
Temperature Incubation Influences Gonadal Gene Expression during Leopard Gecko Development

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. RNAs
2.3. Expression Analysis of Genes Involved in Gonadal Sex Differentiation
2.4. DDRT-PCR
2.5. Confirmation of Differential Gene Expression: Real-Time PCR
3. Results
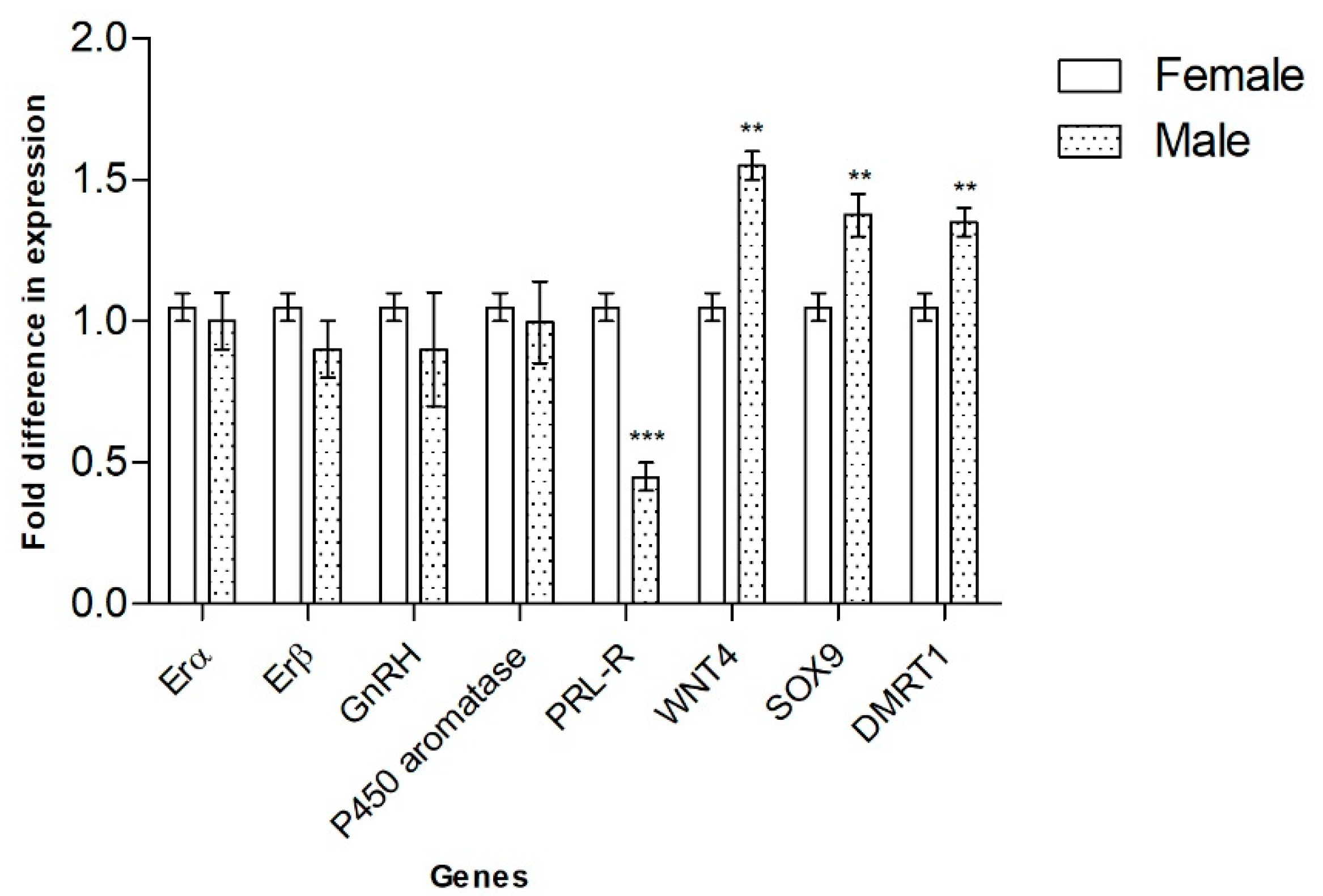
3.1. Genes Involved in Gonadal Sex Differentiation
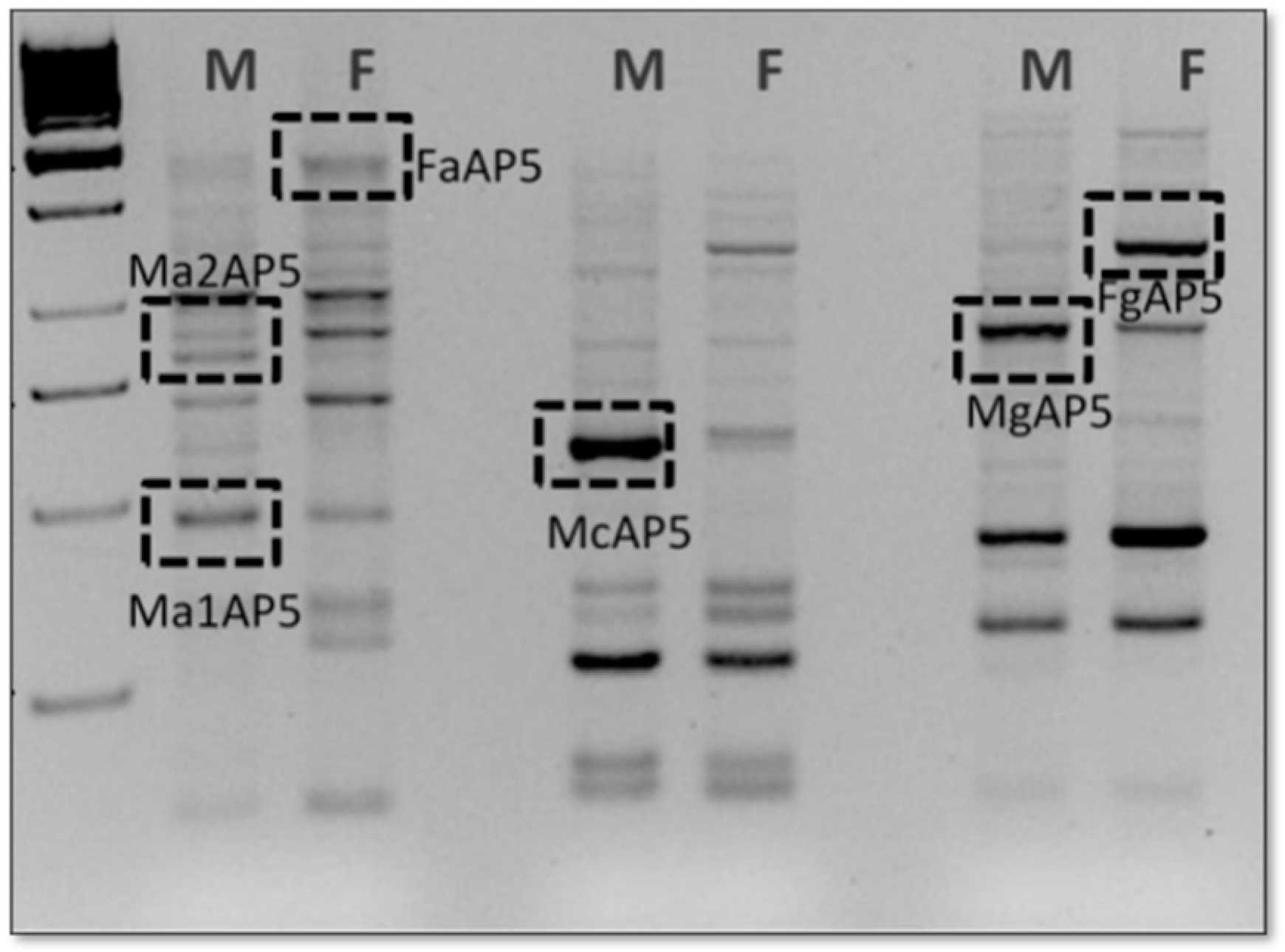
3.2. Identification and Expression Analysis of Seven New Transcripts by DDRT-PCR
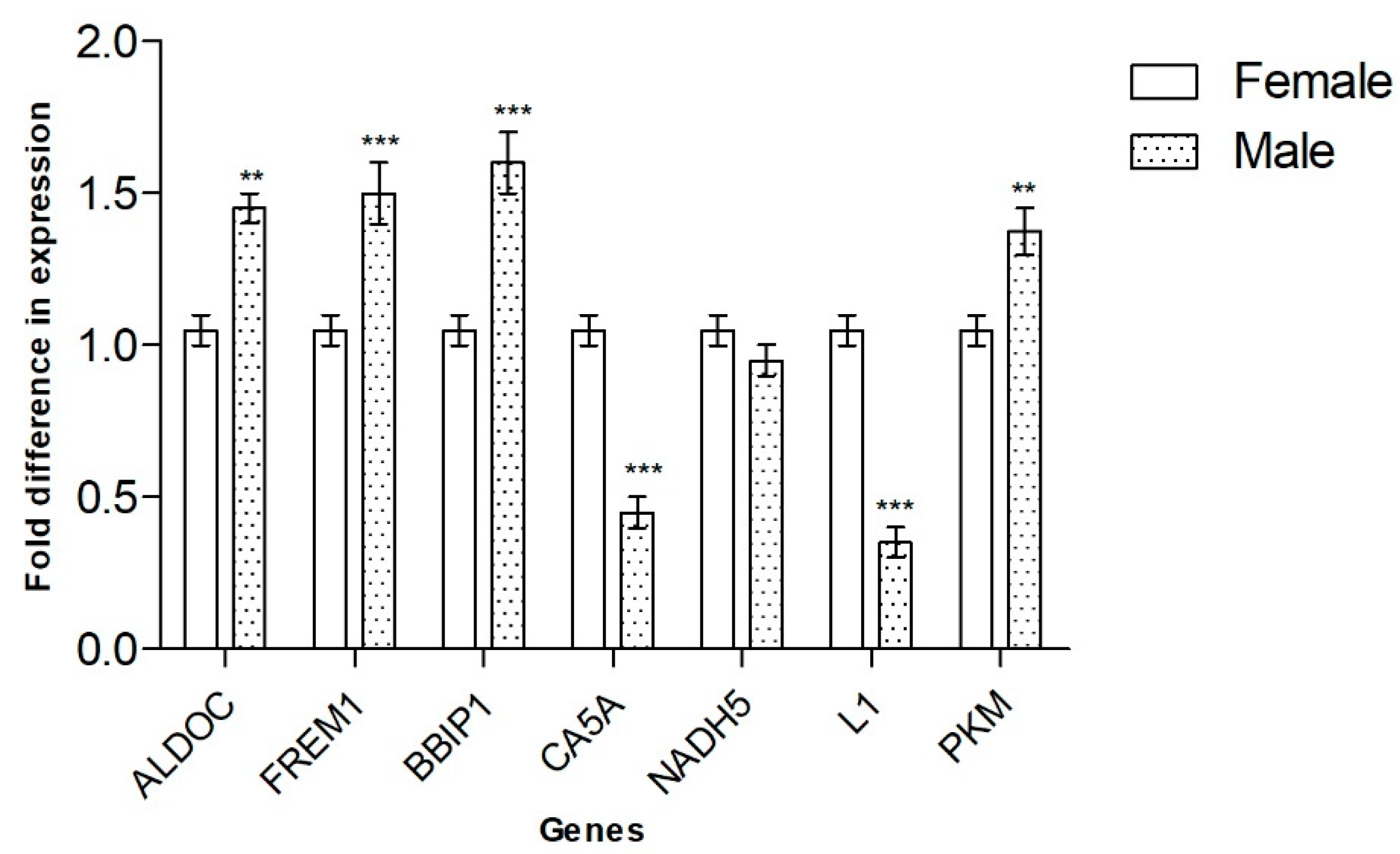
3.3. qRT-PCR Expression Analysis of Genes Identified by DDRT-PCR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Quinn, A.; Koopman, P. The Molecular Genetics of Sex Determination and Sex Reversal in Mammals. Semin. Reprod. Med. 2012, 30, 351–363. [Google Scholar] [CrossRef] [PubMed]
- Mullon, C.; Pomiankowski, A.; Reuter, M. Molecular evolution of Drosophila Sex-lethal and related sex determining genes. BMC Evol. Biol. 2012, 12, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uguz, C.; Iscan, M.; Togan, I. Developmental genetics and physiology of sex differentiation in vertabrates. Environ. Toxicol. Pharmacol. 2003, 14, 9–16. [Google Scholar] [CrossRef]
- Barrionuevo, F.J.; Burgos, M.; Scherer, G.; Jiménez, R. Genes promoting and disturbing testis development. Histol. Histopathol. 2012, 27, 1361–1383. [Google Scholar] [CrossRef]
- Ciofi, C.; Swingland, I.R. Environmental sex determination in reptiles. Appl. Anim. Behav. Sci. 1997, 51, 251–265. [Google Scholar] [CrossRef]
- Janzen, F.J.; Paukstis, G.L. A preliminary test of the adaptive significance of environmental sex determination in reptiles. Evolution 1991, 45, 435–440. [Google Scholar] [CrossRef]
- Miller, D.; Summers, J.; Silber, S. Environmental versus genetic sex determination: A possible factor in dinosaur extinction? Fertil. Steril. 2004, 81, 954–964. [Google Scholar] [CrossRef]
- Girondot, M.; Ben Hassine, S.; Sellos, C.; Godfrey, M.; Guillon, J.-M. Modeling Thermal Influence on Animal Growth and Sex Determination in Reptiles: Being Closer to the Target Gives New Views. Sex. Dev. 2010, 4, 29–38. [Google Scholar] [CrossRef]
- Modi, W.S.; Crews, D. Sex chromosomes and sex determination in reptiles: Commentary. Curr. Opin. Genet. Dev. 2005, 15, 660–665. [Google Scholar] [CrossRef]
- Valenzuela, N. Multivariate Expression Analysis of the Gene Network Underlying Sexual Development in Turtle Embryos with Temperature-Dependent and Genotypic Sex Determination. Sex. Dev. 2010, 4, 39–49. [Google Scholar] [CrossRef]
- Du, W.-G.; Shine, R.; Ma, L.; Sun, B.-J. Adaptive responses of the embryos of birds and reptiles to spatial and temporal variations in nest temperatures. Proc. R. Soc. Boil. Sci. 2019, 286, 20192078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piprek, R.P. Molecular Mechanisms Underlying Female Sex Determination—Antagonism Between Female and Male Pathway. Folia Biol. 2009, 57, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Yildirim, E.; Aksoy, S.; Onel, T.; Yaba, A. Gonadal development and sex determination in mouse. Reprod. Biol. 2020, 20, 115–126. [Google Scholar] [CrossRef] [PubMed]
- Endo, D.; Kanaho, Y.-I.; Park, M.K. Expression of sex steroid hormone-related genes in the embryo of the leopard gecko. Gen. Comp. Endocrinol. 2008, 155, 70–78. [Google Scholar] [CrossRef]
- Valleley, E.M.; Cartwright, E.J.; Croft, N.J.; Markham, A.F.; Coletta, P.L. Characterisation and expression ofSox9 in the Leopard gecko, Eublepharis macularius. J. Exp. Zool. 2001, 291, 85–91. [Google Scholar] [CrossRef]
- Pallotta, M.M.; Turano, M.; Ronca, R.; Mezzasalma, M.; Petraccioli, A.; Odierna, G.; Capriglione, T. Brain Gene Expression is Influenced by Incubation Temperature During Leopard Gecko (Eublepharis macularius) Development. J. Exp. Zool. Mol. Dev. Evol. 2017, 328, 360–370. [Google Scholar] [CrossRef]
- Wise, P.A.; Vickaryous, M.K.; Russell, A.P. An Embryonic Staging Table for in ovo Development of Eublepharis macularius, the Leopard Gecko. Anat. Rec. 2009, 292, 1198–1212. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Maldonado, L.T.; Piedra, A.L.; Mendoza, N.M.; Valencia, A.M.; Martínez, A.M.; Larios, H.M. Expression profiles of Dax1, Dmrt1, and Sox9 during temperature sex determination in gonads of the sea turtle Lepidochelys olivacea. Gen. Comp. Endocrinol. 2002, 129, 20–26. [Google Scholar] [CrossRef]
- Shoemaker, C.M.; Crews, D. Analyzing the coordinated gene network underlying temperature-dependent sex determination in reptiles. Semin. Cell Dev. Biol. 2009, 20, 293–303. [Google Scholar] [CrossRef]
- Shoemaker, C.M.; Queen, J.; Crews, D. Response of Candidate Sex-Determining Genes to Changes in Temperature Reveals Their Involvement in the Molecular Network Underlying Temperature-Dependent Sex Determination. Mol. Endocrinol. 2007, 21, 2750–2763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Western, P.S.; Harry, J.L.; Graves, J.A.; Sinclair, A.H. Temperature-dependent sex determination in the American alligator: Expression of SF1, WT1 and DAX1 during gonadogenesis. Gene 1999, 241, 223–232. [Google Scholar] [CrossRef]
- Valenzuela, N. Evolution of the gene network underlying gonadogenesis in turtles with temperature-dependent and genotypic sex determination. Integr. Comp. Biol. 2008, 48, 476–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capriglione, T.; Vaccaro, M.; Morescalchi, M.; Tammaro, S.; De Iorio, S. Differential DMRT1 Expression in the Gonads of Podarcis sicula (Reptilia: Lacertidae). Sex. Dev. 2010, 4, 104–109. [Google Scholar] [CrossRef]
- Ikeda, Y.; Noguchi, T. Allosteric Regulation of Pyruvate Kinase M2 Isozyme Involves a Cysteine Residue in the Intersubunit Contact. J. Biol. Chem. 1998, 273, 12227–12233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsutsumi, H.; Tani, K.; Fujii, H.; Miwa, S. Expression of L- and M-type pyruvate kinase in human tissues. Genomics 1988, 2, 86–89. [Google Scholar] [CrossRef]
- Yang, W.; Xia, Y.; Ji, H.; Zheng, Y.; Liang, J.; Huang, W.; Gao, X.; Aldape, K.; Lu, Z. Nuclear PKM2 regulates β-catenin transactivation upon EGFR activation. Nature 2011, 480, 118–122. [Google Scholar] [CrossRef] [Green Version]
- Vrontou, S.; Petrou, P.; I Meyer, B.; Galanopoulos, V.K.; Imai, K.; Yanagi, M.; Chowdhury, K.; Scambler, P.; Chalepakis, G. Fras1 deficiency results in cryptophthalmos, renal agenesis and blebbed phenotype in mice. Nat. Genet. 2003, 34, 209–214. [Google Scholar] [CrossRef]
- Loktev, A.V.; Zhang, Q.; Beck, J.S.; Searby, C.C.; Scheetz, T.E.; Bazan, J.F.; Slusarski, D.C.; Sheffield, V.C.; Jackson, P.K.; Nachury, M.V. A BBSome Subunit Links Ciliogenesis, Microtubule Stability, and Acetylation. Dev. Cell 2008, 15, 854–865. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Nishimura, D.; Vogel, T.; Shao, J.; Swiderski, R.; Yin, T.; Searby, C.; Carter, C.C.; Kim, G.; Bugge, K.; et al. BBS7 is required for BBSome formation and its absence in mice results in Bardet-Biedl syndrome phenotypes and selective abnormalities in membrane protein trafficking. J. Cell Sci. 2013, 126, 2372–2380. [Google Scholar] [CrossRef]
- Juszczak, G.R.; Stankiewicz, A.M. Glucocorticoids, genes and brain function. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2018, 82, 136–168. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Montiel, W.; Collí-Dula, R.C.; Ramón-Ugalde, J.P.; Martínez-Núñez, M.A.; Zamora-Bustillos, R. RNA-seq Transcriptome Analysis in Ovarian Tissue of Pelibuey Breed to Explore the Regulation of Prolificacy. Genes 2019, 10, 358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pokharel, K.; Peippo, J.; Weldenegodguad, M.; Honkatukia, M.; Li, M.-H.; Kantanen, J. Gene Expression Profiling of Corpus luteum Reveals Important Insights about Early Pregnancy in Domestic Sheep. Genes 2020, 11, 415. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Gene | Oligo Forward Sequence (5′-3′) | Oligo Reverse Sequence (5′-3′) |
|---|---|---|
| Erα | CACCCTGGAAAGCTGTTGTT | TTCGGAATCGAGTAGCAGTG |
| Erβ | ATCCCGGCAAGCTAATCTTT | CAGCTCTCGAAACCTTGAAGT |
| GnRH | GTCTTGCTGGCCTCTCCTC | GTGGTCTCCTGCCAGTGTTC |
| P450 aromatase | TGAACACCCTCAGTGTGGAA | TCAGVTTTGGCATGTCTTCA |
| PRL | AAGGCCATGGAGATTGAGG | GGAGGCCTGACCAAGTAGAA |
| PRL-R | ATGGAGGTCTCCCCACTAAT | AACAGGAATTGGGTCCTCCT |
| WNT4 | CTGCAACAAGACCTCCAAGG | AGCAGCACCGTGGAATTG |
| SOX9 | GGGCAAGCTTTGGAGGTTAC | TGGGCTGGTACTTATAGTCTGGA |
| DMRT1 | GCAGGGGATCCTACCAAAGT | AGAAGGCAGCAAGCTCAAGA |
| ALDOC | TACCATGGTGTTGTGCAAGC | CTTCACGCTGCATTTTCTCA |
| FREM1 | GGAATGTCAACCAAGATGTGG | CAGGGGAGATCAGAACCACT |
| BBIP1 | CGTGAGCTGTAGCTTTGCAG | CTGCCTTACCCACAGCACTT |
| CA5A | TTGCAAAGTTATGGGGAGGA | TCAAGCAGGGTTTATTTCTCATC |
| NADH5 | GCTACAGGTAAATCCGCTCAA | AGTAGGGCAGAAACGGGAGT |
| L1 non-LTR retrotransposons | ATCATCGTGGGCCTCTTTGC | AGCAGCACCGTGGAATTG |
| PKM | AGAGCTGCTTGTACGCCTGT | CCAGATTTCCAAAGGACAGTG |
| Primers | Sequence |
|---|---|
| 3′ oligo (dT) H-T11G | AAGCTTTTTTTTTTTG |
| 3′ oligo (dT) H-T11A | AAGCTTTTTTTTTTTA |
| 3′ oligo (dT) H-T11C | AAGCTTTTTTTTTTTC |
| H-AP1 | AAGCTTGATTGCC |
| H-AP2 | AAGCTTCGACTGT |
| H-AP3 | AAGCTTTGGTCAG |
| H-AP4 | AAGCTTCTCAACG |
| H-AP5 | AAGCTTAGTAGGC |
| H-AP6 | AAGCTTGCACCAT |
| H-AP7 | AAGCTTAACGAGG |
| H-AP8 | AAGCTTTTACCGC |
| Clone | E-Value | Length (BP) | Identity (%) | Results |
|---|---|---|---|---|
| McAP2 | e−121 | 450 | 98% | Gekko japonicus, aldolase fructose-bisphosphate C (ALDOC) |
| Ma1AP5 | 2e−41 | 528 | 83% | Anolis carolinensis, pyruvate kinase muscle isozyme-like (PKM) |
| Ma2AP5 | 2e−60 | 710 | 95% | Anolis carolinensis, FRAS1 related extracellular matrix 1 (FREM1) |
| FaAP5 | 4e−148 | 1011 | 73% | Hemitheconyx caudicinctus, NADH dehydrogenase subunit 5 (mitochondrion) (NADH-CoQ reduttasii) |
| McAP5 | 1e−144 | 592 | 83% | Anolis carolinensis, BBSome-interacting protein 1-like (BBIP1) |
| Fg1AP6 | 9e−15 | 137 | 77% | Sphaerodactylus townsendi, carbonic anhydrase 5A (CA5A) |
| FcAP7 | 1e−14 | 202 | 76% | Aanolis carolinensis, L1 non-LTR retrotransposons |
| Gene | Identification | Embryo Gonads Stage 30 |
|---|---|---|
| Erα | qRT-PCR | No differential expression |
| Erβ | qRT-PCR | No differential expression |
| GnRH | qRT-PCR | No differential expression |
| P450 aromatase | qRT-PCR | No differential expression |
| PRL | qRT-PCR | Not detected |
| PRL-R | qRT-PCR | FPT |
| WNT4 | qRT-PCR | MPT |
| SOX9 | qRT-PCR | MPT |
| DMRT1 | qRT-PCR | MPT |
| ALDOC | DDRT-PCR | MPT |
| FREM1 | DDRT-PCR | MPT |
| BBIP1 | DDRT-PCR | MPT |
| CA5A | DDRT-PCR | FPT |
| NADH5 | DDRT-PCR | No differential expression |
| L1 non-LTR retrotransposons | DDRT-PCR | FPT |
| PKM | DDRT-PCR | MPT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pallotta, M.M.; Fogliano, C.; Carotenuto, R. Temperature Incubation Influences Gonadal Gene Expression during Leopard Gecko Development. Animals 2022, 12, 3186. https://doi.org/10.3390/ani12223186
Pallotta MM, Fogliano C, Carotenuto R. Temperature Incubation Influences Gonadal Gene Expression during Leopard Gecko Development. Animals. 2022; 12(22):3186. https://doi.org/10.3390/ani12223186
Chicago/Turabian StylePallotta, Maria Michela, Chiara Fogliano, and Rosa Carotenuto. 2022. "Temperature Incubation Influences Gonadal Gene Expression during Leopard Gecko Development" Animals 12, no. 22: 3186. https://doi.org/10.3390/ani12223186