
Effect of Methylmercury Exposure on Bioaccumulation and Nonspecific Immune Respsonses in Hybrid Grouper Epinephelus fuscoguttatus × Epinephelus lanceolatus


 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. The In Vitro Effect of Methylmercury
2.1.1. Cell Viability
2.1.2. Phagocytic Activity Assay
2.1.3. Superoxide Anion (O2−) Production Assay
2.2. The In Vivo Effect of Methylmercury
2.2.1. Dietary Exposure of Methylmercury
2.2.2. Measurement of Methylmercury Bioaccumulation
2.3. Statistical Analysis
3. Results
3.1. Effect of Methylmercury on Cell Viability In Vitro
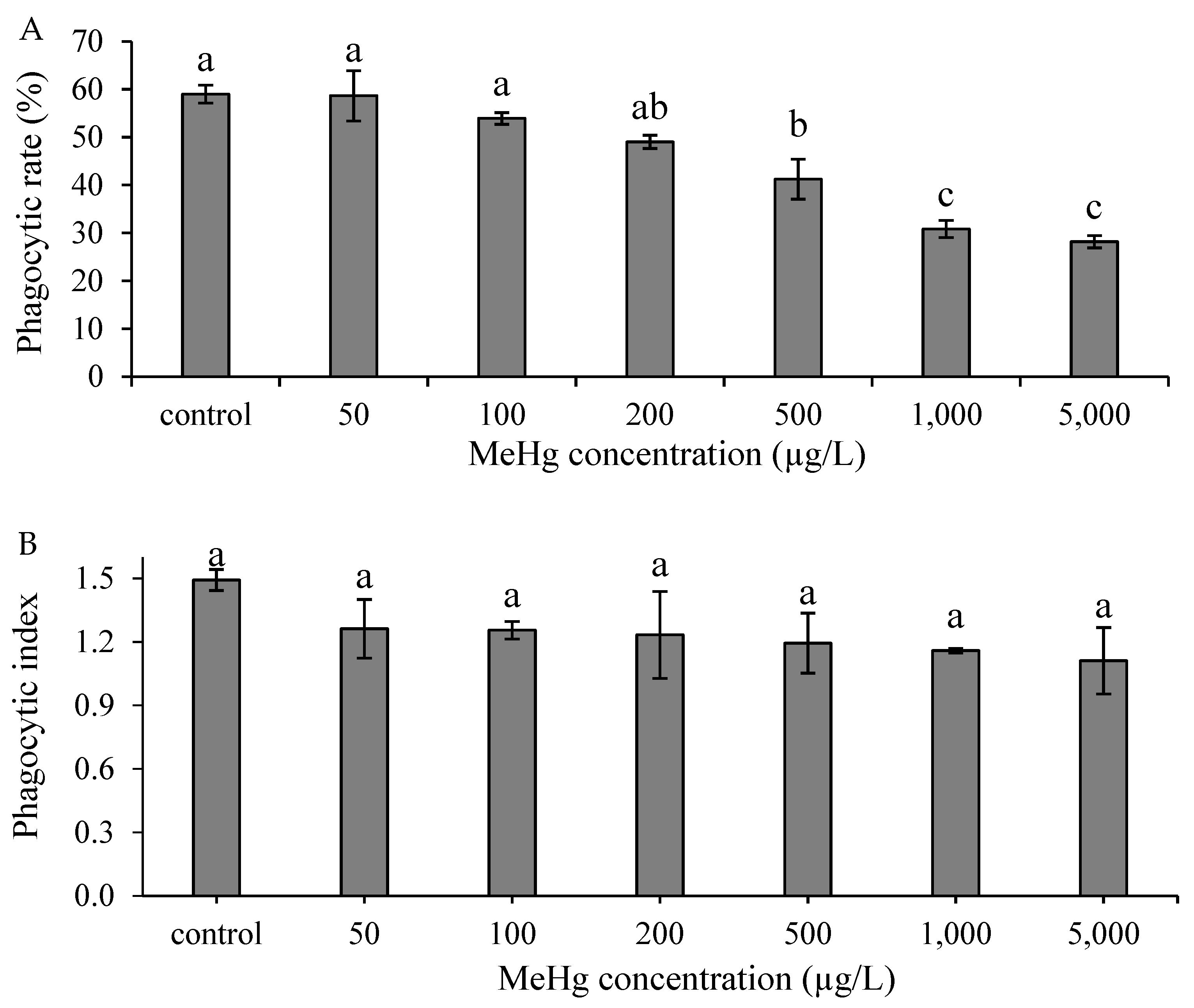
3.2. Effect of Methylmercury on Nonspecific Immune Responses In Vitro
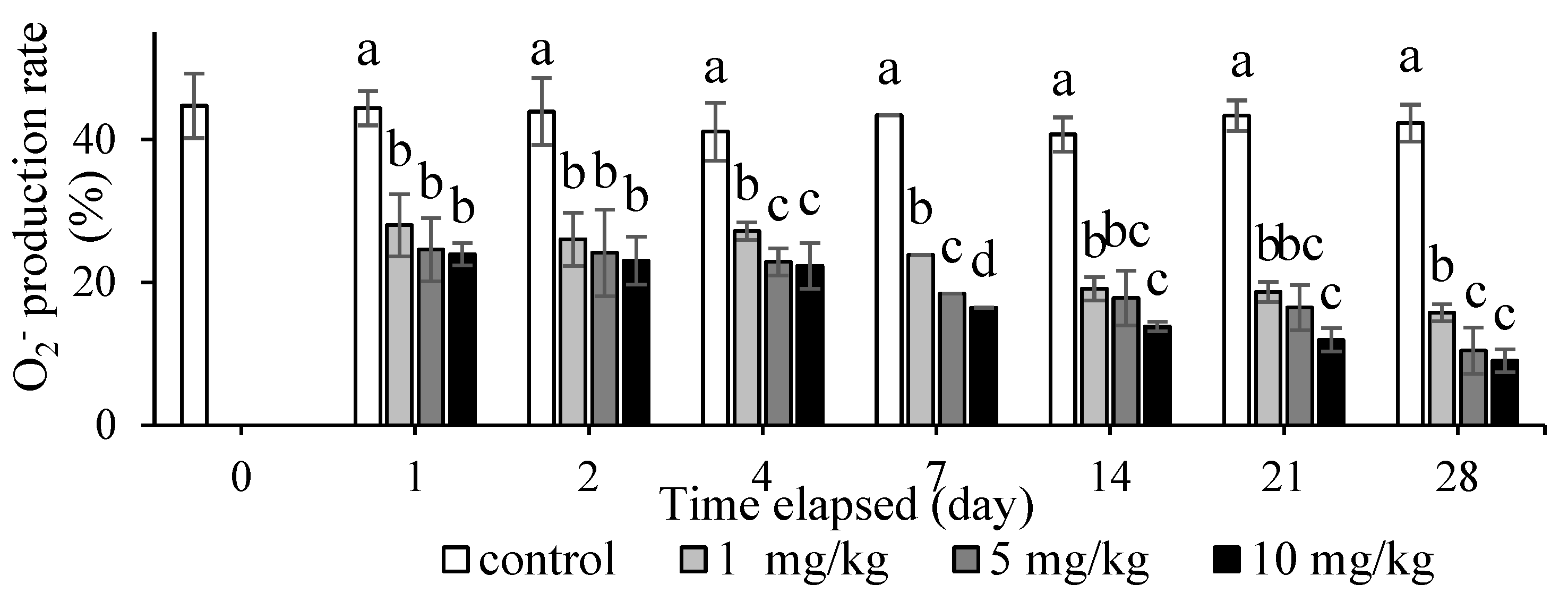
3.3. Effect of Methylmercury Dietary Exposure on Nonspecific Immune Responses
3.4. Level of Methylmercury Bioaccumulation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture 2020: Sustainability in Action. Available online: https://www.fao.org/documents/card/en/c/ca9229en/ (accessed on 17 November 2021).
- Che Ismail, C.-Z.; Koh, I.; Md sheriff, S. Cryopreservation of spermatozoa on grouper species: A review. Rev. Aquac. 2018, 12, 26–32. [Google Scholar]
- Afero, F.; Miao, S.; Perez, A.A. Economic analysis of tiger grouper Epinephelus fuscoguttatus and humpback grouper Cromileptes altivelis commercial cage culture in Indonesia. Aquac. Int. 2009, 18, 725–739. [Google Scholar] [CrossRef]
- Fan, B.; Yang, S.; Wang, L.; Chen, X.; Liu, X.; Zhang, Y.; Li, S.; Zhang, H.; Meng, Z.; Lin, H. Hybridization of tiger grouper (Epinephelus fuscoguttatus ♀) x giant grouper (Epinephelus lanceolatus ♂) using cryopreserved sperm. Cryobiology 2020, 95, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Ching, F.F.; Othman, N.; Anuar, A.; Shapawi, R.; Senoo, S. Natural spawning, embryonic and larval development of F2 hybrid grouper, tiger grouper Epinephelus fuscoguttatus × giant grouper E. lanceolatus. Int. Aquat. Res. 2018, 10, 391–402. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.-S.; Hsu, C.-Y. Ecological considerations of cage aquaculture in Taiwan. J. Fish. Soc. Taiwan 2006, 33, 139–146. [Google Scholar]
- Zhang, Y.; Chu, C.; Li, T.; Xu, S.; Liu, L.; Ju, M. A water quality management strategy for regionally protected water through health risk assessment and spatial distribution of heavy metal pollution in 3 marine reserves. Sci. Total Environ. 2017, 599–600, 721–731. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.-G.; Lin, Q.; Huang, H.-H.; Wang, L.-G.; Ning, J.-J.; Du, F.-Y. Heavy metals in fish tissues/stomach contents in four marine wild commercially valuable fish species from the western continental shelf of South China Sea. Mar. Pollut. Bull. 2017, 114, 1125–1129. [Google Scholar] [CrossRef]
- Gochfeld, M. Cases of mercury exposure, bioavailability, and absorption. Ecotoxicol. Environ. Saf. 2003, 56, 174–179. [Google Scholar] [CrossRef]
- Jinadasa, B.; Jayasinghe, G.; Pohl, P.; Fowler, S.W. Mitigating the impact of mercury contaminants in fish and other seafood—A review. Mar. Pollut. Bull. 2021, 171, 112710. [Google Scholar] [CrossRef]
- Miao, X.; Hao, Y.; Tang, X.; Xie, Z.; Liu, L.; Luo, S.; Huang, Q.; Zou, S.; Zhang, C.; Li, J. Analysis and health risk assessment of toxic and essential elements of the wild fish caught by anglers in Liuzhou as a large industrial city of China. Chemosphere 2020, 243, 125337. [Google Scholar] [CrossRef]
- Zhong, W.; Zhang, Y.; Wu, Z.; Yang, R.; Chen, X.; Yang, J.; Zhu, L. Health risk assessment of heavy metals in freshwater fish in the central and eastern North China. Ecotoxicol. Environ. Saf. 2018, 157, 343–349. [Google Scholar] [CrossRef]
- Raihan, S.M.; Moniruzzaman, M.; Park, Y.; Lee, S.; Bai, S.C. Evaluation of Dietary Organic and Inorganic Mercury Threshold Levels on Induced Mercury Toxicity in a Marine Fish Model. Animals 2020, 10, 405. [Google Scholar] [CrossRef] [Green Version]
- Jang, J.-W.; Lee, S.; Lee, B.-J.; Hur, S.-W.; Son, M.-H.; Kim, K.-W.; Kim, K.-D.; Han, H.-S. A comparative study of effects of dietary mercuric chloride and methylmercury chloride on growth performance, tissue accumulation, stress and immune responses, and plasma measurements in Korean rockfish, Sebastes schlegeli. Chemosphere 2020, 260, 127611. [Google Scholar] [CrossRef] [PubMed]
- Awasthi, A.; Rathore, G.; Pradhan, P.K.; Rebello, S.C.; Khan, M.Y.; Lakra, W.S. Isolation and characterization of head kidney derived macrophages of Labeo rohita. J. Environ. Biol. 2014, 35, 949–954. [Google Scholar] [PubMed]
- Chang, C.S.; Huang, S.L.; Cheng, S.; Cheng, S.N. Innate immune responses and efficacy of using mushroom beta-glucan mixture (MBG) on orange-spotted grouper, Epinephelus coioides, aquaculture. Fish Shellfish Immun. 2013, 35, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Domínguez-Borbor, C.; Chalén-Alvarado, B.; Rodríguez, J.A. A simple in vitro method to evaluate the toxicity of functional additives used in shrimp aquaculture. MethodsX 2018, 5, 90–95. [Google Scholar] [CrossRef] [PubMed]
- Yue, F.; Pan, L.; Xie, P.; Zheng, D.; Li, J. Immune responses and expression of immune-related genes in swimming crab Portunus trituberculatus exposed to elevated ambient ammonia-N stress. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2010, 157, 246–251. [Google Scholar] [CrossRef]
- Rotllant, J.; Parra, D.; Peters, R.; Boshra, H.; Sunyer, J.O. Generation, purification and functional characterization of three C3a anaphylatoxins in rainbow trout: Role in leukocyte chemotaxis and respiratory burst. Dev. Comp. Immunol. 2004, 28, 815–828. [Google Scholar] [CrossRef]
- Xu, X.; Wang, W.-X. Mercury exposure and source tracking in distinct marine-caged fish farm in southern China. Environ. Pollut. 2017, 220, 1138–1146. [Google Scholar] [CrossRef] [PubMed]
- Morcillo, P.; Esteban, M.A.; Cuesta, A. Mercury and its toxic effects on fish. AIMS Environ. Sci. 2017, 4, 386–402. [Google Scholar] [CrossRef]
- Morcillo, P.; Cordero, H.; Meseguer, J.; Esteban, M.; Cuesta, A. In vitro immunotoxicological effects of heavy metals on European sea bass (Dicentrarchus labrax L.) head-kidney leucocytes. Fish Shellfish Immunol. 2015, 47, 245–254. [Google Scholar] [CrossRef]
- Wu, F.; Huang, W.; Liu, Q.; Xu, X.; Zeng, J.; Cao, L.; Hu, J.; Xu, X.; Gao, Y.; Jia, S. Responses of Antioxidant Defense and Immune Gene Expression in Early Life Stages of Large Yellow Croaker (Pseudosciaena crocea) Under Methyl Mercury Exposure. Front. Physiol. 2018, 9, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parisi, M.G.; Pirrera, J.; La Corte, C.; Dara, M.; Parrinello, D.; Cammarata, M. Effects of organic mercury on Mytilus galloprovincialis hemocyte function and morphology. J. Comp. Physiol. B 2021, 191, 143–158. [Google Scholar] [CrossRef] [PubMed]
- Morcillo, P.; Esteban, M.Á.; Cuesta, A. Heavy metals produce toxicity, oxidative stress and apoptosis in the marine teleost fish SAF-1 cell line. Chemosphere 2016, 144, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Tong, J.; Wang, Y.; Lu, Y. In vitro evaluation of inorganic and methyl mercury mediated cytotoxic effect on neural cells derived from different animal species. J. Environ. Sci. 2016, 41, 138–145. [Google Scholar] [CrossRef] [PubMed]
- ISO 10993-5:2009(E); Biological evaluation of medical devices—Part 5: Tests for in vitro cytotoxicity. ISO: Geneva, Switzerland, 2009. Available online: http://nhiso.com/wp-content/uploads/2018/05/ISO-10993-5-2009.pdf (accessed on 17 November 2021).
- Esteban, M.; Cuesta, A.; Chaves-Pozo, E.; Meseguer, J. Phagocytosis in Teleosts. Implications of the New Cells Involved. Biology 2015, 4, 907–922. [Google Scholar] [CrossRef] [Green Version]
- Uribe-Querol, E.; Rosales, C. Phagocytosis. In Reference Module in Biomedical Sciences; Elsevier: Amsterdam, The Netherlands, 2021. [Google Scholar]
- Tort, L. HORMONAL RESPONSES TO STRESS|Impact of Stress in Health and Reproduction. In Encyclopedia of Fish Physiology; Farrell, A.P., Ed.; Academic Press: San Diego, CA, USA, 2011; pp. 1541–1552. [Google Scholar]
- Fournier, M.; Pellerin, J.; Clermont, Y.; Morin, Y.; Brousseau, P. Effects of in vivo exposure of Mya arenaria to organic and inorganic mercury on phagocytic activity of hemocytes. Toxicology 2001, 161, 201–211. [Google Scholar] [CrossRef]
- El-Boshy, M.E.; El-Ashram, A.M.; Abdelhamid, F.M.; Gadalla, H.A. Immunomodulatory effect of dietary Saccharomyces cerevisiae, beta-glucan and laminaran in mercuric chloride treated Nile tilapia (Oreochromis niloticus) and experimentally infected with Aeromonas hydrophila. Fish Shellfish Immunol. 2010, 28, 802–808. [Google Scholar] [CrossRef]
- Łuczyńska, J.; Paszczyk, B.; Łuczyński, M.J. Fish as a bioindicator of heavy metals pollution in aquatic ecosystem of Pluszne Lake, Poland, and risk assessment for consumer’s health. Ecotoxicol. Environ. Saf. 2018, 153, 60–67. [Google Scholar] [CrossRef]
- Järv, L.; Kotta, J.; Simm, M. Relationship between biological characteristics of fish and their contamination with trace metals: A case study of perch Perca fluviatilis L. in the Baltic Sea. Proc. Est. Acad. Sci. 2013, 62, 193–201. [Google Scholar] [CrossRef]
- Huang, S.S.-Y.; Strathe, A.B.; Fadel, J.G.; Lin, P.; Liu, T.-Y.; Hung, S.S.O. Absorption, distribution, and elimination of graded oral doses of methylmercury in juvenile white sturgeon. Aquat. Toxicol. 2012, 122–123, 163–171. [Google Scholar] [CrossRef]
- Garnero, P.L.; Monferran, M.V.; González, G.A.; Griboff, J.; de los Ángeles, B.M. Assessment of exposure to metals, As and Se in water and sediment of a freshwater reservoir and their bioaccumulation in fish species of different feeding and habitat preferences. Ecotoxicol. Environ. Saf. 2018, 163, 492–501. [Google Scholar] [CrossRef] [PubMed]
- Ikemoto, T.; Kunito, T.; Tanaka, H.; Baba., N.; Miyazaki., N.; Tanabe., S. Detoxification mechanism of heavy metals in marine mammals and seabirds: Interaction of selenium with mercury, silver, copper, zinc, and cadmium in liver. Arch Environ. Contam. Toxicol. 2004, 47, 402–413. [Google Scholar] [CrossRef]
- Ayyat, M.S.; Ayyat, A.M.N.; Abd El-Latif, K.M.; Hessein, A.A.A.; Al-Sagheer, A.A. Inorganic mercury and dietary safe feed additives enriched diet impacts on growth, immunity, tissue bioaccumulation, and disease resistance in Nile tilapia (Oreochromis niloticus). Aquat. Toxicol. 2020, 224, 105494. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dose (mg/kg) | Time Elapsed (Day) | ||||||
|---|---|---|---|---|---|---|---|
| 0 | 1 | 4 | 7 | 14 | 21 | 28 | |
| Control | 40.56 ± 3.10 B | 33.73 ± 2.75 bCD | 25.40 ± 1.07 bD | 47.37 ± 3.09 bA | 33.17 ± 1.89 cCD | 28.37 ± 0.05 cD | 39.83 ± 0.83 dBC |
| 1 mg/kg | 38.57 ± 2.25 bB | 31.90 ± 3.09 abB | 76.57 ± 9.90 abB | 71.40 ± 6.14 bB | 71.50 ± 6.33 bB | 224.60 ± 33.87 cA | |
| 5 mg/kg | 33.13 ± 1.76 bB | 50.00 ± 5.83 aB | 89.17 ± 6.97 abB | 116.53 ± 2.91 aB | 154.60 ± 32.74 abB | 427.50 ± 94.02 bA | |
| 10 mg/kg | 42.07 ± 2.46 aB | 46.27 ± 9.71 aB | 130.27 ± 34.72 aB | 130.07 ± 14.08 aB | 177.80 ± 72.07 aB | 1213.17 ± 192.08 aA | |
| Dose (mg/kg) | Time Elapsed (Day) | ||||||
|---|---|---|---|---|---|---|---|
| 0 | 1 | 4 | 7 | 14 | 21 | 28 | |
| Control | 19.4 ± 0.75 B | 28.00 ± 1.02 aB | 24.43 ± 5.83 aB | 34.23 ± 0.68 bA | 18.77 ± 4.46 cB | 26.13 ± 2.19 cB | 29.57 ± 1.23 dB |
| 1 mg/kg | 33.30 ± 1.40 aC | 31.63 ± 9.97 aC | 133.13 ± 32.26 abB | 159.63 ± 18.35 bB | 218.57 ± 11.48 bB | 478.73 ± 51.69 cA | |
| 5 mg/kg | 30.23 ± 1.52 aC | 53.83 ± 26.44 aC | 134.63 ± 16.41 abC | 408.00 ± 43.29 aB | 374.63 ± 76.46 abB | 1136.67 ± 91.59 bA | |
| 10 mg/kg | 51.63 ± 20.74 aB | 93.53 ± 38.30 aB | 249.50 ± 97.37 aB | 458.77 ± 90.92 aB | 331.67 ± 44.73 aB | 3428.0 ± 346.34 aA | |
| Dose (mg/kg) | Time Elapsed (Day) | ||||||
|---|---|---|---|---|---|---|---|
| 0 | 1 | 4 | 7 | 14 | 21 | 28 | |
| Control | 18.77 ± 3.51 A | 10.83 ± 0.54 aB | 6.17 ± 1.05 cB | 22.93 ± 0.40 bA | 5.43 ± 1.25 cB | 10.60 ± 0.28 cB | 17.87 ± 1.51 dA |
| 1 mg/kg | 13.23 ± 2.41 aC | 23.87 ± 6.33 bC | 128.77 ± 56.34 aB | 149.33 ± 19.01 bB | 211.63 ± 16.16 bB | 433.97 ± 14.48 cA | |
| 5 mg/kg | 21.10 ± 1.61 aD | 56.93 ± 13.70 aCD | 189.13 ± 35.01 aBCD | 266.00 ± 29.12 aBC | 336.57 ± 50.95 aB | 1063.67 ± 159.79 bA | |
| 10 mg/kg | 36.30 ± 19.04 aB | 103.87 ± 59.37 aB | 222.70 ± 79.69 aB | 316.73 ± 53.88 aB | 252.93 ± 75.78 aB | 4037.67 ± 287.47 aA | |
| Tissue | Dose (mg/kg) | Time Elapsed (Day) | ||||||
|---|---|---|---|---|---|---|---|---|
| 0 | 1 | 4 | 7 | 14 | 21 | 28 | ||
| Muscle | Control | 40.56 ± 3.10 a | 33.73 ± 2.75 abc | 25.40 ± 1.07 ab | 47.37 ± 3.09 cd | 33.17 ± 1.89 ef | 28.37 ± 0.05 fg | 39.83 ± 0.83 d |
| 1 mg/kg | 38.57 ± 2.25 abc | 31.90 ± 3.09 ab | 76.57 ± 9.90 bcd | 71.40 ± 6.14 ef | 71.50 ± 6.33 efg | 224.60 ± 33.87 d | ||
| 5 mg/kg | 33.13 ± 1.76 abc | 50.00 ± 5.83 ab | 89.17 ± 6.97 bcd | 116.53 ± 2.91 ef | 154.60 ± 32.74 defg | 427.50 ± 94.02 d | ||
| 10 mg/kg | 42.07 ± 2.46 ab | 46.27 ± 9.71 ab | 130.27 ± 34.72 abcd | 130.07 ± 14.08 ef | 177.80 ± 72.07 cdef | 1213.17 ± 192.08 c | ||
| Liver | Control | 19.4 ± 0.75b | 28.00 ± 1.02 abc | 24.43 ± 5.83 ab | 34.23 ± 0.68 cd | 18.77 ± 4.46 f | 26.13 ± 2.19 fg | 29.57 ± 1.23 d |
| 1 mg/kg | 33.30 ± 1.40 abc | 31.63 ± 9.97 ab | 133.13 ± 32.26 abcd | 159.63 ± 18.35 de | 218.57 ± 11.48 abcde | 478.73 ± 51.69 d | ||
| 5 mg/kg | 30.23 ± 1.52 abc | 53.83 ± 26.44 ab | 134.63 ± 16.41 abcd | 408.00 ± 43.29 ab | 374.63 ± 76.46 a | 1136.67 ± 91.59 c | ||
| 10 mg/kg | 51.63 ± 20.74 a | 93.53 ± 38.30 a | 249.50 ± 97.37 a | 458.77 ± 90.92 a | 331.67 ± 44.73 abc | 3428.00 ± 346.34 b | ||
| Kidney | Control | 18.77 ± 3.51b | 10.83 ± 0.54 c | 6.17 ± 1.05 b | 22.93 ± 0.40 d | 5.43 ± 1.25 f | 10.60 ± 0.28 g | 17.87 ± 1.51 d |
| 1 mg/kg | 13.23 ± 2.41 bc | 23.87 ± 6.33 ab | 128.77 ± 56.34 abcd | 149.33 ± 19.01 de | 211.63 ± 16.16 bcde | 433.97 ± 14.48 d | ||
| 5 mg/kg | 21.10 ± 1.61 bc | 56.93 ± 13.70 ab | 189.13 ± 35.01 abcd | 266.00 ± 29.12 cd | 336.57 ± 50.95 ab | 1063.67 ± 159.79 c | ||
| 10 mg/kg | 36.30 ± 19.04 abc | 103.87 ± 59.37 a | 222.70 ± 79.69 ab | 316.73 ± 53.88 bc | 252.93 ± 75.78 abcd | 4037.67 ± 287.47 a | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chuang, H.-C.; Huang, H.-T.; Dewi, N.-R.; Hsiao, H.-H.; Chen, B.-Y.; Liao, Z.-H.; Lee, M.-C.; Lee, P.-T.; Wu, Y.-S.; Lin, Y.-J.; et al. Effect of Methylmercury Exposure on Bioaccumulation and Nonspecific Immune Respsonses in Hybrid Grouper Epinephelus fuscoguttatus × Epinephelus lanceolatus. Animals 2022, 12, 147. https://doi.org/10.3390/ani12020147
Chuang H-C, Huang H-T, Dewi N-R, Hsiao H-H, Chen B-Y, Liao Z-H, Lee M-C, Lee P-T, Wu Y-S, Lin Y-J, et al. Effect of Methylmercury Exposure on Bioaccumulation and Nonspecific Immune Respsonses in Hybrid Grouper Epinephelus fuscoguttatus × Epinephelus lanceolatus. Animals. 2022; 12(2):147. https://doi.org/10.3390/ani12020147
Chicago/Turabian StyleChuang, Hsiang-Chieh, Huai-Ting Huang, Novi-Rosmala Dewi, Hsi-Hua Hsiao, Bo-Ying Chen, Zhen-Hao Liao, Meng-Chou Lee, Po-Tsang Lee, Yu-Sheng Wu, Yu-Ju Lin, and et al. 2022. "Effect of Methylmercury Exposure on Bioaccumulation and Nonspecific Immune Respsonses in Hybrid Grouper Epinephelus fuscoguttatus × Epinephelus lanceolatus" Animals 12, no. 2: 147. https://doi.org/10.3390/ani12020147