

Visual and Rapid Detection of Porcine Epidemic Diarrhea Virus (PEDV) Using Reverse Transcription Loop-Mediated Isothermal Amplification Method
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Viral Materials
2.2. DNA and RNA Extraction
2.3. Primer Design
2.4. RT-PCR
2.5. RT-LAMP
2.6. Temperature Optimization
2.7. Specificity of RT-LAMP Method
2.8. Comparison of the Sensitivity between RT-LAMP and RT-PCR
2.9. Analysis of RT-LAMP and RT-PCR Products
2.10. Detection of Clinical Specimens
3. Results
3.1. Temperature Optimization of the PEDV RT-LAMP Method
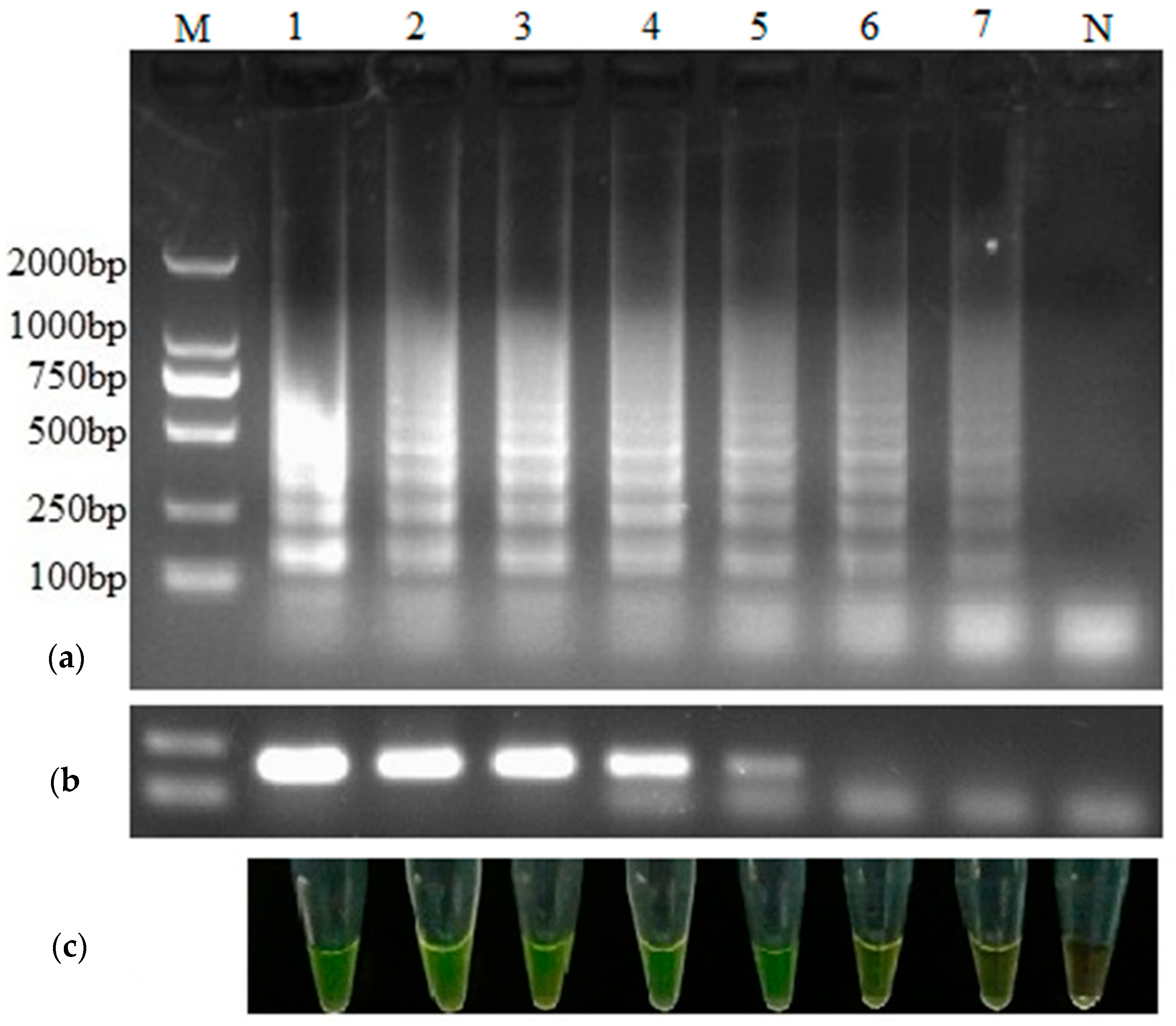
3.2. Specificity of RT-LAMP Method
3.3. Comparison of the Sensitivity between RT-LAMP and RT-PCR
3.4. Detection of Clinical Specimens
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Collin, E.A.; Anbalagan, S.; Okda, F.; Batman, R.; Nelson, E.; Hause, B.M. An inactivated vaccine made from a U.S. field isolate of porcine epidemic disease virus is immunogenic in pigs as demonstrated by a dose-titration. BMC Vet. Res. 2015, 11, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makadiya, N.; Brownlie, R.; van den Hurk, J.; Berube, N.; Allan, B.; Gerdts, V.; Zakhartchouk, A. S1 domain of the porcine epidemic diarrhea virus spike protein as a vaccine antigen. Virol. J. 2016, 13, 57. [Google Scholar] [CrossRef] [Green Version]
- Song, D.; Moon, H.; Kang, B. Porcine epidemic diarrhea: A review of current epidemiology and available vaccines. Clin. Exp. Vaccine Res. 2015, 4, 166–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, K.; Saif, L.J.; Wang, Q. Porcine epidemic diarrhea virus (PEDV): An update on etiology, transmission, pathogenesis, and prevention and control. Virus Res. 2020, 286, 198045. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Wang, H.Y. Porcine enteric coronaviruses: An updated overview of the pathogenesis, prevalence, and diagnosis. Vet. Res. Commun. 2021, 45, 75–86. [Google Scholar] [CrossRef] [PubMed]
- Pensaert, M.B.; de Bouck, P. A new coronavirus-like particle associated with diarrhea in swine. Arch. Virol. 1978, 58, 243–247. [Google Scholar] [CrossRef] [Green Version]
- Kweon, C.H.; Lee, J.G.; Han, M.G.; Kang, Y.B. Rapid diagnosis of porcine epidemic diarrhea virus infection by polymerase chain reaction. J. Vet. Med. Sci. 1997, 59, 231–232. [Google Scholar] [CrossRef] [Green Version]
- Kusanagi, K.; Kuwahara, H.; Katoh, T.; Nunoya, T.; Ishikawa, Y.; Samejima, T.; Tajima, M. Isolation and serial propagation of porcine epidemic diarrhea virus in cell cultures and partial characterization of the isolate. J. Vet. Med. Sci. 1992, 54, 313–318. [Google Scholar] [CrossRef] [Green Version]
- Stott, C.J.; Temeeyasen, G.; Tripipat, T.; Kaewprommal, P.; Tantituvanont, A.; Piriyapongsa, J.; Nilubol, D. Evolutionary and epidemiological analyses based on spike genes of porcine epidemic diarrhea virus circulating in Thailand in 2008–2015. Infect. Genet. Evol. 2017, 50, 70–76. [Google Scholar] [CrossRef]
- Chen, J.; Wang, C.; Shi, H.; Qiu, H.; Liu, S.; Chen, X.; Zhang, Z.; Feng, L. Molecular epidemiology of porcine epidemic diarrhea virus in China. Arch. Virol. 2010, 155, 1471–1476. [Google Scholar] [CrossRef]
- Li, W.; Li, H.; Liu, Y.; Pan, Y.; Deng, F.; Song, Y.; Tang, X.; He, Q. New variants of porcine epidemic diarrhea virus, China, 2011. Emerg. Infect. Dis. 2012, 18, 1350–1353. [Google Scholar] [CrossRef] [PubMed]
- Vlasova, A.N.; Marthaler, D.; Wang, Q.; Culhane, M.R.; Rossow, K.D.; Rovira, A.; Collins, J.; Saif, L.J. Distinct characteristics and complex evolution of PEDV strains, North America, May 2013-February 2014. Emerg. Infect. Dis. 2014, 20, 1620–1628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carvajal, A.; Lanza, I.; Diego, R.; Rubio, P.; Cármenes, P. Evaluation of a blocking ELISA using monoclonal antibodies for the detection of porcine epidemic diarrhea virus and its antibodies. J. Vet. Diagn. Investig. 1995, 7, 60–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, X.L.; Yu, L.Y.; Liu, J. Development and evaluation of enzyme-linked immunosorbent assay based on recombinant nucleocapsid protein for detection of porcine epidemic diarrhea (PEDV) antibodies. Vet. Microbiol. 2007, 123, 86–92. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.; Chae, C. RT-PCR-based dot blot hybridization for the detection and differentiation between porcine epidemic diarrhea virus and transmissible gastroenteritis virus in fecal samples using a non-radioactive digoxigenin cDNA probe. J. Virol. Methods 2005, 123, 141–146. [Google Scholar] [CrossRef]
- Kim, S.H.; Kim, I.J.; Pyo, H.M.; Tark, D.S.; Song, J.Y.; Hyun, B.H. Multiplex real-time RT-PCR for the simultaneous detection and quantification of transmissible gastroenteritis virus and porcine epidemic diarrhea virus. J. Virol. Methods 2007, 146, 172–177. [Google Scholar] [CrossRef]
- Song, D.S.; Kang, B.K.; Oh, J.S.; Ha, G.W.; Yang, J.S.; Moon, H.J.; Jang, Y.S.; Park, B.K. Multiplex reverse transcription-PCR for rapid differential detection of porcine epidemic diarrhea virus, transmissible gastroenteritis virus, and porcine group A rotavirus. J. Vet. Diagn. Investig. 2006, 18, 278–281. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Zhang, T.; Song, D.; Huang, T.; Peng, Q.; Chen, Y.; Li, A.; Zhang, F.; Wu, Q.; Ye, Y.; et al. Comparison and evaluation of conventional RT-PCR, SYBR green I and TaqMan real-time RT-PCR assays for the detection of porcine epidemic diarrhea virus. Mol. Cell Probes. 2017, 33, 36–41. [Google Scholar] [CrossRef]
- Ihira, M.; Yoshikawa, T.; Enomoto, Y.; Akimoto, S.; Ohashi, M.; Suga, S.; Nishimura, N.; Ozaki, T.; Nishiyama, Y.; Notomi, T.; et al. Rapid diagnosis of human herpesvirus 6 infection by a novel DNA amplification method, loop-mediated isothermal amplification. J. Clin. Micro. Biol. 2004, 42, 140–145. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.F.; Cui, S.J.; Zhu, C. Loop-mediated isothermal amplification for rapid detection and differentiation of wild-type pseudorabies and gene-deleted virus vaccines. J. Virol. Methods 2010, 169, 239–243. [Google Scholar] [CrossRef]
- Iwamoto, T.; Sonobe, T.; Hayashi, K. Loop-mediated isothermal amplification for direct detection of Mycobacterium tuberculosis complex, M. avium, and M. intracellulare in sputum samples. J. Clin. Microbiol. 2003, 41, 2616–2622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, W.; Wang, J.; Sun, M.; Zheng, Y.; Li, L.; Zhang, X.; Sun, J. Rapid detection of encephalomyocarditis virus by one-step reverse transcription loop-mediated isothermal amplification method. Virus Res. 2014, 189, 75–78. [Google Scholar] [CrossRef] [PubMed]
- Fujino, M.; Yoshida, N.; Yamaguchi, S.; Hosaka, N.; Ota, Y.; Notomi, T.; Nakayama, T. A simple method for the detection of measles virus genome by loop-mediated isothermal amplification (LAMP). J. Med. Virol. 2005, 76, 406–413. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; Shi, W.; Han, F.; Xu, Y.; Zhu, L.; Zou, Y.; Wu, X.; Zhu, H.; Tan, F.; Tao, S.; et al. Specific, simple and rapid detection of porcine circovirus type 2 using the loop-mediated isothermal amplification method. Virol. J. 2011, 8, 126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Postel, A.; Letzel, T.; Frischmann, S.; Grund, C.; Beer, M.; Harder, T. Evaluation of two commercial loop-mediated isothermal amplification assays for detection of avian influenza H5 and H7 hemagglutinin genes. J. Vet. Diagn. Investig. 2010, 22, 61–66. [Google Scholar] [CrossRef] [Green Version]
- Gou, H.; Deng, J.; Pei, J.; Wang, J.; Liu, W.; Zhao, M.; Chen, J. Rapid and sensitive detection of type II porcine reproductive and respiratory syndrome virus by reverse transcription loop-mediated isothermal amplification combined with a vertical flow visualization strip. J. Virol. Methods 2014, 209, 86–94. [Google Scholar] [CrossRef]
- Dukes, J.P.; King, D.P.; Alexandersen, S. Novel reverse transcription loop-mediated isothermal amplification for rapid detection of foot-and-mouth disease virus. Arch. Virol. 2006, 151, 1093–1106. [Google Scholar] [CrossRef]
- Ren, X.; Li, P. Development of reverse transcription loop-mediated isothermal amplification for rapid detection of porcine epidemic diarrhea virus. Virus Genes. 2011, 42, 229–235. [Google Scholar] [CrossRef]
- Sun, Q.W.; Wu, S.Q.; Yang, C.J.; Wang, J.L.; Ge, J.; Chang, B.; Xu, T. Development of a reverse transcription loop-mediated isothermal amplification assay for visual detection of porcine epidemic diarrhea virus. J. Anim. Vet. Adv. 2012, 11, 1897–1900. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Shi, L.; Lv, X.; Yao, W.; Cao, M.; Yu, H.; Wang, X.; Zheng, S. Development of a real-time reverse transcription loop-mediated isothermal amplification method for the rapid detection of porcine epidemic diarrhea virus. Virol. J. 2015, 12, 76. [Google Scholar] [CrossRef]
- Gou, H.; Deng, J.; Wang, J.; Pei, J.; Liu, W.; Zhao, M.; Chen, J. Rapid and sensitive detection of porcine epidemic diarrhea virus by reverse transcription loop-mediated isothermal amplification combined with a vertical flow visualization strip. Mol. Cell Probes. 2015, 29, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Mai, T.N.; Nguyen, V.D.; Yamazaki, W.; Okabayashi, T.; Mitoma, S.; Notsu, K.; Sakai, Y.; Yamaguchi, R.; Norimine, J.; Sekiguchi, S. Development of pooled testing system for porcine epidemic diarrhoea using real-time fluorescent reversetranscription loop-mediated isothermal amplification assay. BMC Vet. Res. 2018, 14, 172. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.K.; Kim, H.R.; Kim, D.Y.; Kim, J.M.; Kwon, N.Y.; Park, J.H.; Park, J.Y.; Kim, S.H.; Lee, K.K.; Lee, C.; et al. A simple colorimetric detection of porcine epidemic diarrhea virus by reverse transcription loop-mediated isothermal amplification assay using hydroxynaphthol blue metal indicator. J. Virol. Methods. 2021, 298, 114289. [Google Scholar] [CrossRef] [PubMed]
- Kocherhans, R.; Bridgen, A.; Ackermann, M.; Tobler, K. Completion of the porcine epidemic diarrhoea coronavirus (PEDV) genome sequence. Virus Genes. 2001, 23, 137–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.; Li, Z.; Zou, Y.; Wicht, O.; van Kuppeveld, F.J.M.; Rottier, P.J.M.; Bosch, B.J. Manipulation of the porcine epidemic diarrhea virus genome using targeted RNA recombination. PLoS ONE. 2013, 8, e69997. [Google Scholar] [CrossRef] [Green Version]
- Notomi, T.; Okayama, H.; Masubuchi, H.; Yonekawa, T.; Watanabe, K.; Amino, N.; Hase, T. Loop-mediated isothermal amplification of DNA. Nucleic Acids Res. 2000, 28, E63. [Google Scholar] [CrossRef] [Green Version]
- Nagamine, K.; Hase, T.; Notomi, T. Accelerated reaction by loop-mediated isothermal amplification using loop primers. Mol. Cell. Probes. 2002, 16, 223–229. [Google Scholar] [CrossRef]
- Wang, Z.; Yin, J.; Chen, S.; Qiao, X.; Tang, L.; Li, Y. Development of an antigen capture enzyme-linked immunosorbent assay for virus detection based on porcine epidemic diarrhea virus monoclonal antibodies. Viral Immunol. 2015, 28, 184–189. [Google Scholar] [CrossRef] [Green Version]
- Sozzi, E.; Luppi, A.; Lelli, D.; Martin, A.M.; Canelli, E.; Brocchi, E.; Lavazza, A.; Cordioli, P. Comparison of enzyme-linked immunosorbent assay and RT-PCR for the detection of porcine epidemic diarrhoea virus. Res. Vet. Sci. 2010, 88, 166–168. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Primer | Type | Position (nt) 1 | Sequence (5′-3′) |
|---|---|---|---|
| PEDV F3 | forward outer primer | 842–859 | GGAGGAGAATTCCCAAGG |
| PEDV B3 | reverse outer primer | 1022–1039 | AAGAGTCCGCTAGCTCAC |
| PEDV FIP | forward inner primer (F2-F1c) | F1c: 900–924 F2: 860–877 | TTCCGCATCTCCAAAATTTTTGAAG GCGAAAATAGCGTAGCAG |
| PEDV BIP | reverse inner primer (B1c-B2) | B1c: 939–959 B2: 1002–1019 | TGTTGATGCCTCAGGCTATGC ACAGCCACATTACCACCA |
| PEDV LF | Loop forward primer | 879–895 | CCCTGGGTCCGAAGCAA |
| PEDV LB | Loop outer primer | 975–993 | AGCACCAAATGTTGCAGCA |
| RT-LAMP | RT-PCR | Coincidence Rate | ||
|---|---|---|---|---|
| + | − | |||
| + | 57 | 54 | 3 | 96.1% (99/103) |
| − | 46 | 1 | 45 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, C.; Liang, J.; Yang, D.; Zhang, Q.; Miao, D.; He, X.; Du, Y.; Zhang, W.; Ni, J.; Zhao, K. Visual and Rapid Detection of Porcine Epidemic Diarrhea Virus (PEDV) Using Reverse Transcription Loop-Mediated Isothermal Amplification Method. Animals 2022, 12, 2712. https://doi.org/10.3390/ani12192712
Li C, Liang J, Yang D, Zhang Q, Miao D, He X, Du Y, Zhang W, Ni J, Zhao K. Visual and Rapid Detection of Porcine Epidemic Diarrhea Virus (PEDV) Using Reverse Transcription Loop-Mediated Isothermal Amplification Method. Animals. 2022; 12(19):2712. https://doi.org/10.3390/ani12192712
Chicago/Turabian StyleLi, Chunhua, Jieling Liang, Dan Yang, Qi Zhang, Denian Miao, Xizhong He, Yanan Du, Wanjing Zhang, Jianping Ni, and Kai Zhao. 2022. "Visual and Rapid Detection of Porcine Epidemic Diarrhea Virus (PEDV) Using Reverse Transcription Loop-Mediated Isothermal Amplification Method" Animals 12, no. 19: 2712. https://doi.org/10.3390/ani12192712