Identification and Characterization of lncRNAs Expression Profile Related to Goat Skeletal Muscle at Different Development Stages


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Preparation and Sample Collection
2.2. RNA Isolation, LncRNA-Seq Library Construction, and Sequencing
2.3. Prediction of Multi-Exon LncRNAs
2.4. Classification and Differential Expression Analysis
2.5. Co-Location and Co-Expression Analysis, and Functional Annotation Analysis
2.6. Verification of LncRNA Expression Pattern with RT-qPCR
2.7. Construction of the CeRNA (lncRNA-miRNA-mRNA) Network
2.8. Statistical Analysis
3. Results
3.1. RNA-Seq Data Filtering, Mapping, and Transcript Assembly
3.2. Identification and Confirmation of LncRNAs in Goat Longissimus Dorsi Tissue
3.3. Differential Expression Analysis
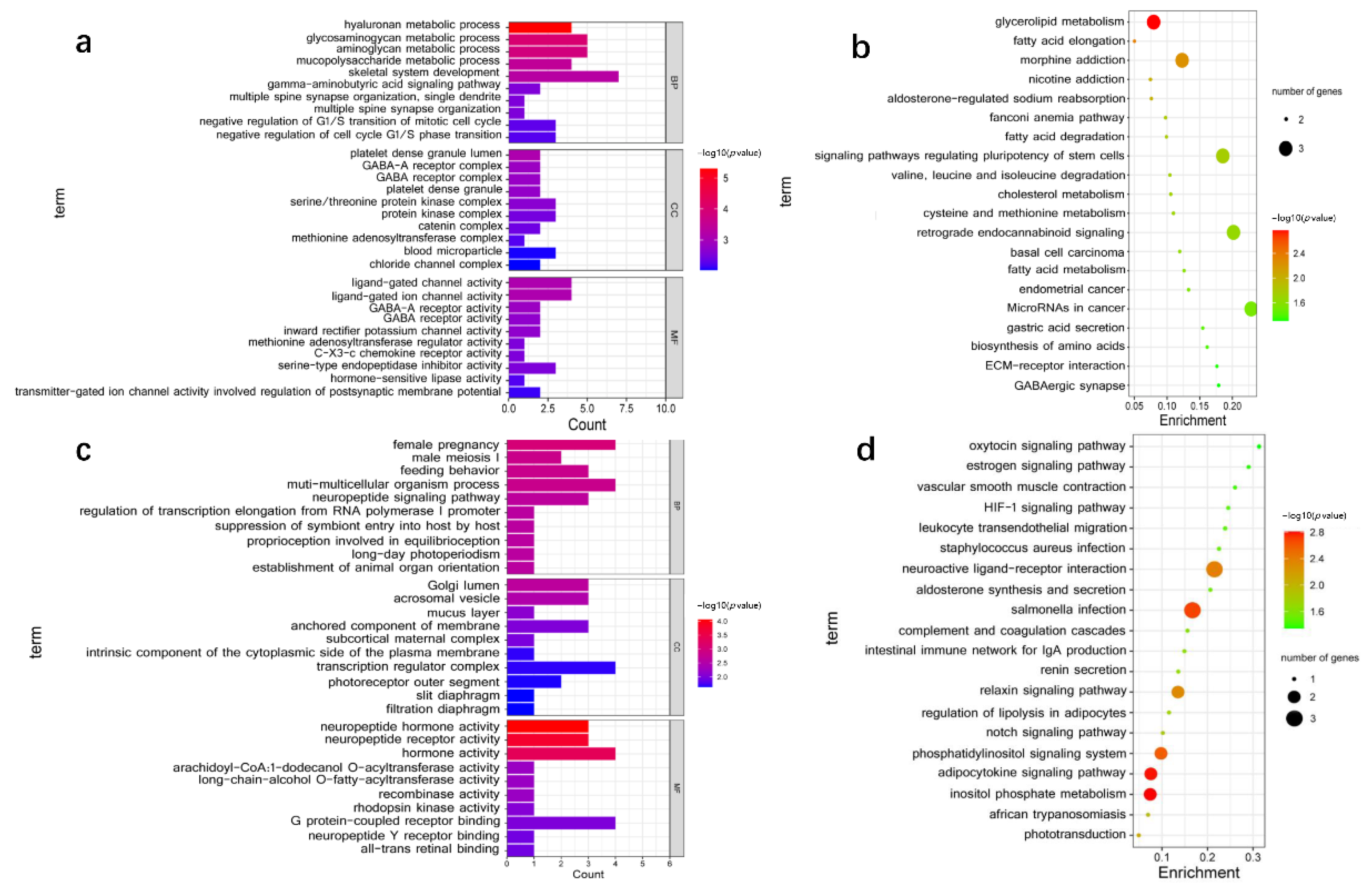
3.4. Functional Enrichment Analysis
3.5. Verification of DE LncRNAs Expression Profile with RT-qPCR
3.6. Construction and Bioinformatics Analysis of the ceRNA Network
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zeng, C.; Shi, H.; Kirkpatrick, L.T.; Ricome, A.; Park, S.; Scheffler, J.M.; Hannon, K.M.; Grant, A.L.; Gerrard, D.E. Driving an Oxidative Phenotype Protects Myh4 Null Mice From Myofiber Loss During Postnatal Growth. Front. Physiol. 2021, 12, 785151. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Choi, Y.M.; Lee, J.; Shin, S.; Kim, S.; Suh, Y.; Lee, K. Differential Expression of MSTN Isoforms in Muscle between Broiler and Layer Chickens. Animals 2022, 12, 539. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.G.; Yuan, Y.; Zhou, D.K.; Ma, Y.H.; Mahrous, K.F.; Wang, S.Z.; He, Y.M.; Duan, X.H.; Zhang, W.Y.; E, G. Genome-wide selection signal analysis of Australian Boer goat reveals artificial selection imprinting on candidate genes related to muscle development. Anim. Genet. 2021, 52, 550–555. [Google Scholar] [CrossRef]
- Shen, J.; Hao, Z.; Wang, J.; Hu, J.; Liu, X.; Li, S.; Ke, N.; Song, Y.; Lu, Y.; Hu, L.; et al. Comparative Transcriptome Profile Analysis of Longissimus dorsi Muscle Tissues From Two Goat Breeds with Different Meat Production Performance Using RNA-Seq. Front. Genet. 2020, 11, 619399. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Zhang, J.; Zhu, J.Q.; Gu, S.; Sun, Q.; Zhou, G.M.; Fu, C.X.; Li, Q.; Chen, L.Y.; Li, D.X.; et al. Genetic diversity of nine populations of the black goat (Capra hircus) in Sichuan, PR China. Zoolog. Sci. 2006, 23, 229–234. [Google Scholar] [CrossRef]
- Liu, K.; Jin, Y.; Zhang, F.Y.; Zhang, Y.Z.; Quan, X.Q.; Han, Q.D.; Liu, L.X.; Liu, Y.G.; Lv, S.J.; Qu, X.M. The complete mitochondrial DNA sequence of Yimeng black goat (Capra hircus) and its potential application in mutton discrimination. Mitochondrial DNA B Resour. 2020, 5, 2303–2305. [Google Scholar] [CrossRef] [PubMed]
- Prensner, J.R.; Chinnaiyan, A.M. The emergence of lncRNAs in cancer biology. Cancer Discov. 2011, 1, 391–407. [Google Scholar] [CrossRef] [Green Version]
- Ming, G.F.; Wu, K.; Hu, K.; Chen, Y.; Xiao, J. NAMPT regulates senescence, proliferation, and migration of endothelial progenitor cells through the SIRT1 AS lncRNA/miR-22/SIRT1 pathway. Biochem. Biophys. Res. Commun. 2016, 478, 1382–1388. [Google Scholar] [CrossRef]
- Shi, X.; Sun, M.; Liu, H.; Yao, Y.; Song, Y. Long non-coding RNAs: A new frontier in the study of human diseases. Cancer Lett. 2013, 339, 159–166. [Google Scholar] [CrossRef]
- Sendler, E.; Johnson, G.D.; Mao, S.; Goodrich, R.J.; Diamond, M.P.; Hauser, R.; Krawetz, S.A. Stability, delivery and functions of human sperm RNAs at fertilization. Nucleic Acids Res. 2013, 41, 4104–4117. [Google Scholar] [CrossRef]
- Ma, L.; Bajic, V.B.; Zhang, Z. On the classification of long non-coding RNAs. RNA Biol. 2013, 10, 925–933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Li, X. The role of noncoding RNA in hepatocellular carcinoma. Gland Surg. 2013, 2, 25–29. [Google Scholar] [PubMed]
- Bonasio, R.; Shiekhattar, R. Regulation of transcription by long noncoding RNAs. Annu. Rev. Genet. 2014, 48, 433–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morlando, M.; Ballarino, M.; Fatica, A.; Bozzoni, I. The role of long noncoding RNAs in the epigenetic control of gene expression. ChemMedChem 2014, 9, 505–510. [Google Scholar] [CrossRef]
- Mathieu, E.L.; Belhocine, M.; Dao, L.T.; Puthier, D.; Spicuglia, S. Functions of lncRNA in development and diseases. Med. Sci. 2014, 30, 790–796. [Google Scholar]
- Zhan, S.; Qin, C.; Li, D.; Zhao, W.; Nie, L.; Cao, J.; Guo, J.; Zhong, T.; Wang, L.; Li, L.; et al. A Novel Long Noncoding RNA, lncR-125b, Promotes the Differentiation of Goat Skeletal Muscle Satellite Cells by Sponging miR-125b. Front. Genet. 2019, 10, 1171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, C.; Wu, M.; Wang, C.; Liu, R.; Zhao, H.; Yang, L.; Liu, J.; Wang, Y.; Zhang, S.; Yuan, Z.; et al. Long Noncoding RNA Lnc-SEMT Modulates IGF2 Expression by Sponging miR-125b to Promote Sheep Muscle Development and Growth. Cell. Physiol. Biochem. 2018, 49, 447–462. [Google Scholar] [CrossRef]
- Ma, X.; Fu, D.; Chu, M.; Ding, X.; Wu, X.; Guo, X.; Kalwar, Q.; Pei, J.; Bao, P.; Liang, C.; et al. Genome-Wide Analysis Reveals Changes in Polled Yak Long Non-coding RNAs in Skeletal Muscle Development. Front. Genet. 2020, 11, 365. [Google Scholar] [CrossRef]
- Ren, C.; Deng, M.; Fan, Y.; Yang, H.; Zhang, G.; Feng, X.; Li, F.; Wang, D.; Wang, F.; Zhang, Y. Genome-Wide Analysis Reveals Extensive Changes in LncRNAs during Skeletal Muscle Development in Hu Sheep. Genes 2017, 8, 191. [Google Scholar] [CrossRef] [Green Version]
- Yuan, C.; Zhang, K.; Yue, Y.; Guo, T.; Liu, J.; Niu, C.; Sun, X.; Feng, R.; Wang, X.; Yang, B. Analysis of dynamic and widespread lncRNA and miRNA expression in fetal sheep skeletal muscle. PeerJ 2020, 8, e9957. [Google Scholar] [CrossRef]
- Li, Q.; Liu, R.; Zhao, H.; Di, R.; Lu, Z.; Liu, E.; Wang, Y.; Chu, M.; Wei, C. Identification and Characterization of Long Noncoding RNAs in Ovine Skeletal Muscle. Animals 2018, 8, 127. [Google Scholar] [CrossRef] [PubMed]
- Ling, Y.; Zheng, Q.; Sui, M.; Zhu, L.; Xu, L.; Zhang, Y.; Liu, Y.; Fang, F.; Chu, M.; Ma, Y.; et al. Comprehensive Analysis of LncRNA Reveals the Temporal-Specific Module of Goat Skeletal Muscle Development. Int. J. Mol. Sci. 2019, 20, 3950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhan, S.; Dong, Y.; Zhao, W.; Guo, J.; Zhong, T.; Wang, L.; Li, L.; Zhang, H. Genome-wide identification and characterization of long non-coding RNAs in developmental skeletal muscle of fetal goat. BMC Genom. 2016, 17, 666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ling, Y.; Zheng, Q.; Jing, J.; Sui, M.; Zhu, L.; Li, Y.; Zhang, Y.; Liu, Y.; Fang, F.; Zhang, X. RNA-Seq Reveals miRNA Role Shifts in Seven Stages of Skeletal Muscles in Goat Fetuses and Kids. Front. Genet. 2020, 11, 684. [Google Scholar] [CrossRef]
- Kong, L.; Zhang, Y.; Ye, Z.Q.; Liu, X.Q.; Zhao, S.Q.; Wei, L.; Gao, G. CPC: Assess the protein-coding potential of transcripts using sequence features and support vector machine. Nucleic Acids Res. 2007, 35, W345–W349. [Google Scholar] [CrossRef]
- Sun, L.; Luo, H.; Bu, D.; Zhao, G.; Yu, K.; Zhang, C.; Liu, Y.; Chen, R.; Zhao, Y. Utilizing sequence intrinsic composition to classify protein-coding and long non-coding transcripts. Nucleic Acids Res. 2013, 41, e166. [Google Scholar] [CrossRef]
- Sonnhammer, E.L.; Eddy, S.R.; Birney, E.; Bateman, A.; Durbin, R. Pfam: Multiple sequence alignments and HMM-profiles of protein domains. Nucleic Acids Res. 1998, 26, 320–322. [Google Scholar] [CrossRef] [Green Version]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef] [Green Version]
- Han, L.; Zhang, K.; Shi, Z.; Zhang, J.; Zhu, J.; Zhu, S.; Zhang, A.; Jia, Z.; Wang, G.; Yu, S.; et al. LncRNA pro fi le of glioblastoma reveals the potential role of lncRNAs in contributing to glioblastoma pathogenesis. Int. J. Oncol. 2012, 40, 2004–2012. [Google Scholar]
- Liu, Y.; Qi, B.; Xie, J.; Wu, X.; Ling, Y.; Cao, X.; Kong, F.; Xin, J.; Jiang, X.; Wu, Q.; et al. Filtered reproductive long non-coding RNAs by genome-wide analyses of goat ovary at different estrus periods. BMC Genom. 2018, 19, 866. [Google Scholar] [CrossRef]
- Liu, Y.; Zhou, Z.; He, X.; Tao, L.; Jiang, Y.; Lan, R.; Hong, Q.; Chu, M. Integrated analyses of miRNA-mRNA expression profiles of ovaries reveal the crucial interaction networks that regulate the prolificacy of goats in the follicular phase. BMC Genom. 2021, 22, 812. [Google Scholar] [CrossRef] [PubMed]
- Thomson, D.W.; Dinger, M.E. Endogenous microRNA sponges: Evidence and controversy. Nat. Rev. Genet. 2016, 17, 272–283. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Kanehisa, M. Toward understanding the origin and evolution of cellular organisms. Protein Sci. 2019, 28, 1947–1951. [Google Scholar] [CrossRef]
- Kanehisa, M.; Furumichi, M.; Sato, Y.; Ishiguro-Watanabe, M.; Tanabe, M. KEGG: Integrating viruses and cellular organisms. Nucleic Acids Res. 2021, 49, D545–D551. [Google Scholar] [CrossRef] [PubMed]
- Berri, C.; Le Bihan-Duval, E.; Debut, M.; Sante-Lhoutellier, V.; Baeza, E.; Gigaud, V.; Jego, Y.; Duclos, M.J. Consequence of muscle hypertrophy on characteristics of Pectoralis major muscle and breast meat quality of broiler chickens. J. Anim. Sci. 2007, 85, 2005–2011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tripathi, A.K.; Patel, A.K.; Shah, R.K.; Patel, A.B.; Shah, T.M.; Bhatt, V.D.; Joshi, C.G. Transcriptomic dissection of myogenic differentiation signature in caprine by RNA-Seq. Mech. Dev. 2014, 132, 79–92. [Google Scholar] [CrossRef]
- Braun, T.; Gautel, M. Transcriptional mechanisms regulating skeletal muscle differentiation, growth and homeostasis. Nat. Rev. Mol. Cell Biol. 2011, 12, 349–361. [Google Scholar] [CrossRef]
- Zhou, J.; Xiong, Q.; Chen, H.; Yang, C.; Fan, Y. Identification of the Spinal Expression Profile of Non-coding RNAs Involved in Neuropathic Pain Following Spared Nerve Injury by Sequence Analysis. Front Mol. Neurosci. 2017, 10, 91. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Zhao, Y.; Bao, X.; Zhu, X.; Kwok, Y.K.; Sun, K.; Chen, X.; Huang, Y.; Jauch, R.; Esteban, M.A.; et al. LncRNA Dum interacts with Dnmts to regulate Dppa2 expression during myogenic differentiation and muscle regeneration. Cell Res. 2015, 25, 335–350. [Google Scholar] [CrossRef] [Green Version]
- Cabili, M.N.; Trapnell, C.; Goff, L.; Koziol, M.; Tazon-Vega, B.; Regev, A.; Rinn, J.L. Integrative annotation of human large intergenic noncoding RNAs reveals global properties and specific subclasses. Genes Dev. 2011, 25, 1915–1927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Derrien, T.; Johnson, R.; Bussotti, G.; Tanzer, A.; Djebali, S.; Tilgner, H.; Guernec, G.; Martin, D.; Merkel, A.; Knowles, D.G.; et al. The GENCODE v7 catalog of human long noncoding RNAs: Analysis of their gene structure, evolution, and expression. Genome Res. 2012, 22, 1775–1789. [Google Scholar] [CrossRef] [PubMed]
- Yue, B.; Li, H.; Liu, M.; Wu, J.; Li, M.; Lei, C.; Huang, B.; Chen, H. Characterization of lncRNA-miRNA-mRNA Network to Reveal Potential Functional ceRNAs in Bovine Skeletal Muscle. Front. Genet. 2019, 10, 91. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Cai, B.; Abdalla, B.A.; Zhu, X.; Zheng, M.; Han, P.; Nie, Q.; Zhang, X. LncIRS1 controls muscle atrophy via sponging miR-15 family to activate IGF1-PI3K/AKT pathway. J. Cachexia Sarcopenia Muscle 2019, 10, 391–410. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Zhang, Y.; Li, T.; Ma, Z.; Jia, H.; Chen, Q.; Zhao, Y.; Zhai, L.; Zhong, R.; Li, C.; et al. Long non-coding RNA Linc-RAM enhances myogenic differentiation by interacting with MyoD. Nat. Commun. 2017, 8, 14016. [Google Scholar] [CrossRef] [Green Version]
- Cesana, M.; Cacchiarelli, D.; Legnini, I.; Santini, T.; Sthandier, O.; Chinappi, M.; Tramontano, A.; Bozzoni, I. A long noncoding RNA controls muscle differentiation by functioning as a competing endogenous RNA. Cell 2011, 147, 358–369. [Google Scholar] [CrossRef] [Green Version]
- Knoll, M.; Lodish, H.F.; Sun, L. Long non-coding RNAs as regulators of the endocrine system. Nat. Rev. Endocrinol. 2015, 11, 151–160. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Qin, C.; Cao, G.; Xin, W.; Feng, C.; Zhang, W. Systematic Analysis of Long Noncoding RNAs in the Senescence-accelerated Mouse Prone 8 Brain Using RNA Sequencing. Mol. Ther. Nucleic Acids 2016, 5, e343. [Google Scholar] [CrossRef] [Green Version]
- Parisi, A.; Lacour, F.; Giordani, L.; Colnot, S.; Maire, P.; Le Grand, F. APC is required for muscle stem cell proliferation and skeletal muscle tissue repair. J. Cell Biol. 2015, 210, 717–726. [Google Scholar] [CrossRef] [Green Version]
- Micheli, L.; Leonardi, L.; Conti, F.; Maresca, G.; Colazingari, S.; Mattei, E.; Lira, S.A.; Farioli-Vecchioli, S.; Caruso, M.; Tirone, F. PC4/Tis7/IFRD1 stimulates skeletal muscle regeneration and is involved in myoblast differentiation as a regulator of MyoD and NF-kappaB. J. Biol. Chem. 2011, 286, 5691–5707. [Google Scholar] [CrossRef] [Green Version]
- Nie, M.; Deng, Z.L.; Liu, J.; Wang, D.Z. Noncoding RNAs, Emerging Regulators of Skeletal Muscle Development and Diseases. BioMed Res. Int. 2015, 2015, 676575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mueller, A.C.; Cichewicz, M.A.; Dey, B.K.; Layer, R.; Reon, B.J.; Gagan, J.R.; Dutta, A. MUNC, a long noncoding RNA that facilitates the function of MyoD in skeletal myogenesis. Mol Cell Biol. 2015, 35, 498–513. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhang, T.; Zhang, S.; Huang, J.; Zhang, G.; Xie, K.; Wang, J.; Wu, H.; Dai, G. Identification of Long Non-Coding RNA-Associated Competing Endogenous RNA Network in the Differentiation of Chicken Preadipocytes. Genes 2019, 10, 795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuang, L.; Lei, M.; Li, C.; Zhang, X.; Ren, Y.; Zheng, J.; Guo, Z.; Zhang, C.; Yang, C.; Mei, X.; et al. Identification of Long Non-Coding RNAs Related to Skeletal Muscle Development in Two Rabbit Breeds with Different Growth Rate. Int. J. Mol. Sci. 2018, 19, 2046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Zhong, J.; Zhang, C.; Chai, Z.; Cao, H.; Wang, J.; Zhu, J.; Wang, J.; Ji, Q. The whole-transcriptome landscape of muscle and adipose tissues reveals the ceRNA regulation network related to intramuscular fat deposition in yak. BMC Genom. 2020, 21, 347. [Google Scholar] [CrossRef]
- Zhao, R.; Li, F.Q.; Tian, L.L.; Shang, D.S.; Guo, Y.; Zhang, J.R.; Liu, M. Comprehensive analysis of the whole coding and non-coding RNA transcriptome expression profiles and construction of the circRNA-lncRNA co-regulated ceRNA network in laryngeal squamous cell carcinoma. Funct. Integr. Genom. 2019, 19, 109–121. [Google Scholar] [CrossRef]
- D’Cruz, R.; Plant, P.J.; Pablo, L.A.; Lin, S.; Chackowicz, J.; Correa, J.; Bain, J.; Batt, J. PDLIM7 is a novel target of the ubiquitin ligase Nedd4-1 in skeletal muscle. Biochem. J. 2016, 473, 267–276. [Google Scholar] [CrossRef]
- Camarata, T.; Krcmery, J.; Snyder, D.; Park, S.; Topczewski, J.; Simon, H.G. Pdlim7 (LMP4) regulation of Tbx5 specifies zebrafish heart atrio-ventricular boundary and valve formation. Dev. Biol. 2010, 337, 233–245. [Google Scholar] [CrossRef] [Green Version]
- Ge, X.; Vajjala, A.; McFarlane, C.; Wahli, W.; Sharma, M.; Kambadur, R. Lack of Smad3 signaling leads to impaired skeletal muscle regeneration. Am. J. Physiol. Endocrinol. Metab. 2012, 303, E90–E102. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Oligo Name | Primer Sequence (5′→3′) | Product Length/bp | Annealing Temperature/°C |
|---|---|---|---|
| TCONS_00036756 | F: AGGCTGCAATCCACGCTAA R: TCCAACTCTGTGTGACCCCAT | 170 | 60 |
| TCONS_00066056 | F: AGAAAATGAATCCCTGGAGTGTG R: AACGCTGACCACCATGATGAC | 114 | 60 |
| TCONS_00062751 | F: CTGGGAGAATACAAAGGGGG R: GGATCTACGGGCCTTTTGTCT | 247 | 60 |
| XR_001296113.2 | F: GCCGCCGTGAAGACTATTG R: CCATGAAGCCAGGGTACAAAC | 180 | 60 |
| TCONS_00150002 | F: CCCCGAATGTAAGCAATGAG R: AGGAGACCTACCGCTACCTGAG | 145 | 60 |
| TCONS_00134767 | F: CCCAACAAAGTGCCCAGAC R: GGAGAAGACGGCGTTATGC | 142 | 60 |
| TCONS_00126170 | F: GCTAGTCCCAGACAGCATTCAT R: GGTGTTGTTCTCGCCTGGAA | 252 | 60 |
| TCONS_00124841 | F: CCCTTACCACAGGCACCACT R: CAGGTGAGAAGGTGTGTTCTGG | 104 | 60 |
| TCONS_00121766 | F: TGTCCCCAACCTCGGTATCT R: GGTCAAACCTCTGAGCCTCG | 211 | 60 |
| TCONS_00026838 | F: CTTCTCCTTGCTTGGCACCT R: CAGGTGCCAAGCAAGGAGA | 121 | 60 |
| GAPDH | F: CACGGCACAGTCAAGGCAG R: AGATGATGACCCTCTTGGCG | 196 | 60 |
| Sample Name | Raw Reads | Clean Reads | Raw Bases (G) | Clean Bases (G) | Error Rate (%) | Q20 (%) | Q30 (%) | GC Content (%) |
|---|---|---|---|---|---|---|---|---|
| mon1_1 | 98721676 | 97036650 | 14.81 | 14.56 | 0.03 | 97.47 | 93 | 52.99 |
| mon1_2 | 104194360 | 101986738 | 15.63 | 15.3 | 0.03 | 97.29 | 92.64 | 55.08 |
| mon1_3 | 99110136 | 97525086 | 14.87 | 14.63 | 0.03 | 96.95 | 91.87 | 53.06 |
| mon9_1 | 98408274 | 96645866 | 14.76 | 14.5 | 0.03 | 97.4 | 92.79 | 51.64 |
| mon9_2 | 90534786 | 88937344 | 13.58 | 13.34 | 0.03 | 97.48 | 92.97 | 53.35 |
| mon9_3 | 106156574 | 101082860 | 15.92 | 15.16 | 0.03 | 96.92 | 91.93 | 54.31 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, H.; Wang, X.; Li, W.; Liu, J.; Fan, Y.; Zhang, H.; Yang, J.; Gao, Y.; Liu, Y. Identification and Characterization of lncRNAs Expression Profile Related to Goat Skeletal Muscle at Different Development Stages. Animals 2022, 12, 2683. https://doi.org/10.3390/ani12192683
Han H, Wang X, Li W, Liu J, Fan Y, Zhang H, Yang J, Gao Y, Liu Y. Identification and Characterization of lncRNAs Expression Profile Related to Goat Skeletal Muscle at Different Development Stages. Animals. 2022; 12(19):2683. https://doi.org/10.3390/ani12192683
Chicago/Turabian StyleHan, Haiyin, Xianwei Wang, Wentao Li, Jiannan Liu, Yekai Fan, Hui Zhang, Junqi Yang, Yahui Gao, and Yufang Liu. 2022. "Identification and Characterization of lncRNAs Expression Profile Related to Goat Skeletal Muscle at Different Development Stages" Animals 12, no. 19: 2683. https://doi.org/10.3390/ani12192683