
The Interplay between Central and Peripheral Systems in Feed Intake Regulation in European Seabass (Dicentrarchus labrax) Juveniles

Abstract
:Simple Summary
Abstract
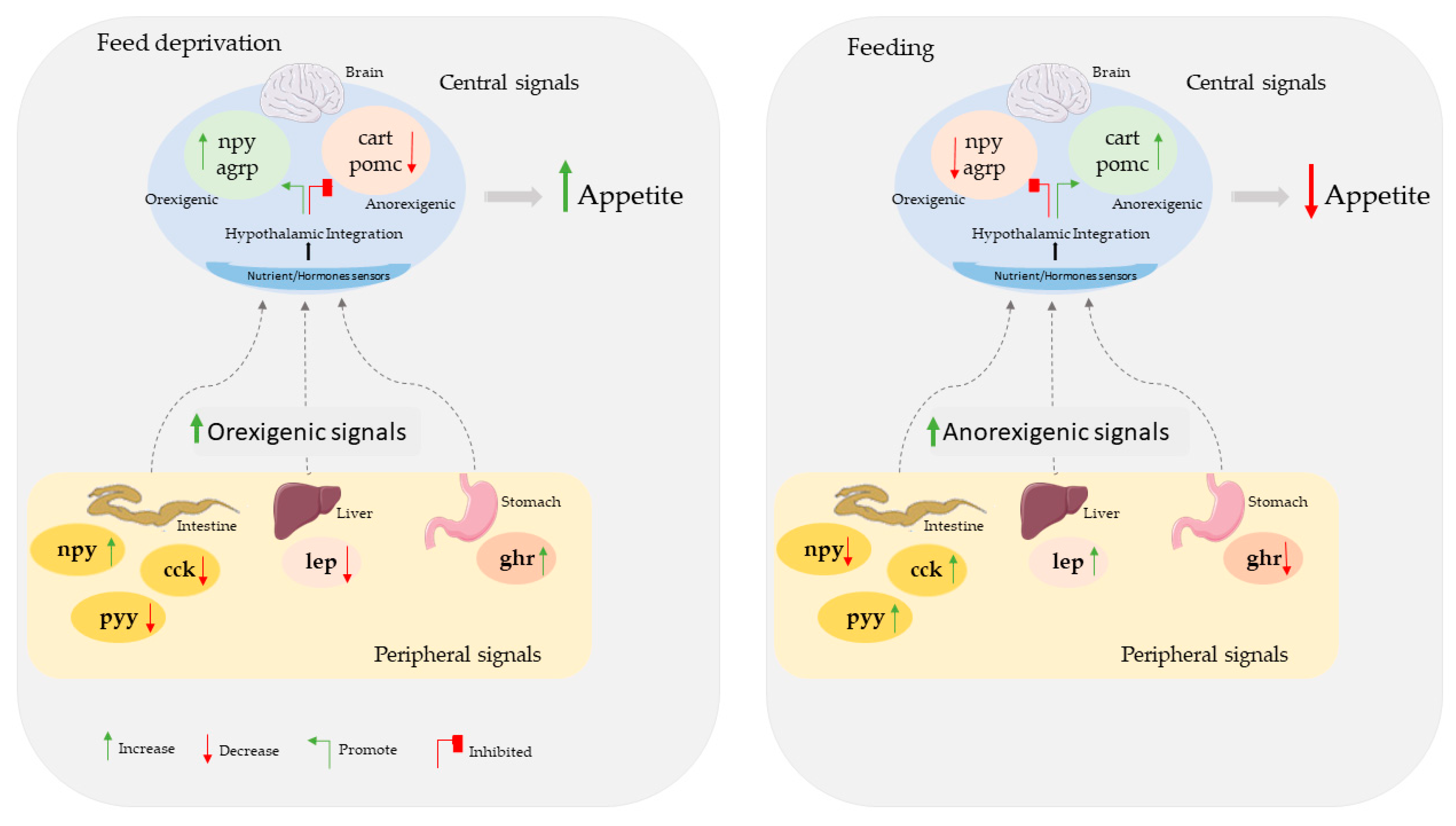
1. Introduction
2. Materials and Methods
2.1. Experimental Conditions
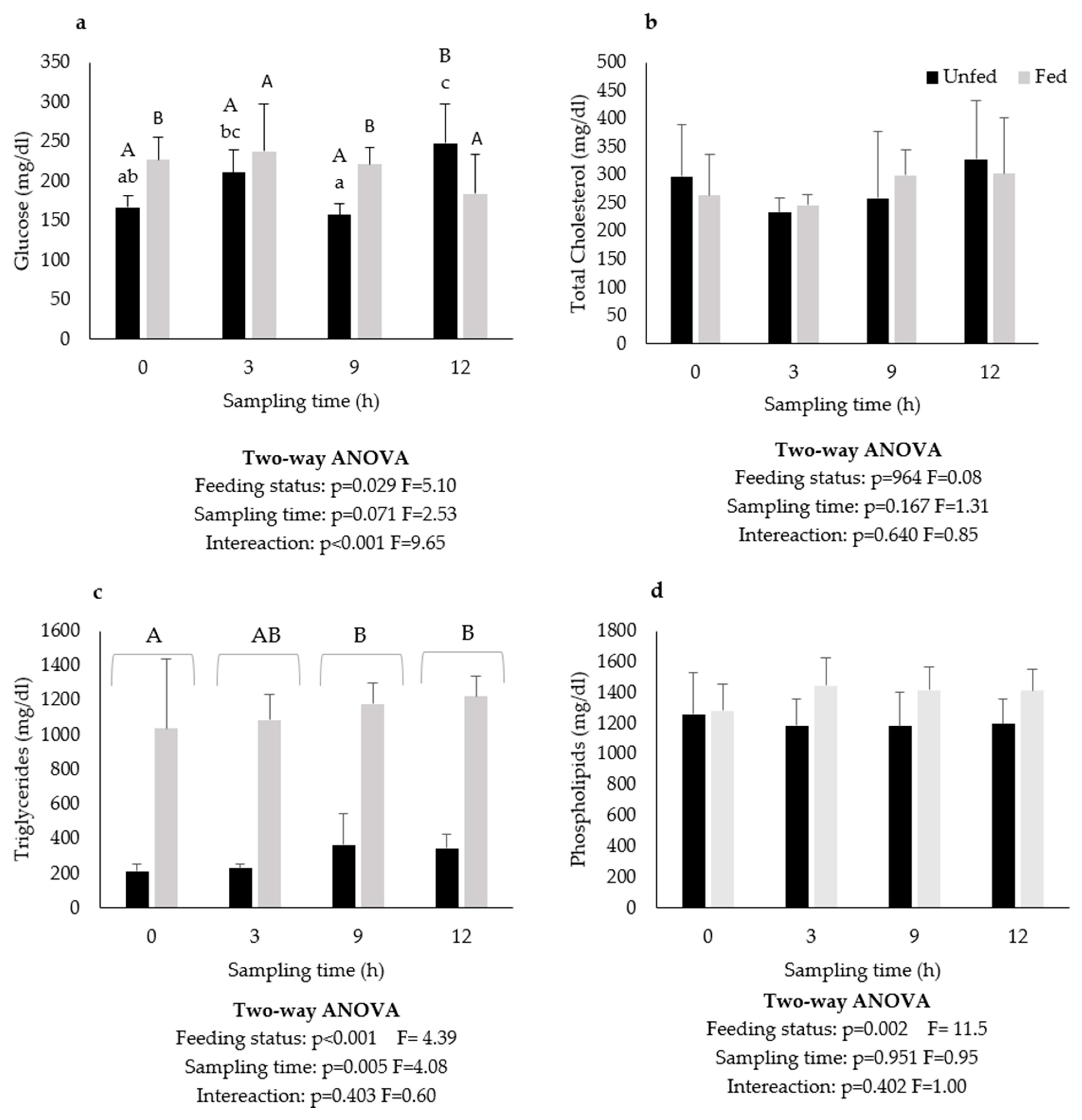
2.2. Plasma Metabolites
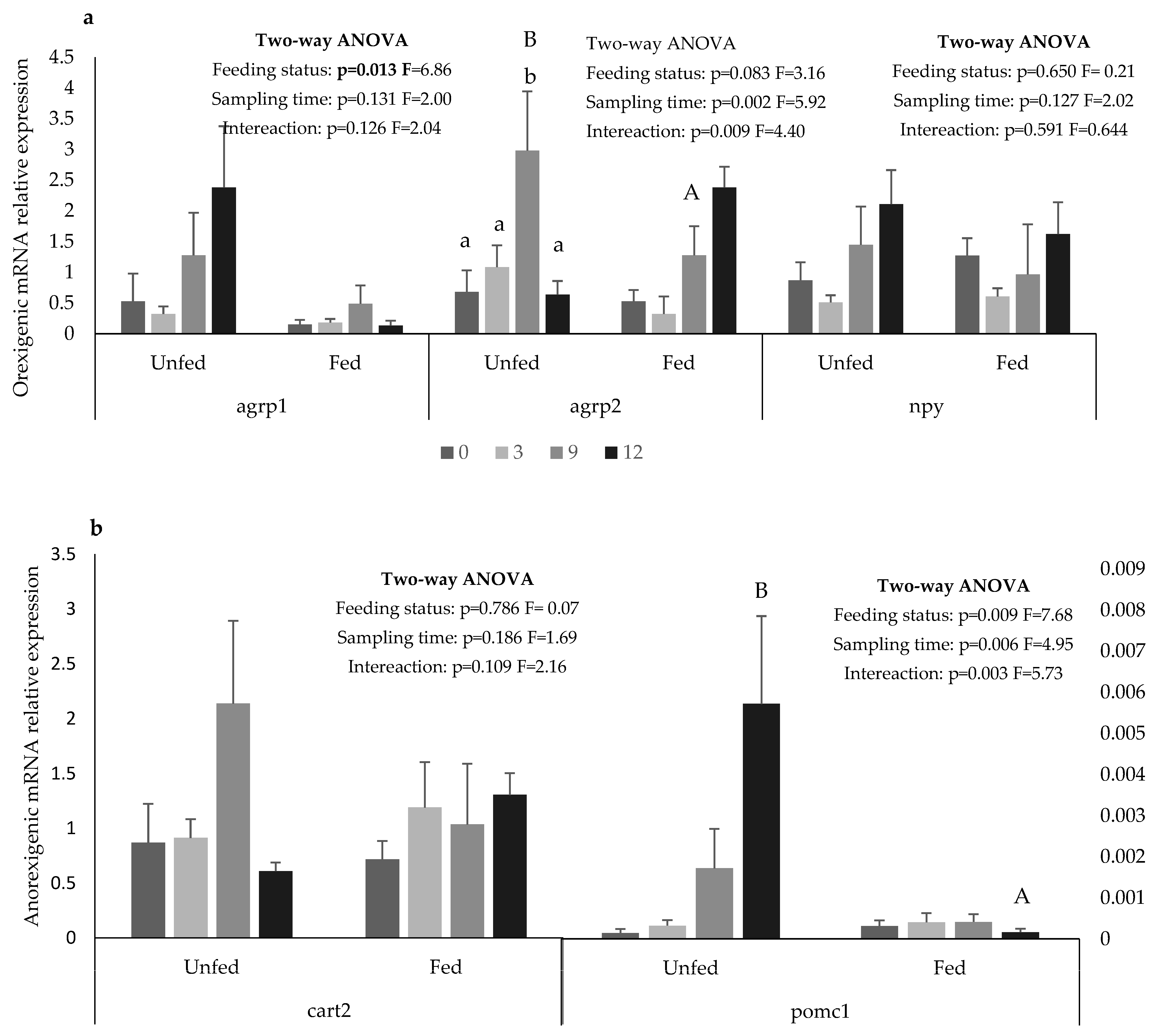
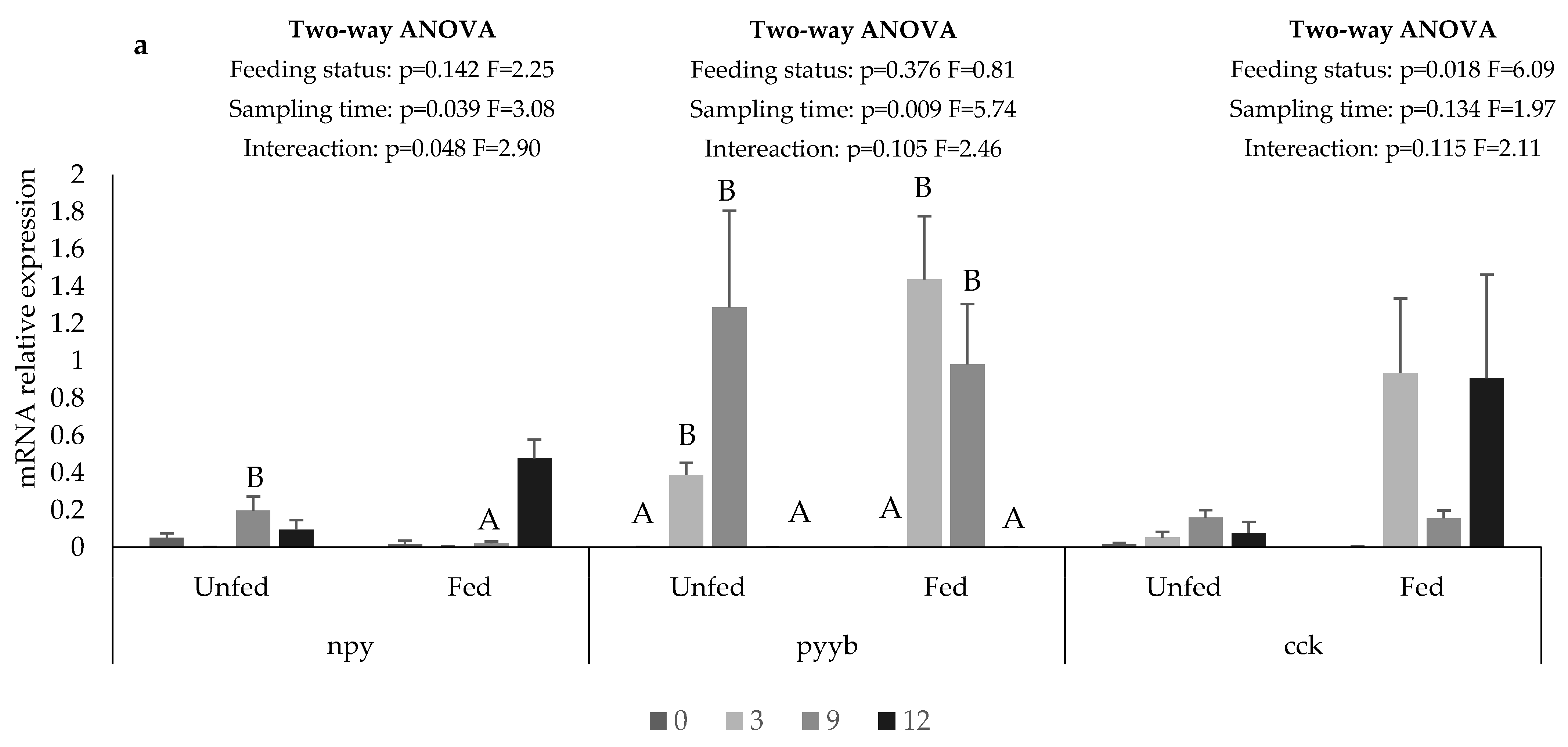
2.3. Gene Expression
2.4. Statistical Analysis
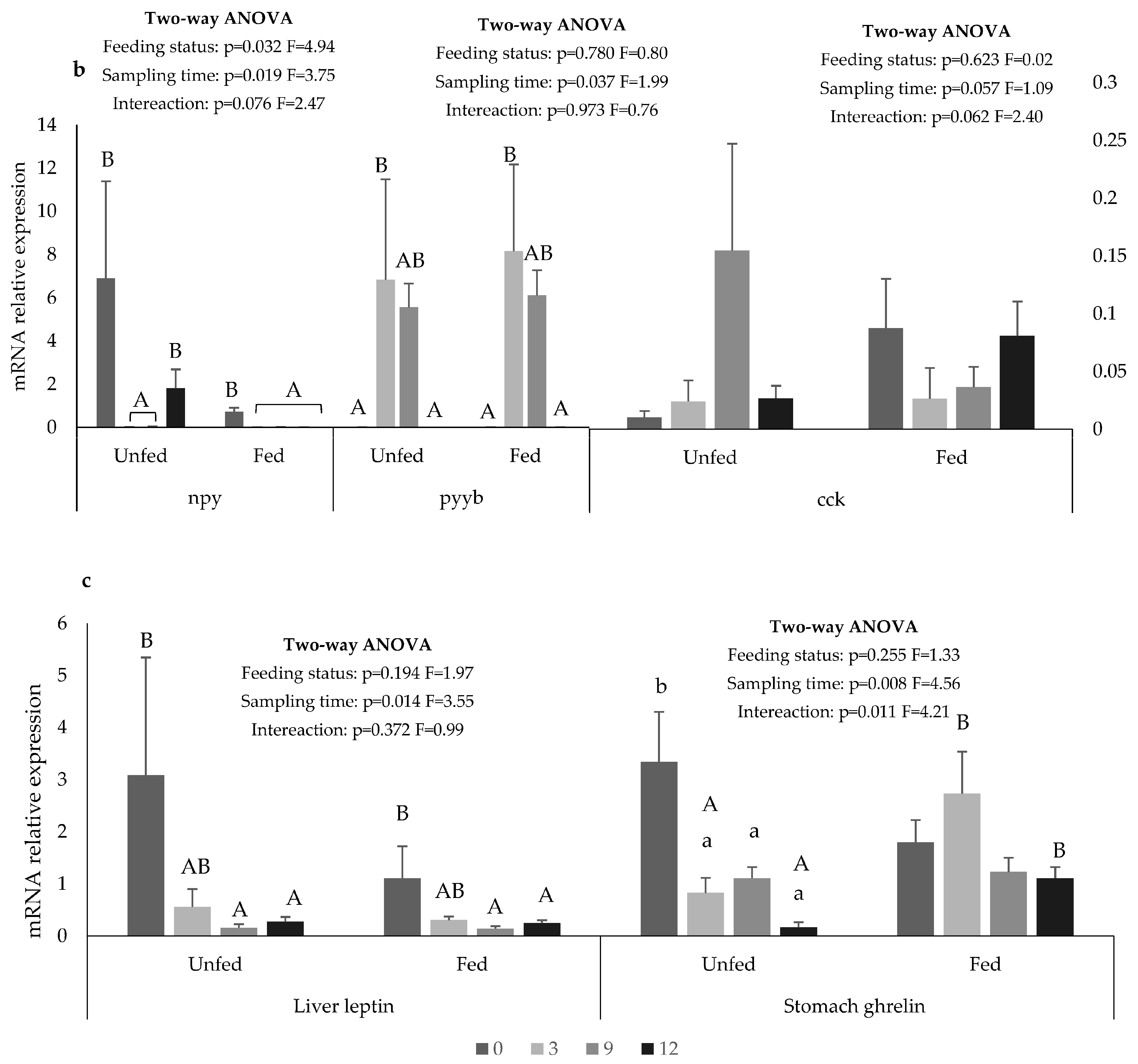
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Food and Agriculture Organization of the United Nations. The State of the World Fisheries and Aquaculture-Meeting the Sustainable Development Goals; FAO: Rome, Italy, 2022; Volume 50, pp. 50–5350. [Google Scholar] [CrossRef] [Green Version]
- Hasan, M.R. Feeding Global Aquaculture Growth. FAO Aquac. Newsl. 2017, 56, 2–3. [Google Scholar]
- Soengas, J.L.; Cerdá-Reverter, J.M.; Delgado, M.J. Central Regulation of Food Intake in Fish: An Evolutionary Perspective. J. Mol. Endocrinol. 2018, 60, R171–R199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volkoff, H.; Canosa, L.F.F.; Unniappan, S.; Cerdá-Reverter, J.M.M.; Bernier, N.J.J.; Kelly, S.P.P.; Peter, R.E.E. Neuropeptides and the Control of Food Intake in Fish. In General, and Comparative Endocrinology; Academic Press Inc.: Cambridge, MA, USA, 15 May 2005; Volume 142, pp. 3–19. [Google Scholar]
- Volkoff, H. The Neuroendocrine Regulation of Food Intake in Fish: A Review of Current Knowledge. Front. Neurosci. 2016, 10, 540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delgado, M.J.; Cerdá-Reverter, J.M.; Soengas, J.L. Hypothalamic Integration of Metabolic, Endocrine, and Circadian Signals in Fish: Involvement in the Control of Food Intake. Front. Neurosci. 2017, 11, 354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rønnestad, I.; Gomes, A.S.; Murashita, K.; Angotzi, R.; Jönsson, E.; Volkoff, H. Appetite-Controlling Endocrine Systems in Teleosts. Front. Endocrinol. 2017, 8, 73. [Google Scholar] [CrossRef] [Green Version]
- Sundström, G.; Larsson, T.A.; Brenner, S.; Venkatesh, B.; Larhammar, D. Evolution of the Neuropeptide Y Family: New Genes by Chromosome Duplications in Early Vertebrates and in Teleost Fishes. Gen. Comp. Endocrinol. 2008, 155, 705–716. [Google Scholar] [CrossRef]
- Assan, D.; Mustapha, U.F.; Chen, H.; Li, Z.; Peng, Y.; Li, G. The Roles of Neuropeptide Y (Npy) and Peptide YY (Pyy) in Teleost Food Intake: A Mini Review. Life 2021, 11, 547. [Google Scholar] [CrossRef]
- Loh, K.; Herzog, H.; Shi, Y.C. Regulation of Energy Homeostasis by the NPY System; Elsevier Current Trends: Amsterdam, The Netherlands, 2015; Volume 26, pp. 125–135. [Google Scholar]
- Kehoe, A.S.; Volkoff, H. Cloning and Characterization of Neuropeptide Y (NPY) and Cocaine and Amphetamine Regulated Transcript (CART) in Atlantic Cod (Gadus morhua). Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2007, 146, 451–461. [Google Scholar] [CrossRef]
- Narnaware, Y.K.; Peter, R.E. Effects of Food Deprivation and Refeeding on Neuropeptide Y (NPY) MRNA Levels in Goldfish. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2001, 129, 633–637. [Google Scholar] [CrossRef]
- Peterson, B.C.; Waldbieser, G.C.; Riley, L.G.; Upton, K.R.; Kobayashi, Y.; Small, B.C. Pre- and Postprandial Changes in Orexigenic and Anorexigenic Factors in Channel Catfish (Ictalurus punctatus). Gen. Comp. Endocrinol. 2012, 176, 231–239. [Google Scholar] [CrossRef]
- MacDonald, E.; Volkoff, H. Neuropeptide Y (NPY), Cocaine- and Amphetamine-Regulated Transcript (CART) and Cholecystokinin (CCK) in Winter Skate (Raja ocellata): CDNA Cloning, Tissue Distribution and MRNA Expression Responses to Fasting. Gen. Comp. Endocrinol. 2009, 161, 252–261. [Google Scholar] [CrossRef]
- Hosomi, N.; Furutani, T.; Takahashi, N.; Masumoto, T.; Fukada, H. Yellowtail Neuropeptide Y: Molecular Cloning, Tissue Distribution, and Response to Fasting. Fish Sci. 2014, 80, 483–492. [Google Scholar] [CrossRef]
- Wei, R.; Zhou, C.; Yuan, D.; Wang, T.; Lin, F.; Chen, H.; Wu, H.; Xin, Z.; Yang, S.; Wang, Y.; et al. Characterization, Tissue Distribution and Regulation of NeuropeptideY in Schizothorax Prenanti. J. Fish Biol. 2014, 85, 278–291. [Google Scholar] [CrossRef]
- Narnaware, Y.K.; Peyon, P.P.; Lin, X.; Peter, R.E.C.N.-C. Regulation of Food Intake by Neuropeptide Y in Goldfish. Am. J. Physiol. Reg. I 2000, 279, R1025–R1034. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Tan, X.; Sui, Y.; Jiao, S.; Wu, Z.; Wang, L.; You, F. The Stimulatory Effect of Neuropeptide Y on Growth Hormone Expression, Food Intake, and Growth in Olive Flounder (Paralichthys olivaceus). Fish Physiol. Biochem. 2017, 43, 11–18. [Google Scholar] [CrossRef]
- Valassi, E.; Scacchi, M.; Cavagnini, F. Neuroendocrine Control of Food Intake. Nutr. Metab. Cardiovasc. Dis. 2008, 18, 158–168. [Google Scholar] [CrossRef]
- Holzer, P.; Reichmann, F.; Farzi, A. Neuropeptide Y, Peptide YY and Pancreatic Polypeptide in the Gut-Brain Axis. Neuropeptides 2012, 46, 261–274. [Google Scholar] [CrossRef] [Green Version]
- Velasco, C.; Blanco, A.M.; Unniappan, S.; Soengas, J.L. The Anorectic Effect of Central PYY1-36 Treatment in Rainbow Trout (Oncorhynchus mykiss) Is Associated with Changes in MRNAs Encoding Neuropeptides and Parameters Related to Fatty Acid Sensing and Metabolism. Gen. Comp. Endocrinol. 2018, 267, 137–145. [Google Scholar] [CrossRef]
- Yan, P.; Jia, J.; Yang, G.; Wang, D.; Sun, C.; Li, W. Duplication of Neuropeptide Y and Peptide YY in Nile Tilapia Oreochromis Niloticus and Their Roles in Food Intake Regulation. Peptides 2017, 88, 97–105. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez, R.; Unniappan, S. Molecular Characterization, Appetite Regulatory Effects and Feeding Related Changes of Peptide YY in Goldfish. Gen. Comp. Endocrinol. 2010, 166, 273–279. [Google Scholar] [CrossRef]
- Yuan, D.; Zhou, C.; Wang, T.; Lin, F.; Chen, H.; Wu, H.; Wei, R.; Xin, Z.; Liu, J.; Gao, Y.; et al. Molecular Characterization and Tissue Expression of Peptide YY in Schizothorax Prenanti: Effects of Periprandial Changes and Fasting on Expression in the Hypothalamus. Regul. Pept. 2014, 190–191, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Gomes, A.S.; Jordal, A.-E.O.; Olsen, K.; Harboe, T.; Power, D.M.; Rønnestad, I. Neuroendocrine Control of Appetite in Atlantic Halibut (Hippoglossus hippoglossus): Changes during Metamorphosis and Effects of Feeding. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2015, 183, 116–125. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Zhang, X.; Hao, J.; Chen, D.; Liu, J.; Gao, Y.; Zhu, J.; Wu, H.; Lin, F.; Pu, Y.; et al. Molecular Cloning, Expression Analysis, and Appetite Regulatory Effect of Peptide YY in Siberian Sturgeon (Acipenser baerii). Gene 2015, 563, 172–179. [Google Scholar] [CrossRef]
- Wall, A.; Volkoff, H. Effects of Fasting and Feeding on the Brain MRNA Expressions of Orexin, Tyrosine Hydroxylase (TH), PYY and CCK in the Mexican Blind Cavefish (Astyanax fasciatus mexicanus). Gen. Comp. Endocrinol. 2013, 183, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Volkoff, H. Appetite Regulating Peptides in Red-Bellied Piranha, Pygocentrus Nattereri: Cloning, Tissue Distribution and Effect of Fasting on MRNA Expression Levels. Peptides 2014, 56, 116–124. [Google Scholar] [CrossRef]
- Ollmann, M.M.; Wilson, B.D.; Yang, Y.; Kerns, J.A.; Gantz, I.; Barsh, G.S.; Ollmann, M.M.; Wilson, B.D.; Yang, Y.; Kerns, J.A.; et al. Antagonism of Central Melanocortin Receptores in Vitro and in Vivo by Agouti-Related Protein. Science 1997, 278, 135–138. [Google Scholar] [CrossRef]
- Cerda, M.; Peter, R.E. Endogenous Melanocortin Antagonist in Fish: Structure, Brain Mapping, and Regulation by Fasting of the Goldfish Agouti-Related Protein Gene. Endocrinology 2003, 144, 4552–4561. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.; Golling, G.; Thacker, T.L.; Cone, R.D. Agouti-Related Protein (AGRP) Is Conserved and Regulated by Metabolic State in the Zebrafish, Danio rerio. Endocrine 2003, 22, 257–265. [Google Scholar] [CrossRef]
- Murashita, K.; Kurokawa, T.; Nilsen, T.O.; Rønnestad, I. Ghrelin, Cholecystokinin, and Peptide YY in Atlantic Salmon (Salmo salar): Molecular Cloning and Tissue Expression. Gen. Comp. Endocrinol. 2009, 160, 223–235. [Google Scholar] [CrossRef]
- Lau, J.; Herzog, H. CART in the Regulation of Appetite and Energy Homeostasis. Front. Neurosci. 2014, 8, 313. [Google Scholar] [CrossRef]
- Volkoff, H.; Peter, R.E. Effects of CART Peptides on Food Consumption, Feeding and Associated Behaviors in the Goldfish, Carassius auratus: Actions on Neuropeptide Y- and Orexin A-Induced Feeding. Brain Res. 2000, 887, 125–133. [Google Scholar] [CrossRef]
- Murashita, K.; Kurokawa, T.; Ebbesson, L.O.E.; Stefansson, S.O.; Rønnestad, I. Characterization, Tissue Distribution, and Regulation of Agouti-Related Protein (AgRP), Cocaine- and Amphetamine-Regulated Transcript (CART) and Neuropeptide Y (NPY) in Atlantic Salmon (Salmo salar). Gen. Comp. Endocrinol. 2009, 162, 160–171. [Google Scholar] [CrossRef]
- Fukada, H.; Murashita, K.; Senzui, A.; Masumoto, T. Multiple Cocaine- and Amphetamine-Regulated Transcript Genes in Yellowtail Seriola quinqueradiata: Cloning, Tissue Distribution in the Brain, and Response to Fasting and Fish Meal Soluble Fraction. Fish. Sci. 2021, 87, 55–64. [Google Scholar] [CrossRef]
- Bonacic, K.; Martínez, A.; Martín-Robles, Á.J.; Muñoz-Cueto, J.A.; Morais, S. Characterization of Seven Cocaine- and Amphetamine-Regulated Transcripts (CARTs) Differentially Expressed in the Brain and Peripheral Tissues of Solea senegalensis (Kaup). Gen. Comp. Endocrinol. 2015, 224, 260–272. [Google Scholar] [CrossRef]
- Coll, A.P.; Farooqi, I.S.; Challis, B.G.; Yeo, G.S.H.; O’Rahilly, S. Proopiomelanocortin and Energy Balance: Insights from Human and Murine Genetics. J. Clin. Endocrinol. Metab. 2004, 89, 2557–2562. [Google Scholar] [CrossRef] [Green Version]
- Kang, D.Y.; Kim, H.C. Functional Relevance of Three Proopiomelanocortin (POMC) Genes in Darkening Camouflage, Blind-Side Hypermelanosis, and Appetite of Paralichthys olivaceus. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2015, 179, 44–56. [Google Scholar] [CrossRef]
- MacDonald, E.; Volkoff, H. Cloning, Distribution and Effects of Season and Nutritional Status on the Expression of Neuropeptide Y (NPY), Cocaine and Amphetamine Regulated Transcript (CART) and Cholecystokinin (CCK) in Winter Flounder (Pseudopleuronectes Americanus). Horm. Behav. 2009, 56, 58–65. [Google Scholar] [CrossRef]
- Ji, W.; Ping, H.C.; Wei, K.J.; Zhang, G.R.; Shi, Z.C.; Yang, R.B.; Zou, G.W.; Wang, W.M. Ghrelin, Neuropeptide Y (NPY) and Cholecystokinin (CCK) in Blunt Snout Bream (Megalobrama amblycephala): CDNA Cloning, Tissue Distribution and MRNA Expression Changes Responding to Fasting and Refeeding. Gen. Comp. Endocrinol. 2015, 223, 108–119. [Google Scholar] [CrossRef]
- Micale, V.; Campo, S.; D’Ascola, A.; Guerrera, M.C.; Levanti, M.B.; Germanà, A.; Muglia, U. Cholecystokinin in White Sea Bream: Molecular Cloning, Regional Expression, and Immunohistochemical Localization in the Gut after Feeding and Fasting. PLoS ONE 2012, 7, e52428. [Google Scholar] [CrossRef] [Green Version]
- Unniappan, S.; Peter, R.E. Structure, Distribution and Physiological Functions of Ghrelin in Fish. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2005, 140, 396–408. [Google Scholar] [CrossRef]
- Unniappan, S.; Canosa, L.F.; Peter, R.E. Orexigenic Actions of Ghrelin in Goldfish: Feeding-Induced Changes in Brain and Gut MRNA Expression and Serum Levels, and Responses to Central and Peripheral Injections. Neuroendocrinology 2004, 79, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Miura, T.; Maruyama, K.; Shimakura, S.I.; Kaiya, H.; Uchiyama, M.; Kangawa, K.; Shioda, S.; Matsuda, K. Neuropeptide Y Mediates Ghrelin-Induced Feeding in the Goldfish, Carassius auratus. Neurosci. Lett. 2006, 407, 279–283. [Google Scholar] [CrossRef] [PubMed]
- Tinoco, A.B.; Näslund, J.; Delgado, M.J.; de Pedro, N.; Johnsson, J.I.; Jönsson, E. Ghrelin Increases Food Intake, Swimming Activity and Growth in Juvenile Brown Trout (Salmo trutta). Physiol. Behav. 2014, 124, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Penney, C.C.; Volkoff, H. Peripheral Injections of Cholecystokinin, Apelin, Ghrelin and Orexin in Cavefish (Astyanax fasciatus mexicanus): Effects on Feeding and on the Brain Expression Levels of Tyrosine Hydroxylase, Mechanistic Target of Rapamycin and Appetite-Related Hormones. Gen. Comp. Endocrinol. 2014, 196, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Velasco, C.; Librán-Pérez, M.; Otero-Rodiño, C.; López-Patiño, M.A.; Míguez, J.M.; Cerdá-Reverter, J.M.; Soengas, J.L. Ghrelin Modulates Hypothalamic Fatty Acid-Sensing and Control of Food Intake in Rainbow Trout. J. Endocrinol. 2016, 228, 25–37. [Google Scholar] [CrossRef] [Green Version]
- Jönsson, E.; Kaiya, H.; Björnsson, B.T. Ghrelin Decreases Food Intake in Juvenile Rainbow Trout (Oncorhynchus mykiss) through the Central Anorexigenic Corticotropin-Releasing Factor System. Gen. Comp. Endocrinol. 2010, 166, 39–46. [Google Scholar] [CrossRef]
- Houseknecht, K.L.; Baile, C.A.; Matteri, R.L.; Spurlock, M.E. The Biology of Leptin: A Review. J. Anim. Sci. 1998, 76, 1405–1420. [Google Scholar] [CrossRef] [Green Version]
- Copeland, D.L.; Duff, R.J.; Liu, Q.; Prokop, J.; Londraville, R.L. Leptin in Teleost Fishes: An Argument for Comparative Study. Front. Physiol. 2011, 2, 26. [Google Scholar] [CrossRef] [Green Version]
- Heo, W.S.; Kim, Y.R.; Kim, E.Y.; Bai, S.C.; Kong, I.S. Effects of Dietary Probiotic, Lactococcus Lactis Subsp. Lactis I2, Supplementation on the Growth and Immune Response of Olive Flounder (Paralichthys olivaceus). Aquaculture 2013, 376–379, 20–24. [Google Scholar] [CrossRef]
- Huising, M.O.; Geven, E.J.W.; Kruiswijk, C.P.; Nabuurs, S.B.; Stolte, E.H.; Spanings, F.A.T.; Lidy Verburg-Van Kemenade, B.M.; Flik, G. Increased Leptin Expression in Common Carp (Cyprinus carpio) after Food Intake but Not after Fasting or Feeding to Satiation. Endocrinology 2006, 147, 5786–5797. [Google Scholar] [CrossRef]
- EUMOFA European Seabass (Dicentrarchus labrax)—Species Profile. Available online: https://www.eumofa.eu/documents/20178/137160/European+seabass_31-1.pdf (accessed on 30 August 2021).
- Pfaffl, M.W. A New Mathematical Model for Relative Quantification in Real-Time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate Normalization of Real-Time Quantitative RT-PCR Data by Geometric Averaging of Multiple Internal Control Genes. Genome Biol. 2002, 3, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Cerdá-Reverter, J.M.; Anglade, I.; Martínez-Rodríguez, G.; Mazurais, D.; Muñoz-Cueto, J.A.; Carrillo, M.; Kah, O.; Zanuy, S. Characterization of Neuropeptide Y Expression in the Brain of a Perciform Fish, the Sea Bass (Dicentrarchus labrax). J. Chem. Neuroanat. 2000, 19, 197–210. [Google Scholar] [CrossRef]
- Agulleiro, M.J.; Cortés, R.; Leal, E.; Ríos, D.; Sánchez, E.; Cerdá-Reverter, J.M. Characterization, Tissue Distribution and Regulation by Fasting of the Agouti Family of Peptides in the Sea Bass (Dicentrarchus labrax). Gen. Comp. Endocrinol. 2014, 205, 251–259. [Google Scholar] [CrossRef]
- Varsamos, S.; Wendelaar Bonga, S.E.; Flik, G.; Queré, R.; Commes, T. Cloning of a Proopiomelanocortin CDNA from the Pituitary Gland of the Sea Bass (Dicentrarchus labrax) and Assessment of MRNA Expression in Different Tissues by Means of Real-Time PCR. J. Endocrinol. 2003, 176, 405–414. [Google Scholar] [CrossRef] [Green Version]
- Gambardella, C.; Gallus, L.; Amaroli, A.; Terova, G.; Masini, M.A.; Ferrando, S. Fasting and Re-Feeding Impact on Leptin and Aquaglyceroporin 9 in the Liver of European Sea Bass (Dicentrarchus labrax). Aquaculture 2012, 354–355, 1–6. [Google Scholar] [CrossRef]
- Terova, G.; Rimoldi, S.; Bernardini, G.; Gornati, R.; Saroglia, M. Sea Bass Ghrelin: Molecular Cloning and MRNA Quantification during Fasting and Refeeding. Gen. Comp. Endocrinol. 2008, 155, 341–351. [Google Scholar] [CrossRef]
- Tillner, R.; Rønnestad, I.; Dhert, P.; Ueberschär, B. The Regulatory Loop between Gut Cholecystokinin and Tryptic Enzyme Activitcxuy in Sea Bass (Dicentrarchus labrax) Larvae Is Influenced by Different Feeding Regimes and Trigger Substances. Aquaculture 2014, 420–421, 139–146. [Google Scholar] [CrossRef]
- Peres, H.; Santos, S.; Oliva-Teles, A. Blood Chemistry Profile as Indicator of Nutritional Status in European Seabass (Dicentrarchus labrax). Fish Physiol. Biochem. 2014, 40, 1339–1347. [Google Scholar] [CrossRef]
- Volkoff, H. The Role of Neuropeptide Y, Orexins, Cocaine and Amphetamine-Related Transcript, Cholecystokinin, Amylin and Leptin in the Regulation of Feeding in Fish. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2006, 144, 325–331. [Google Scholar] [CrossRef]
- Hoskins, L.J.; Volkoff, H. Daily Patterns of MRNA Expression of Two Core Circadian Regulatory Proteins, Clock2 and Per1, and Two Appetite-Regulating Peptides, OX and NPY, in Goldfish (Carassius auratus). Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2012, 163, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Narnaware, Y.K.; Peter, R.E. Influence of Diet Composition on Food Intake and Neuropeptide Y (NPY) Gene Expression in Goldfish Brain. Regul. Pept. 2002, 103, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Campos, V.F.; Robaldo, R.B.; Deschamps, J.C.; Seixas, F.K.; McBride, A.J.A.; Marins, L.F.; Okamoto, M.; Sampaio, L.A.; Collares, T. Neuropeptide Y Gene Expression around Mealtime in the Brazilian Flounder Paralichthys orbignyanus. J. Biosci. 2012, 37, 227–232. [Google Scholar] [CrossRef] [PubMed]
- Bjenning, C.; Holmgren, S.; Farrell, A.P. Neuropeptide Y Potentiates Contractile Response to Norepinephrine in Skate Coronary Artery. Am. J. Physiol. Hear. Circ. Physiol. 1993, 265, H661–H665. [Google Scholar] [CrossRef] [PubMed]
- Bjenning, C.; Hazon, N.; Balasubramaniam, A.; Holmgren, S.; Conlon, J.M. Distribution and Activity of Dogfish NPY and Peptide YY in the Cardiovascular System of the Common Dogfish. Am. J. Physiol. Regul. Integr. Comp. Physiol. 1993, 264, 1119. [Google Scholar] [CrossRef]
- Preston, E.; Jonsson, A.C.; McManus, C.D.; Conlon, J.M.; Courtice, G.P. Comparative Vascular Responses in Elasmobranchs to Different Structures of Neuropeptide Y and Peptide YY. Regul. Pept. 1998, 78, 57–67. [Google Scholar] [CrossRef]
- Shahbazi, F.; Holmgren, S.; Larhammar, D.; Jensen, J. Neuropeptide Y Effects on Vasorelaxation and Intestinal Contraction in the Atlantic Cod Gadus morhua. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2002, 282, 1414–1421. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Liang, X.F.; Yuan, X.; Li, J.; He, Y.; Fang, L.; Guo, X.; Liu, L.; Li, B.; Shen, D. Neuropeptide Y Stimulates Food Intake and Regulates Metabolism in Grass Carp, Ctenopharyngodon idellus. Aquaculture 2013, 380–383, 52–61. [Google Scholar] [CrossRef]
- Cerdá-Reverter, J.M.; Martínez-Rodríguez, G.; Zanuy, S.; Carrillo, M.; Larhammar, D. Molecular Evolution of the Neuropeptide Y (NPY) Family of Peptides: Cloning of Three NPY-Related Peptides from the Sea Bass (Dicentrarchus labrax). Regul. Pept. 2000, 95, 25–34. [Google Scholar] [CrossRef]
- Sundström, G.; Larsson, T.A.; Brenner, S.; Venkatesh, B.; Larhammar, D. Ray-Fin Fish Tetraploidization Gave Rise to Pufferfish Duplicates of NPY and PYY, but Zebrafish NPY Duplicate Was Lost. Ann. N. Y. Acad. Sci. 2005, 1040, 476–478. [Google Scholar] [CrossRef]
- Kalananthan, T.; Murashita, K.; Rønnestad, I.; Ishigaki, M.; Takahashi, K.; Silva, M.S.; Wakabayashi, Y.; Lai, F.; Shimizu, M.; Nilsen, T.O.; et al. Hypothalamic Agrp and Pomc MRNA Responses to Gastrointestinal Fullness and Fasting in Atlantic Salmon (Salmo salar, L.). Front. Physiol. 2020, 11, 61. [Google Scholar] [CrossRef] [Green Version]
- Drew, R.E.; Rodnick, K.J.; Settles, M.; Wacyk, J.; Churchill, E.; Powell, M.S.; Hardy, R.W.; Murdoch, G.K.; Hill, R.A.; Robison, B.D. Effect of Starvation on Transcriptomes of Brain and Liver in Adult Female Zebrafish (Danio rerio). Physiol. Genomics 2008, 35, 283–295. [Google Scholar] [CrossRef] [Green Version]
- Striberny, A.; Ravuri, C.S.; Jobling, M.; Jørgensen, E.H.; Fuentes, J. Seasonal Differences in Relative Gene Expression of Putative Central Appetite Regulators in Arctic Charr (Salvelinus alpinus) Do Not Reflect Its Annual Feeding Cycle. PLoS ONE 2015, 10, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Basto, A.; Valente, L.M.P.; Conde-Sieira, M.; Soengas, J.L. Central Regulation of Food Intake Is Not Affected by Inclusion of Defatted Tenebrio Molitor Larvae Meal in Diets for European Sea Bass (Dicentrarchus labrax). Aquaculture 2021, 544, 737088. [Google Scholar] [CrossRef]
- Valen, R.; Jordal, A.E.O.; Murashita, K.; Rønnestad, I. Postprandial Effects on Appetite-Related Neuropeptide Expression in the Brain of Atlantic salmon, Salmo salar. Gen. Comp. Endocrinol. 2011, 171, 359–366. [Google Scholar] [CrossRef]
- Leder, E.H.; Silverstein, J.T. The Pro-Opiomelanocortin Genes in Rainbow Trout (Oncorhynchus mykiss): Duplications, Splice Variants, and Differential Expression. J. Endocrinol. 2006, 188, 355–363. [Google Scholar] [CrossRef]
- Rubio, V.C.C.; Sánchez-Vázquez, F.J.J.; Madrid, J.A.A. Role of Cholecystokinin and Its Antagonist Proglumide on Macronutrient Selection in European Sea Bass Dicentrarchus labrax, L. Physiol. Behav. 2008, 93, 862–869. [Google Scholar] [CrossRef]
- Murashita, K.; Fukada, H.; Hosokawa, H.; Masumoto, T. Changes in Cholecystokinin and Peptide Y Gene Expression with Feeding in Yellowtail (Seriola quinqueradiata): Relation to Pancreatic Exocrine Regulation. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2007, 146, 318–325. [Google Scholar] [CrossRef]
- Murashita, K.; Fukada, H.; Rønnestad, I.; Kurokawa, T.; Masumoto, T. Nutrient Control of Release of Pancreatic Enzymes in Yellowtail (Seriola quinqueradiata): Involvement of CCK and PY in the Regulatory Loop. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2008, 150, 438–443. [Google Scholar] [CrossRef]
- Babichuk, N.A.; Volkoff, H. Changes in Expression of Appetite-Regulating Hormones in the Cunner (Tautogolabrus Adspersus) during Short-Term Fasting and Winter Torpor. Physiol. Behav. 2013, 120, 54–63. [Google Scholar] [CrossRef]
- Koven, W.; Schulte, P. The Effect of Fasting and Refeeding on MRNA Expression of PepT1 and Gastrointestinal Hormones Regulating Digestion and Food Intake in Zebrafish (Danio rerio). Fish Physiol. Biochem. 2012, 38, 1565–1575. [Google Scholar] [CrossRef] [PubMed]
- Feng, K.; Zhang, G.R.; Wei, K.J.; Xiong, B.X. Molecular Cloning, Tissue Distribution, and Ontogenetic Expression of Ghrelin and Regulation of Expression by Fasting and Refeeding in the Grass Carp (Ctenopharyngodon idellus). J. Exp. Zool. Part A Ecol. Genet. Physiol. 2013, 319, 202–212. [Google Scholar] [CrossRef] [PubMed]
- Eom, J.; Hong, M.; Cone, R.D.; Song, Y. Zebrafish Ghrelin Is Expressed in Pancreatic Endocrine Cells and Regulated by Metabolic State. Biochem. Biophys. Res. Commun. 2013, 439, 115–120. [Google Scholar] [CrossRef]
- Hevrøy, E.M.; Waagbø, R.; Torstensen, B.E.; Takle, H.; Stubhaug, I.; Jørgensen, S.M.; Torgersen, T.; Tvenning, L.; Susort, S.; Breck, O.; et al. Ghrelin Is Involved in Voluntary Anorexia in Atlantic salmon Raised at Elevated Sea Temperatures. Gen. Comp. Endocrinol. 2012, 175, 118–134. [Google Scholar] [CrossRef] [PubMed]
- Blanco, A.M.; Soengas, J.L. Leptin Signalling in Teleost Fish with Emphasis in Food Intake Regulation. Mol. Cell. Endocrinol. 2021, 526, 111209. [Google Scholar] [CrossRef]
- Won, E.T.; Baltzegar, D.A.; Picha, M.E.; Borski, R.J. Cloning and Characterization of Leptin in a Perciform Fish, the Striped Bass (Morone saxatilis): Control of Feeding and Regulation by Nutritional State. Gen. Comp. Endocrinol. 2012, 178, 98–107. [Google Scholar] [CrossRef]
- Yuan, D.; Wang, T.; Zhou, C.; Lin, F.; Chen, H.; Wu, H.; Wei, R.; Xin, Z.; Li, Z. Leptin and Cholecystokinin in Schizothorax Prenanti: Molecular Cloning, Tissue Expression, and MRNA Expression Responses to Periprandial Changes and Fasting. Gen. Comp. Endocrinol. 2014, 204, 13–24. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence (5′-3′) | Ta (°C) | Efficiency | Accession Number |
|---|---|---|---|---|
| Central | ||||
| agrp1 | F: TCTGCTGCGTCTCTCTTCTT | 62.0 | 2.01 | HE660086 |
| R: TCTCTCCAGGTCAGACAGGT | ||||
| agrp2 | F: GGGCAGAGGACACAAAGAAA | 62.0 | 2.01 | HE660087 |
| R: TGTGACTTTCCTGTGGTGGA | ||||
| cart2 | F: CCGAACCTGACCAGCGAGAA | 61.0 | 1.92 | MZ441181 |
| R: GCTCCCCGACATCACACGTT | ||||
| npy | F: ACTCAGCCCTGAGACACTACA | 50.8 | 2.01 | AJ005378 |
| R: ACTGTGGAAGCTGGTCTGTG | ||||
| pomc1 | F: TCCCTGTTCTCTTCCTCCTC | 57.6 | 2.01 | AY691808 |
| R: CTCTGCCGCTATAATCTCGC | ||||
| Peripheral | ||||
| cck | F: TTCTCCTCCCAGCTCTTTGA | 60.0 | 1.92 | DLA_LG9_001730 |
| R: GGAGTCTGCATCCTCCTCAG | ||||
| ghr | F: TTGCTGGTGGTTCTGTTGTG | 61.0 | 1.95 | DQ665912 |
| R: TTGCCTCTGCTCTGAGGTTT | ||||
| lep | F: TCAGTCAGTGGAGGCAAGAG | 60.0 | 1.92 | KJ934254 |
| R: TTTCAGCAGCAGATTGAGGA | ||||
| pyyb | F: GGTGTGTTTGAGCAGCTTCG | 60.0 | 1.92 | AJ005380 |
| R: CAGCGTCTTGGCTTGAATCG | ||||
| Housekeeping | ||||
| ef1α | F: GCTTCGAGGAAATCACCAAG | 60.0 | 1.97 | AJ866727 |
| R: CAACCTTCCATCCCTTGAAC | ||||
| 18s | F: GGAGGTTCGAAGACGATCAG | 63.0 | 2.01 | AY831388.1 |
| R: GAGGTTTCCCGTGTTGAGTC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martins, N.; Castro, C.; Oliva-Teles, A.; Peres, H. The Interplay between Central and Peripheral Systems in Feed Intake Regulation in European Seabass (Dicentrarchus labrax) Juveniles. Animals 2022, 12, 3287. https://doi.org/10.3390/ani12233287
Martins N, Castro C, Oliva-Teles A, Peres H. The Interplay between Central and Peripheral Systems in Feed Intake Regulation in European Seabass (Dicentrarchus labrax) Juveniles. Animals. 2022; 12(23):3287. https://doi.org/10.3390/ani12233287
Chicago/Turabian StyleMartins, Nicole, Carolina Castro, Aires Oliva-Teles, and Helena Peres. 2022. "The Interplay between Central and Peripheral Systems in Feed Intake Regulation in European Seabass (Dicentrarchus labrax) Juveniles" Animals 12, no. 23: 3287. https://doi.org/10.3390/ani12233287