Muñoz, N. Niveles de Introgresiόn Interracial en Liolaemus monticola (Iguanidae). Master’s Thesis, Universidad de Chile, Santiago, Chile, 2012. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 1 September 2022).
- Rogers, J.S. Measures of genetic similarity and genetics distances. Stud. Genet. 1972, 721, 145–153. [Google Scholar]
- Jombart, T. adegenet: A R package for the multivariate analysis of genetic markers. Bioinformatics 2008, 24, 1403–1405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schliep, K.P. phangorn: Phylogenetic analysis in R. Bioinformatics 2011, 27, 592–593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pritchard, J.K.; Matthew-Stephens, M.M.; Donnelly, P. Inference of population structure using multi-locus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef] [PubMed]
- Earl, D.A.; VonHoldt, B.M. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Goudet, J. Hierfstat, a package for R to compute and test hierarchical F-statistics. Mol. Ecol. Notes 2005, 5, 184–186. [Google Scholar] [CrossRef] [Green Version]
- Paradis, E. pegas: An R package for population genetics with an integrated–modular approach. Bioinformatics 2010, 26, 419–420. [Google Scholar] [CrossRef] [Green Version]
- Aljanabi, S.M.; Martinez, L. Universal and rapid salt-extraction of high quality genomic DNA for PCR-based techniques. Nucleic Acids Res. 1997, 25, 4692–4693. [Google Scholar] [CrossRef]
- Palumbi, S.R. Nucleic acids II: The polymerase chain reaction. In Molecular Systematics, 2nd ed.; Hillis, D.M., Moritz, C., Mable, B.K., Eds.; Sinauer Associates, Inc.: Sunderland, MA, USA, 1996; pp. 205–247. [Google Scholar]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gatewayfor inference of large phylogenetic trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A. FigTree v1.4.2, A Graphical Viewer of Phylogenetic Trees. 2004. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 15 September 2022).
- Leigh, J.W.; Bryant, D. popart: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Kolnicki, R.L. Kinetochore reproduction in animal evolution: Cell biological explanation of karyotypic fission theory. Proc. Natl. Acad. Sci. USA 2000, 97, 9493–9497. [Google Scholar] [CrossRef] [Green Version]
- Dobigny, G.; Britton-Davidian, J.; Robinson, T.J. Chromosomal polymorphism in mammals: An evolutionary perspective. Biol. Rev. 2017, 92, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Porter, C.A.; Sites, J.W., Jr. Evolution of Sceloporus grammicus complex (Sauria: Iguanidae) in central Mexico: Population cytogenetics. Syst. Biol. 1986, 35, 334–358. [Google Scholar] [CrossRef]
- Goyenechea, I.; Mendoza-Quijano, F.; Flores-Villela, O.; Reed, K.M. Extreme chromosomal polytypy in a population of Sceloporus grammicus (Sauria: Phrynosomatidae) at Santuario Mapethé, Hidalgo, México. J. Herpetol. 1996, 30, 39–45. [Google Scholar] [CrossRef]
- Hanski, I.; Gaggiotti, O. (Eds.) Metapopulation biology: Past, present, and future. In Ecology, Genetics and Evolution of Metapopulations; Academic Press: New York, NY, USA, 2004; pp. 3–22. [Google Scholar] [CrossRef]
- Eriksson, J.; Hohmann, G.; Boesch, C.; Vigilant, L. Rivers influence the population genetic structure of bonobos (Pan paniscus). Mol. Ecol. 2004, 13, 3425–3435. [Google Scholar] [CrossRef] [PubMed]
- Araya-Donoso, R.; Torres-Perez, F.; Lamborot, M.; Veliz, D. Hybridization and polyploidy in the weeping lizard Liolaemus chiliensis (Squamata: Liolaemidae). Biol. J. Linn. Soc. 2019, 128, 963–974. [Google Scholar] [CrossRef]
- Sallaberry-Pincheira, N.; Garin, C.F.; González-Acuña, D.; Sallaberry, M.A.; Vianna, J.A. Genetic divergence of Chilean long-tailed snake (Philodryas chamissonis) across latitudes: Conservation threats for different lineages. Divers. Distrib. 2011, 17, 152–162. [Google Scholar] [CrossRef]
- Torres-Pérez, F.; Boric-Bargetto, D.; Rodríguez-Valenzuela, E.; Escobar, C.; Palma, R.E. Molecular phylogenetic analyses reveal the importance of taxon sampling in cryptic diversity: Liolaemus nigroviridis and L. monticola (Liolaeminae) as focal species. Rev. Chil. Hist. Nat. 2017, 90, 5. [Google Scholar] [CrossRef] [Green Version]
- Brüggen, J. Fundamentos de la geología de Chile; Instituto Geográfico Militar: Santiago, Chile, 1950; 382p. [Google Scholar]
- Vuilleumier, B.S. Pleistocene changes in the fauna and flora of South America. Science 1971, 173, 771–780. [Google Scholar] [CrossRef]
- Caviedes, C. Geomorfología del Cuaternario del valle del Aconcagua, Chile Central; Geographischen Institute der Albert-Ludwigs-Universität: Freiburg, Germany, 1972; p. 153. [Google Scholar]
- Heusser, C.J. Late-Pleistocene pollen diagrams from the Province of Llanquihue, southern Chile. Proc. Am. Philos. Soc. 1966, 110, 269–305. [Google Scholar]
- Olave, M.; Avila, L.J.; Sites, J.W., Jr.; Morando, M. Hybridization could be a common phenomenon within the highly diverse lizard genus Liolaemus. J. Evol. Biol. 2018, 31, 893–903. [Google Scholar] [CrossRef]
- Biltueva, L.; Vorobieva, N.; Perelman, P.; Trifonov, V.; Volobouev, V.; Panov, V.; Ilyashenko, V.; Onischenko, S.; O’Brien, P.; Yang, F.; et al. Karyotype evolution of Eulipotyphla (Insectivora): The genome homology of seven Sorex species revealed by comparative chromosome painting and banding data. Cytogenet. Genome Res. 2011, 135, 51–64. [Google Scholar] [CrossRef] [PubMed]
- Torgasheva, A.A.; Basheva, E.A.; Gomez Fernandez, M.J.; Mirol, P.; Borodin, P.M. Chromosomes and speciation in tuco-tuco (Ctenomys, Hystricognathi, Rodentia). Russ. J. Genet. Appl. 2017, 7, 350–357. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Race | Location | Race | Location |
|---|---|---|---|
| Southern, 2n = 34 | 1. Lontué | Northern Modified 1, 2n = 38–40 | 16. Colorado River North |
| 2. Los Queñes | Multiple Fissions 1, 2n = 42–44 | 17. Rocín River | |
| 3. Cantillana | 18. Los Patos | ||
| 4. Clarillo River | 19. Chalaco | ||
| 5. Yeso River South | 20. Lo Vicuña | ||
| Northern, 2n = 38–40 | 5. Yeso River North | Multiple Fissions 2, 2n = 42–44 | 21. Cabrería |
| 6. El Manzano | 22. El Soldado | ||
| 7. El Alfalfal | 23. Mina Cerrillos | ||
| 8. Yerba Loca | 24. Hierro Viejo | ||
| 9. El Roble | Northern Modified 2, 2n = 35–38 | 25. Culimó | |
| 10. La Dormida | 26. Tilama | ||
| 11. La Campana | 27. Tranquilla | ||
| 12. Chacabuco | 28. Chillepín | ||
| 13. Saladillo | 29. Salamanca | ||
| Northern Modified 1, 2n = 38–40 | 14. Blanco River | 30. Illapel | |
| 15. Colorado River South | 31. Santa Virginia |
| S | N | NM1 | MF1 | MF2 | NM2 | |
|---|---|---|---|---|---|---|
| Ar | 1 | 1.142 | 1.412 | 1.749 | 1.544 | 1.429 |
| Ho | 0 | 0.060 | 0.115 | 0.273 | 0.155 | 0.381 |
| He | 0 | 0.064 | 0.157 | 0.294 | 0.199 | 0.210 |
| FIS | - | 0.058 * | 0.273 * | 0.078 | 0.126 | −0.810 * |
| S | N | NM1 | MF1 | MF2 | NM2 | |
|---|---|---|---|---|---|---|
| Pair 1 | 1 | 1 | 1 | 0.113 | 0.675 | 1 |
| Pair 2 | 1 | 1 | 1 | 0.838 | 0.784 | 1 |
| Pair 3 | 1 | 0.844 | 0.089 | 0.971 | 0.081 | 0.014 * |
| Pair 4 | 1 | 1 | 1 | 1 | 1 | 0.014 * |
| Pair 5 | 1 | 1 | 1 | 1 | 1 | 1 |
| Pair 6 | 1 | 1 | 0.014 * | 0.542 | 0.002 * | 0.221 |
| Pair 7 | 1 | 1 | 0.381 | 0.001 * | <0.001 * | 1 |
| S | N | NM1 | MF1 | MF2 | NM2 | |
| S | - | 0.853 * | 0.712 * | 0.680 * | 0.777 * | 0.653 * |
| N | 0.332 * | - | 0.151 * | 0.497 * | 0.645 * | 0.652 * |
| NM1 | 0.355 * | 0.036 * | - | 0.443 * | 0.590 | 0.530 |
| MF1 | 0.422 * | 0.176 * | 0.073 * | - | 0.123 * | 0.552 * |
| MF2 | 0.351 * | 0.078 * | 0.028 * | 0.159 * | - | 0.684 * |
| NM2 | 0.349 * | 0.228 * | 0.194 * | 0.380 * | 0.167 * | - |
| S | N | NM1 | MF1 | MF2 | NM2 | |
|---|---|---|---|---|---|---|
| n | 14 | 20 | 7 | 16 | 19 | 19 |
| K | 8 | 16 | 6 | 7 | 10 | 13 |
| S | 91 | 68 | 56 | 12 | 52 | 38 |
| H | 0.824 | 0.979 | 0.952 | 0.867 | 0.895 | 0.947 |
| π | 0.025 | 0.022 | 0.028 | 0.003 | 0.026 | 0.012 |
| D | −1.949 | −1.071 | −1.086 | −2.403 * | −0.731 | −0.837 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lamborot, M.; Ossa, C.G.; Aravena-Muñoz, N.; Véliz, D.; Araya-Donoso, R. Chromosome Evolution of the Liolaemus monticola (Liolaemidae) Complex: Chromosomal and Molecular Aspects. Animals 2022, 12, 3372. https://doi.org/10.3390/ani12233372
Lamborot M, Ossa CG, Aravena-Muñoz N, Véliz D, Araya-Donoso R. Chromosome Evolution of the Liolaemus monticola (Liolaemidae) Complex: Chromosomal and Molecular Aspects. Animals. 2022; 12(23):3372. https://doi.org/10.3390/ani12233372
Chicago/Turabian StyleLamborot, Madeleine, Carmen Gloria Ossa, Nicolás Aravena-Muñoz, David Véliz, and Raúl Araya-Donoso. 2022. "Chromosome Evolution of the Liolaemus monticola (Liolaemidae) Complex: Chromosomal and Molecular Aspects" Animals 12, no. 23: 3372. https://doi.org/10.3390/ani12233372