
Global Warming and Dairy Cattle: How to Control and Reduce Methane Emission



Abstract
:Simple Summary
Abstract
1. Introduction
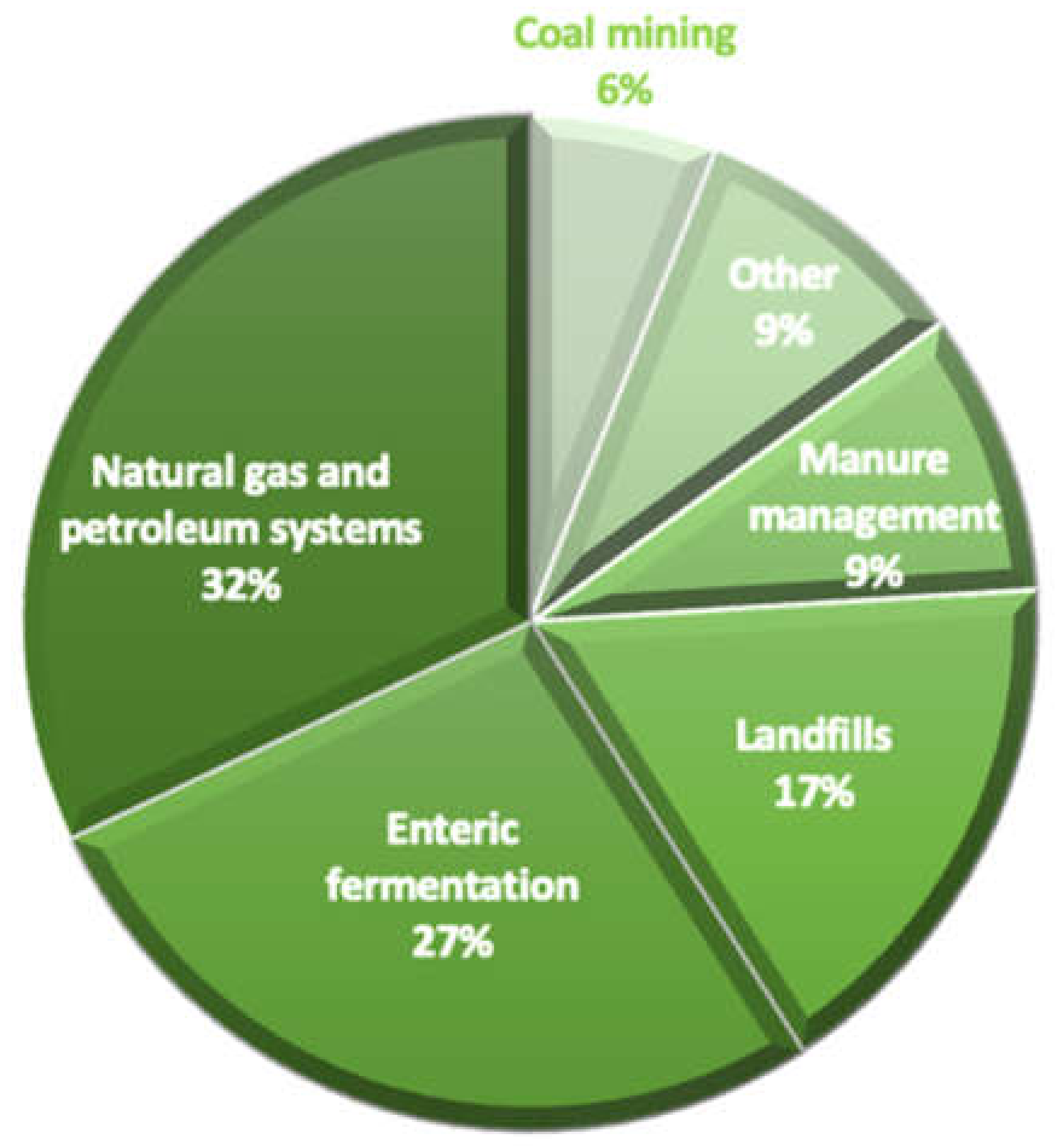
2. Role of Dairy Cattle in Global Warming
3. Measurement and Estimation of CH4
3.1. Methane Prediction Models

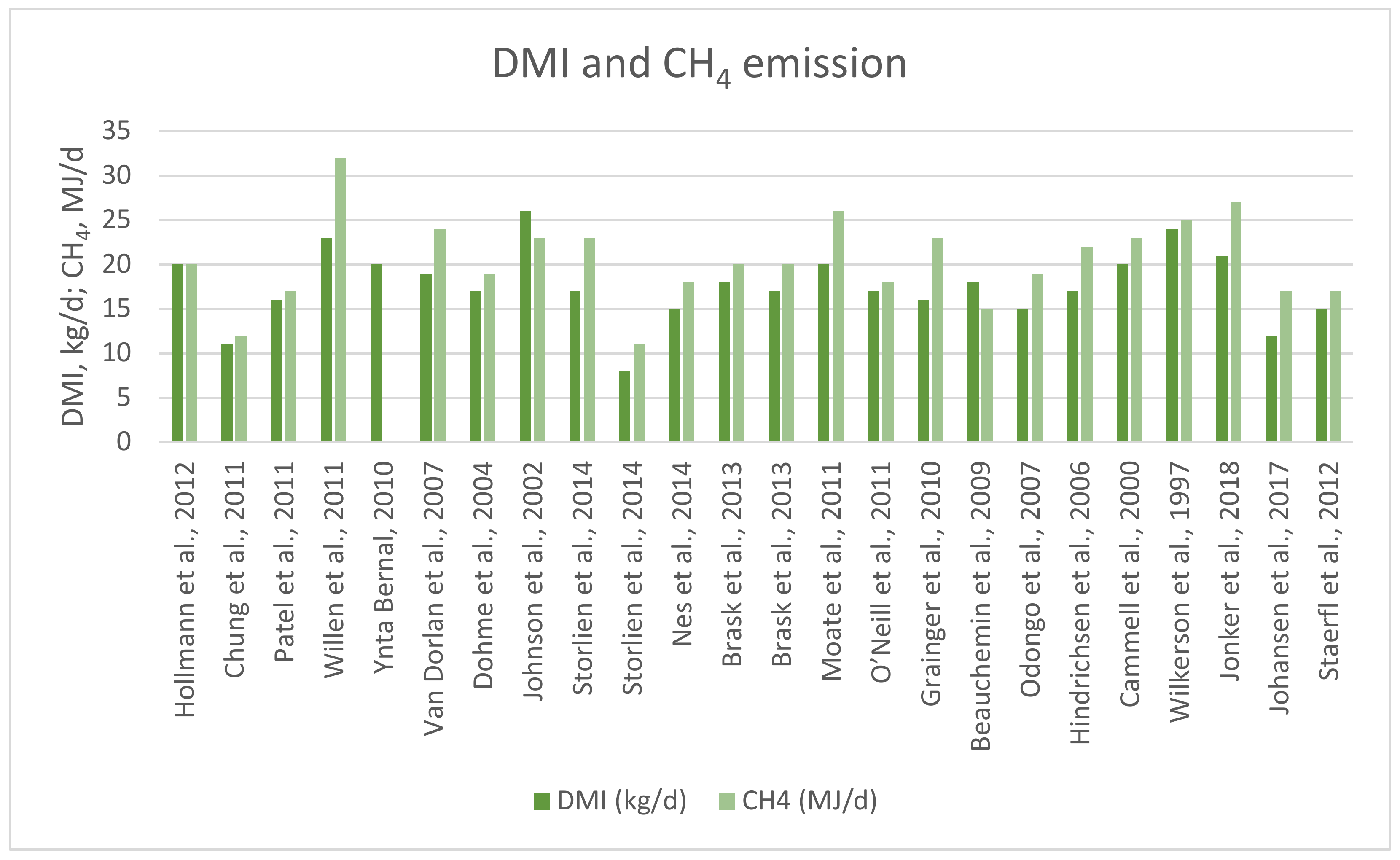
| Lactation Stage | Roughage | Concentrate | DMI (kg/d) | CH4 Collection Technique | CH4 (MJ/d) | References |
|---|---|---|---|---|---|---|
| L | Corn silage | Ground corn | 20 | Room tracer approach | 20 (14–26) | [66] |
| NL | Grass hay or barley silage | Barley grain | 11 | Sulfur hexafluoride tracer gas technique | 12 (11–17) | [67] |
| L | Grass silage | Oats, barley, peas and rapeseed cake | 16 | Sulfur hexafluoride tracer gas technique | 17 (16–18) | [68] |
| L | Grass silage | Barley, wheat and maize | 23 | Sulfur hexafluoride tracer gas technique | 32 (28–36) | [69] |
| L | Grass silage | Barley, wheat and oats | 20 | Sulfur hexafluoride tracer gas technique | 26 (24–28) | [70] |
| L | Ryegrass, white and red clover | Pelleted barley | 19 | Chamber | 24 (23–26) | [71] |
| L | Grass and maize silage | Barley | 17 | Chamber | 19 (17–21) | [72] |
| L | Alfalfa hay and alfalfa silage | Barley, corn and peas | 26 | Room tracer approach | 23 (22–25) | [73] |
| L | Grass silage | Barley | 17 | Sulfur hexafluoride tracer gas technique | 23 (20–29) | [62] |
| NL | Grass silage | Wheat starch (non-NDF concentrate) | 8 | Sulfur hexafluoride tracer gas technique | 11 (10–12) | [62] |
| L | Grass silage | Wheat starch (non-NDF concentrate) | 15 | Sulfur hexafluoride tracer gas technique | 18 (17–19) | [74] |
| L | Grass and corn silage | Rapeseed meal, rapeseed cake, cracked rapeseed and rapeseed oil | 18 | Sulfur hexafluoride tracer gas technique | 20 (17–23) | [75] |
| L | Grass silage and maize silage | Rapeseed meal, whole crushed rapeseed | 17 | Sulfur hexafluoride tracer gas technique | 20 (18–22) | [76] |
| L | Alfalfa hay and ryegrass silage | Cracked wheat grain | 20 | Chamber | 26 (25–28) | [77] |
| L | Corn and grass silage | Soybean meal and rolled barley | 17 | Sulfur hexafluoride tracer gas technique | 18 (14–22) | [78] |
| L | Corn silage and alfalfa haylage | Cracked wheat grain | 16 | Sulfur hexafluoride tracer gas technique | 23 (21–25) | [79] |
| L | Barley silage | Steam rolled barley and pelleted supplement | 18 | Chamber | 15 (13–16) | [30] |
| L | Haylage, corn silage and high | Corn gluten and soybean meal | 15 | Head hood | 19 (15–23) | [80] |
| L | Hay, grass and corn silage | Barley and wheat bran | 17 | Chamber | 22 (18–24) | [81] |
| L | Corn and grass silage | Rapeseed meal, sunflower meal, ground wheat and maize gluten feed | 20 | Chamber | 23 (22–23) | [82] |
| L | Alfalfa silage High moisture corn and | High moisture corn and dry corn | 24 | Chamber | 25 (24–26) | [83] |
| L | Ryegrass, white clover, or mature, diverse pasture | 0 | 21 | Greenfeed system | 27 (26–28) | [84] |
| L | Grass clover silage | 0 | 12 | Chamber | 17 | [85] |
| L | Ryegrass | 0 | 15 | Chamber | 17 (16-19) | [86] |
3.2. Biomarkers for Controlling of GHG
3.2.1. Rumination Time
3.2.2. Rumen Microbiome
3.3. Methane Emissions and Animal Performance in Dairy
4. Methods to Reduce GHG
4.1. Feed Supplements
4.1.1. Algae, Bromoform
Algae Cultivating and Sustainability
4.1.2. Phytocompounds: Polyphenolic Substances (Tannins, Saponins), Essential Oils, Flavonoids
Phytocompounds. Sustainability
4.1.3. Oils: Rapeseed Oil
4.2. Reducing Greenhouse Gas Emissions through Genetic Selection
5. Conclusions
6. Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Black, J.L.; Davison, T.M.; Box, I. Methane Emissions from Ruminants in Australia: Mitigation Potential and Applicability of Mitigation Strategies. Animals 2021, 11, 951. [Google Scholar] [CrossRef] [PubMed]
- De Gouw, J.A.; Veefkind, J.P.; Roosenbrand, E.; Dix, B.; Lin, J.C.; Landgraf, J.; Levelt, P.F. Daily Satellite Observations of Methane from Oil and Gas Production Regions in the United States. Sci. Rep. 2020, 10, 1379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- La, H.; Hettiaratchi, J.P.A.; Achari, G.; Hettiaratchi, J.P.; Achari, G.; Dunfield, P.F. Biofiltration of methane. Bioresour. Technol. 2018, 268, 759–772. [Google Scholar] [CrossRef] [PubMed]
- Your Bowl of Rice Is Hurting the Climate Too. Bloomberg.com, 3 June 2019. Available online: https://www.bloomberg.com/news/articles/2019-06-03/your-bowl-of-rice-is-hurting-the-climate-too (accessed on 15 September 2022).
- Wang, Z.-H.; Wang, L.-H.; Liang, H.; Peng, T.; Xia, G.-P.; Zhang, J.; Zhao, Q.-Z. Methane and nitrous oxide emission characteristics of high-yielding rice field. Environ. Sci. Pollut. Res. Int. 2021, 28, 15021–15031. [Google Scholar] [CrossRef]
- Sun, H.; Zhou, S.; Zhang, J.; Wang, C. Year-to-year climate variability affects methane emission from paddy fields under irrigated conditions. Environ. Sci. Pollut. Res. Int. 2020, 27, 14780–14789. [Google Scholar] [CrossRef]
- Linquist, B.; van Groenigen, K.J.; Adviento-Borbe, M.A.; Pittelkow, C.; van Kessel, C. An agronomic assessment of greenhouse gas emissions from major cereal crops. Glob. Chang. Biol. 2012, 18, 194–209. [Google Scholar] [CrossRef]
- Vázquez-Carrillo, M.F.; Montelongo-Pérez, H.D.; González-Ronquillo, M.; Castillo-Gallegos, E.; Castelán-Ortega, O. Effects of Three Herbs on Methane Emissions from Beef Cattle. Animals 2020, 10, 1671. [Google Scholar] [CrossRef]
- Hardan, A.; Garnsworthy, P.C.; Bell, M.J. Detection of Methane Eructation Peaks in Dairy Cows at a Robotic Milking Station Using Signal Processing. Animals 2022, 12, 26. [Google Scholar] [CrossRef]
- Garnsworthy, P.C.; Difford, G.F.; Bell, M.J.; Bayat, A.; Huhtanen, P. Comparison of Methods to Measure Methane for Use in Genetic Evaluation of Dairy Cattle. Animals 2019, 9, 837. [Google Scholar] [CrossRef] [Green Version]
- Dillon, J.A.; Stackhouse-Lawson, K.R.; Thoma, G.J.; Gunter, S.A.; Rotz, C.A. Current state of enteric methane and the carbon footprint of beef and dairy cattle in the United States. Anim. Front. 2021, 11, 57–68. [Google Scholar] [CrossRef]
- Holtshausen, L.; Benchaar, C.; Kröbel, R.; Beauchemin, K.A. Canola Meal versus Soybean Meal as Protein Supplements in the Diets of Lactating Dairy Cows Affects the Greenhouse Gas Intensity of Milk. Animals 2021, 11, 1636. [Google Scholar] [CrossRef]
- U.S. Environmental Protection Agency. Overview of Greenhouse Gases. 2015. Available online: https://www.epa.gov/ghgemissions/overview-greenhouse-gases (accessed on 30 June 2022).
- Grešáková, Ľ.; Holodová, M.; Szumacher-Strabel, M.; Huang, H.; Ślósarz, P. Mineral status and enteric methane production in dairy cows during different stages of lactation. BMC Vet. Res. 2021, 17, 287. [Google Scholar] [CrossRef]
- Mikuła, R.; Pszczola, M.; Rzewuska, K.; Mucha, S.; Nowak, W.; Strabel, T. The Effect of Rumination Time on Milk Performance and Methane Emission of Dairy Cows Fed Partial Mixed Ration Based on Maize Silage. Animals 2022, 12, 50. [Google Scholar] [CrossRef]
- Kozłowska, M.; Cieślak, A.; Jóźwik, A.; El-Sherbiny, M.; Gogulski, M. Effects of Partially Replacing Grass Silage by Lucerne Silage Cultivars in a High-Forage Diet on Ruminal Fermentation, Methane Production, and Fatty Acid Composition in the Rumen and Milk of Dairy Cows. 2021. Available online: https://pubag.nal.usda.gov/catalog/7367430 (accessed on 30 June 2022).
- Gerber, P.J.; Mottet, A.; Opio, C.I.; Falcucci, A.; Teillard, F. Environmental impacts of beef production: Review of challenges and perspectives for durability. Meat Sci. 2015, 109, 2–12. [Google Scholar] [CrossRef]
- Lassen, J.; Løvendahl, P.; Madsen, J. Accuracy of noninvasive breath methane measurements using Fourier transform infrared methods on individual cows. J. Dairy Sci. 2012, 95, 890–898. [Google Scholar] [CrossRef] [Green Version]
- Knapp, J.R.; Laur, G.L.; Vadas, P.A.; Weiss, W.P.; Tricarico, J.M. Invited review: Enteric methane in dairy cattle production: Quantifying the opportunities and impact of reducing emissions. J. Dairy Sci. 2014, 97, 3231–3261. [Google Scholar] [CrossRef] [Green Version]
- Cottle, D.J.; Velazco, J.; Hegarty, R.S.; Mayer, D.G. Estimating daily methane production in individual cattle with irregular feed intake patterns from short-term methane emission measurements. Animal 2015, 9, 1949–1957. [Google Scholar] [CrossRef] [Green Version]
- Janssen, P.H. Influence of hydrogen on rumen methane formation and fermentation balances through microbial growth kinetics and fermentation thermodynamics. Anim. Feed Sci. Technol. 2010, 160, 1–22. [Google Scholar] [CrossRef]
- Jiménez-Ocampo, R.; Montoya-Flores, M.D.; Herrera-Torres, E.; Pámanes-Carrasco, G.; Arceo-Castillo, J.I. Effect of Chitosan and Naringin on Enteric Methane Emissions in Crossbred Heifers Fed Tropical Grass. Animals 2021, 11, 1599. [Google Scholar] [CrossRef]
- Kinley, R.D.; de Nys, R.; Vucko, M.J.; Machado, L.; Tomkins, N.W. The red macroalgae Asparagopsis taxiformis is a potent natural antimethanogenic that reduces methane production during in vitro fermentation with rumen fluid. Anim. Prod. Sci. 2016, 56, 282–289. [Google Scholar] [CrossRef]
- Patra, A.K. The effect of dietary fats on methane emissions, and its other effects on digestibility, rumen fermentation and lactation performance in cattle: A meta-analysis. Livest. Sci. 2013, 155, 244–254. [Google Scholar] [CrossRef]
- Bekele, W.; Guinguina, A.; Zegeye, A.; Simachew, A.; Ramin, M. Contemporary Methods of Measuring and Estimating Methane Emission from Ruminants. Methane 2022, 1, 82–95. [Google Scholar] [CrossRef]
- Negussie, E.; Lehtinen, J.; Mäntysaari, P.; Lidauer, M.H. Non-invasive individual methane measurement in dairy cows. Animal 2017, 11, 890–899. [Google Scholar] [CrossRef]
- Scaling Point and Plot Measurements of Greenhouse Gas Fluxes, Balances, and Intensities to Whole Farms and Landscapes—CIFOR Knowledge. Available online: https://www.cifor.org/knowledge/publication/5803/ (accessed on 30 June 2022).
- Kristiansen, S.; Painter, J.; Shea, M. Animal Agriculture and Climate Change in the US and UK Elite Media: Volume, Responsibilities, Causes and Solutions. Environ. Commun. 2021, 15, 153–172. [Google Scholar] [CrossRef]
- Beauchemin, K.A.; Kreuzer, M.; O’Mara, F.; McAllister, T.A. Nutritional management for enteric methane abatement: A review. Aust. J. Exp. Agric. 2008, 48, 21–27. [Google Scholar] [CrossRef]
- Beauchemin, K.A.; McGinn, S.M.; Benchaar, C.; Holtshausen, L. Crushed sunflower, flax, or canola seeds in lactating dairy cow diets: Effects on methane production, rumen fermentation, and milk production. J. Dairy Sci. 2009, 92, 2118–2127. [Google Scholar] [CrossRef]
- Patra, A.K. Trends and Projected Estimates of GHG Emissions from Indian Livestock in Comparisons with GHG Emissions from World and Developing Countries. Asian Australas. J. Anim. Sci. 2014, 27, 592–599. [Google Scholar] [CrossRef] [Green Version]
- Herrero, M.; Thornton, P.K. Livestock and global change: Emerging issues for sustainable food systems. Proc. Natl. Acad. Sci. USA 2013, 110, 20878–20881. [Google Scholar] [CrossRef] [Green Version]
- Gerber, P.J.; Food and Agriculture Organization of the United Nations. Tackling Climate Change Through Livestock: A Global Assessment of Emissions and Mitigation Opportunities; Food and Agriculture Organization of the United Nations: Rome, Italy, 2013. [Google Scholar]
- Aan den Toorn, S.I.; Worrell, E.; Broek, M. Meat, dairy, and more: Analysis of material, energy, and greenhouse gas flows of the meat and dairy supply chains in the EU28 for 2016. J. Ind. Ecol. 2020, 24, 601–614. [Google Scholar] [CrossRef] [Green Version]
- Tubiello, F.N.; Salvatore, M.; Rossi, S.; Ferrara, A.; Fitton, N.; Smith, P. The FAOSTAT database of greenhouse gas emissions from agriculture. Environ. Res. Lett. 2013, 8, 015009. [Google Scholar] [CrossRef]
- Can We Make Cow Burps Climate-Friendly? | Research and Innovation. Available online: https://ec.europa.eu/research-and-innovation/en/horizon-magazine/can-we-make-cow-burps-climate-friendly (accessed on 23 July 2022).
- Abbott, D.W.; Aasen, I.M.; Beauchemin, K.A.; Grondahl, A.; Gruninger, R. Seaweed and Seaweed Bioactives for Mitigation of Enteric Methane: Challenges and Opportunities. Animals 2020, 10, 2432. [Google Scholar] [CrossRef] [PubMed]
- Huws, S.A.; Creevey, C.J.; Oyama, L.B.; Mizrahi, I.; Denman, S.E. Addressing Global Ruminant Agricultural Challenges Through Understanding the Rumen Microbiome: Past, Present, and Future. Front. Microbiol. 2018, 9, 2161. [Google Scholar] [CrossRef] [PubMed]
- Henderson, G.; Cox, F.; Ganesh, S.; Jonker, A.; Young, W.; Janssen, P.H. Rumen microbial community composition varies with diet and host, but a core microbiome is found across a wide geographical range. Sci. Rep. 2015, 5, 14567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasson, G.; Kruger Ben-Shabat, S.; Seroussi, E.; Doron-Faigenboim, A.; Shterzer, N. Heritable Bovine Rumen Bacteria Are Phylogenetically Related and Correlated with the Cow’s Capacity To Harvest Energy from Its Feed. mBio 2017, 8, e00703-17. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Szumacher-Strabel, M.; Patra, A.K.; Ślusarczyk, S.; Lechniak, D. Chemical and phytochemical composition, in vitro ruminal fermentation, methane production, and nutrient degradability of fresh and ensiled Paulownia hybrid leaves. Anim. Feed Sci. Technol. 2021, 279, 115038. [Google Scholar] [CrossRef]
- Hammond, K.J.; Crompton, L.A.; Bannink, A.; Dijkstra, J.; Yáñez-Ruiz, D.R. Review of current in vivo measurement techniques for quantifying enteric methane emission from ruminants. Anim. Feed Sci. Technol. 2016, 219, 13–30. [Google Scholar] [CrossRef] [Green Version]
- Lassen, J.; Løvendahl, P. Heritability estimates for enteric methane emissions from Holstein cattle measured using noninvasive methods. J. Dairy Sci. 2016, 99, 1959–1967. [Google Scholar] [CrossRef] [Green Version]
- Garnsworthy, P.C.; Craigon, J.; Hernandez-Medrano, J.H.; Saunders, N. On-farm methane measurements during milking correlate with total methane production by individual dairy cows. J. Dairy Sci. 2012, 95, 3166–3180. [Google Scholar] [CrossRef] [Green Version]
- Huhtanen, P.; Cabezas-Garcia, E.H.; Utsumi, S.; Zimmerman, S. Comparison of methods to determine methane emissions from dairy cows in farm conditions. J. Dairy Sci. 2015, 98, 3394–3409. [Google Scholar] [CrossRef] [Green Version]
- Murray, R.M.; Bryant, A.M.; Leng, R.A. Rates of production of methane in the rumen and large intestine of sheep. Br. J. Nutr. 1976, 36, 1–14. [Google Scholar] [CrossRef]
- Brouček, J. Methods of Methane Measurement in Ruminants. Slovak J. Anim. Sci. 2014, 47, 51–60. [Google Scholar]
- Hammond, K.J.; Waghorn, G.C.; Hegarty, R.S. The GreenFeed system for measurement of enteric methane emission from cattle. Anim. Prod. Sci. 2016, 56, 181–189. [Google Scholar] [CrossRef]
- Hellwing, A.L.F.; Lund, P.; Weisbjerg, M.R.; Brask, M.; Hvelplund, T. Technical note: Test of a low-cost and animal-friendly system for measuring methane emissions from dairy cows. J. Dairy Sci. 2012, 95, 6077–6085. [Google Scholar] [CrossRef]
- Yan, T.; Mayne, C.S.; Gordon, F.G.; Porter, M.G.; Agnew, R.E. Mitigation of enteric methane emissions through improving efficiency of energy utilization and productivity in lactating dairy cows. J. Dairy Sci. 2010, 93, 2630–2638. [Google Scholar] [CrossRef] [Green Version]
- Kristen, J.; Mark, H.; Hal, W.; Lamb, B.; Zimmerman, P. Measurement of methane emissions from ruminant livestock using a sulfur hexafluoride tracer technique. Environ. Sci. Technol. 1994, 28, 359–362. [Google Scholar]
- Berndt, A.; Boland, T.; Deighton, M.; Gere, J.; Grainger, C. Guidelines for Use of the Sulphur Hexafluoride (SF6) Tracer Technique to Measure Enteric Methane Emissions from Ruminants; Ministry for Primary Industries: Wellington, New Zealand, 2014. [CrossRef]
- Madsen, J.; Bjerg, B.S.; Hvelplund, T.; Weisbjerg, M.R.; Lund, P. Methane and carbon dioxide ratio in excreted air for quantification of the methane production from ruminants. Livest. Sci. 2010, 129, 223–227. [Google Scholar] [CrossRef]
- Goopy, J.P.; Chang, C.; Tomkins, N. A Comparison of Methodologies for Measuring Methane Emissions from Ruminants. In Methods for Measuring Greenhouse Gas Balances and Evaluating Mitigation Options in Smallholder Agriculture; Rosenstock, T.S., Rufino, M.C., Butterbach-Bahl, K., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 97–117. [Google Scholar]
- Huhtanen, P.; Bayat, A.R.; Lund, P.; Hellwing, A.L.F.; Weisbjerg, M.R. Short communication: Variation in feed efficiency hampers use of carbon dioxide as a tracer gas in measuring methane emissions in on-farm conditions. J. Dairy Sci. 2020, 103, 9090–9095. [Google Scholar] [CrossRef]
- Chagunda, M.G.G.; Yan, T. Do methane measurements from a laser detector and an indirect open-circuit respiration calorimetric chamber agree sufficiently closely? Anim. Feed Sci. Technol. 2011, 165, 8–14. [Google Scholar] [CrossRef]
- Chagunda, M.G.G. Opportunities and challenges in the use of the Laser Methane Detector to monitor enteric methane emissions from ruminants. Anim. Int. J. Anim. Biosci. 2013, 7 (Suppl. 2), 394–400. [Google Scholar] [CrossRef] [Green Version]
- Sorg, D.; Mühlbach, S.; Rosner, F.; Kuhla, B.; Derno, M. Agreement of Two Laser Methane Detectors with Respiration Chambers and in a Dairy Barn; Wageningen Academic Publishers: Wageningen, The Netherlands, 2016; Volume 22, p. 267. [Google Scholar]
- Oss, D.; Marcondes, M.; Machado, F.; Pereira, L.; Tomich, T.; Ribeiro, G.; Chizzotti, M.; Ferreira, A.; Campos, M.; Mauricio, R.; et al. An evaluation of the face mask system based on short-term measurements compared with the sulfur hexafluoride (SF6) tracer, and respiration chamber techniques for measuring CH4 emissions. Anim. Feed Sci. Technol. 2016, 216, 49–57. [Google Scholar] [CrossRef]
- Brosh, A. Heart rate measurements as an index of energy expenditure and energy balance in ruminants: A review. J. Anim. Sci. 2007, 85, 1213–1227. [Google Scholar] [CrossRef]
- Zhang, B. Development of Prediction Models of Methane Production by Sheep and Cows Using Rumen Microbiota Data. Master Thesis, Ohio State University, Columbus, OH, USA, 2018. [Google Scholar]
- Storlien, T.M.; Volden, H.; Almøy, T.; Beauchemin, K.A.; McAllister, T.A.; Harstad, O.M. Prediction of enteric methane production from dairy cows. Acta Agric. Scand. Sect. Anim. Sci. 2014, 64, 98–109. [Google Scholar] [CrossRef]
- Niu, P.; Schwarm, A.; Bonesmo, H.; Kidane, A.; Aspeholen, Å.B.; Storlien, T.M.; Kreuzer, M.; Alvarez, C.; Sommerseth, J.K.; Prestløkken, E. A Basic Model to Predict Enteric Methane Emission from Dairy Cows and Its Application to Update Operational Models for the National Inventory in Norway. Animals 2021, 11, 1891. [Google Scholar] [CrossRef]
- Niu, M.; Kebreab, E.; Hristov, A.N.; Oh, J.; Arndt, C.; Bannink, A.; Bayat, A.; Brito, A.F.; Boland, T.; Casper, D.; et al. Prediction of enteric methane production, yield, and intensity in dairy cattle using an intercontinental database. Glob. Chang. Biol. 2018, 24, 3368–3389. [Google Scholar] [CrossRef] [Green Version]
- Warner, D.; Bannink, A.; Hatew, B.; van Laar, H.; Dijkstra, J. Effects of grass silage quality and level of feed intake on enteric methane production in lactating dairy cows. J. Anim. Sci. 2017, 95, 3687–3700. [Google Scholar] [CrossRef]
- Hollmann, M.; Powers, W.J.; Fogiel, A.C.; Liesman, J.S.; Bello, N.M.; Beede, D.K. Enteric methane emissions and lactational performance of Holstein cows fed different concentrations of coconut oil. J. Dairy Sci. 2012, 95, 2602–2615. [Google Scholar] [CrossRef]
- Chung, Y.-H.; He, M.L.; McGinn, S.M.; McAllister, T.A.; Beauchemin, K.A. Linseed suppresses enteric methane emissions from cattle fed barley silage, but not from those fed grass hay. Anim. Feed Sci. Technol. 2011, 166–167, 321–329. [Google Scholar] [CrossRef]
- Patel, M.; Wredle, E.; Börjesson, G.; Danielsson, R.; Iwaasa, A.D.; Spörndly, E.; Bertilsson, J. Enteric methane emissions from dairy cows fed different proportions of highly digestible grass silage. Acta Agric. Scand. Sect. Anim. Sci. 2011, 61, 128–136. [Google Scholar] [CrossRef]
- Willén, A. Methane Production from Dairy Cows. 2011, Volume 335. Available online: https://stud.epsilon.slu.se/2328/ (accessed on 17 September 2022).
- Bernal, C.Y. Methane Production of Dairy Cows Fed Cereals with or Without Protein Supplement and High Quality Silage; Swedish University of Agricultural Science: Uppsala, Sweden, 2010; Volume 317, p. 46. [Google Scholar]
- Van Dorland, H.A.; Wettstein, H.-R.; Leuenberger, H.; Kreuzer, M. Effect of supplementation of fresh and ensiled clovers to ryegrass on nitrogen loss and methane emission of dairy cows. Livest. Sci. 2007, 111, 57–69. [Google Scholar] [CrossRef]
- Dohme, F.; Machmüller, A.; Sutter, F.; Kreuzer, M. Digestive and metabolic utilization of lauric, myristic and stearic acid in cows, and associated effects on milk fat quality. Arch. Anim. Nutr. 2004, 58, 99–116. [Google Scholar] [CrossRef]
- Johnson, K.A.; Kincaid, R.L.; Westberg, H.H.; Gaskins, C.T.; Lamb, B.K.; Cronrath, J.D. The Effect of Oilseeds in Diets of Lactating Cows on Milk Production and Methane Emissions. J. Dairy Sci. 2002, 85, 1509–1515. [Google Scholar] [CrossRef]
- Nes, S.K.; Garmo, T.; Chaves, A.V.; Prestløkken, E.; Volden, H.; Iwaasa, A.D.; Krizsan, S.J.; Beauchemin, K.A.; McAllister, T.A.; Norell, L.; et al. Effects of maturity of ensiled grass on enteric methane emissions from dairy cows offered with two levels of concentrate. 2014, unpublished manuscript. [Google Scholar]
- Brask, M.; Lund, P.; Hellwing, A.L.F.; Poulsen, M.; Weisbjerg, M.R. Enteric methane production, digestibility and rumen fermentation in dairy cows fed different forages with and without rapeseed fat supplementation. Anim. Feed Sci. Technol. 2013, 184, 67–79. [Google Scholar] [CrossRef]
- Brask, M.; Lund, P.; Weisbjerg, M.R.; Hellwing, A.L.F.; Poulsen, M.; Larsen, M.K.; Hvelplund, T. Methane production and digestion of different physical forms of rapeseed as fat supplements in dairy cows. J. Dairy Sci. 2013, 96, 2356–2365. [Google Scholar] [CrossRef] [Green Version]
- Moate, P.J.; Williams, S.R.O.; Grainger, C.; Hannah, M.C.; Ponnampalam, E.N.; Eckard, R.J. Influence of cold-pressed canola, brewers grains and hominy meal as dietary supplements suitable for reducing enteric methane emissions from lactating dairy cows. Anim. Feed Sci. Technol. 2011, 166–167, 254–264. [Google Scholar] [CrossRef]
- O’Neill, B.F.; Deighton, M.H.; O’Loughlin, B.M.; Mulligan, F.J.; Boland, T.M.; O’Donovan, M.; Lewis, E. Effects of a perennial ryegrass diet or total mixed ration diet offered to spring-calving Holstein-Friesian dairy cows on methane emissions, dry matter intake, and milk production. J. Dairy Sci. 2011, 94, 1941–1951. [Google Scholar] [CrossRef] [Green Version]
- Grainger, C.; Beauchemin, K.A. Can enteric methane emissions from ruminants be lowered without lowering their production? Anim. Feed Sci. Technol. 2011, 166–167, 308–320. [Google Scholar] [CrossRef]
- Odongo, N.E.; Or-Rashid, M.M.; Kebreab, E.; France, J.; McBride, B.W. Effect of Supplementing Myristic Acid in Dairy Cow Rations on Ruminal Methanogenesis and Fatty Acid Profile in Milk. J. Dairy Sci. 2007, 90, 1851–1858. [Google Scholar] [CrossRef] [Green Version]
- Hindrichsen, I.K.; Wettstein, H.-R.; Machmüller, A.; Kreuzer, M. Methane emission, nutrient degradation and nitrogen turnover in dairy cows and their slurry at different milk production scenarios with and without concentrate supplementation. Agric. Ecosyst. Environ. 2006, 113, 150–161. [Google Scholar] [CrossRef]
- Cammell, S.B.; Sutton, J.D.; Beever, D.E.; Humphries, D.J.; Phipps, R.H. The effect of crop maturity on the nutritional value of maize silage for lactating dairy cows. 1. Energy and nitrogen utilization. Anim. Sci. 2000, 71, 381–390. [Google Scholar] [CrossRef]
- Wilkerson, V.A.; Glenn, B.P.; McLeod, K.R. Energy and Nitrogen Balance in Lactating Cows Fed Diets Containing Dry or High Moisture Corn in Either Rolled or Ground Form. J. Dairy Sci. 1997, 80, 2487–2496. [Google Scholar] [CrossRef]
- Jonker, A.; Farrell, L.; Scobie, D.; Dynes, R.; Edwards, G.; Hague, H.; McAuliffe, R.; Taylor, A.; Knight, T.; Waghorn, G.; et al. Methane and carbon dioxide emissions from lactating dairy cows grazing mature ryegrass/white clover or a diverse pasture comprising ryegrass, legumes and herbs. Anim. Prod. Sci. 2018, 59, 1063–1069. [Google Scholar] [CrossRef]
- Johansen, M.; Hellwing, A.L.F.; Lund, P.; Weisbjerg, M.R. Metabolisable protein supply to lactating dairy cows increased with increasing dry matter concentration in grass-clover silage. Anim. Feed Sci. Technol. 2017, 227, 95–106. [Google Scholar] [CrossRef]
- Staerfl, S.M.; Amelchanka, S.L.; Kälber, T.; Soliva, C.R.; Kreuzer, M.; Zeitz, J.O. Effect of feeding dried high-sugar ryegrass (‘AberMagic’) on methane and urinary nitrogen emissions of primiparous cows. Livest. Sci. 2012, 150, 293–301. [Google Scholar] [CrossRef]
- Clément, P.; Guatteo, R.; Delaby, L.; Rouillé, B.; Chanvallon, A.; Philipot, J.M.; Bareille, N. Short communication: Added value of rumination time for the prediction of dry matter intake in lactating dairy cows. J. Dairy Sci. 2014, 97, 6531–6535. [Google Scholar] [CrossRef] [Green Version]
- Kaufman, E.I.; Asselstine, V.H.; LeBlanc, S.J.; Duffield, T.F.; DeVries, T.J. Association of rumination time and health status with milk yield and composition in early-lactation dairy cows. J. Dairy Sci. 2018, 101, 462–471. [Google Scholar] [CrossRef]
- Watt, L.J.; Clark, C.E.F.; Krebs, G.L.; Petzel, C.E.; Nielsen, S.; Utsumi, S.A. Differential rumination, intake, and enteric methane production of dairy cows in a pasture-based automatic milking system. J. Dairy Sci. 2015, 98, 7248–7263. [Google Scholar] [CrossRef] [Green Version]
- Byskov, M.V.; Nadeau, E.; Johansson, B.E.O.; Nørgaard, P. Variations in automatically recorded rumination time as explained by variations in intake of dietary fractions and milk production, and between-cow variation. J. Dairy Sci. 2015, 98, 3926–3937. [Google Scholar] [CrossRef] [Green Version]
- Zetouni, L.; Difford, G.F.; Lassen, J.; Byskov, M.V.; Norberg, E.; Løvendahl, P. Is rumination time an indicator of methane production in dairy cows? J. Dairy Sci. 2018, 101, 11074–11085. [Google Scholar] [CrossRef] [Green Version]
- Min, B.-R.; Lee, S.; Jung, H.; Miller, D.N.; Chen, R. Enteric Methane Emissions and Animal Performance in Dairy and Beef Cattle Production: Strategies, Opportunities, and Impact of Reducing Emissions. Animals 2022, 12, 948. [Google Scholar] [CrossRef]
- Popova, M.; Martin, C.; Eugène, M.; Mialon, M.M.; Doreau, M.; Morgavi, D.P. Effect of fibre- and starch-rich finishing diets on methanogenic Archaea diversity and activity in the rumen of feedlot bulls. Anim. Feed Sci. Technol. 2011, 166–167, 113–121. [Google Scholar] [CrossRef] [Green Version]
- Hammond, K.J.; Humphries, D.J.; Crompton, L.A.; Green, C.; Reynolds, C.K. Methane emissions from cattle: Estimates from short-term measurements using a GreenFeed system compared with measurements obtained using respiration chambers or sulphur hexafluoride tracer. Anim. Feed Sci. Technol. 2015, 203, 41–52. [Google Scholar] [CrossRef]
- Trupa, A.; Aplocina, E.; Degola, L. Forage quality and feed intake effect on methane emissions from dairy farming. Eng. Rural. Dev. 2015, 20, 601–605. [Google Scholar]
- Van Gastelen, S.; Antunes-Fernandes, E.C.; Hettinga, K.A.; Klop, G.; Alferink, S.J.J.; Hendriks, W.H.; Dijkstra, J. Enteric methane production, rumen volatile fatty acid concentrations, and milk fatty acid composition in lactating Holstein-Friesian cows fed grass silage- or corn silage-based diets. J. Dairy Sci. 2015, 98, 1915–1927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguerre, M.J.; Wattiaux, M.A.; Powell, J.M.; Broderick, G.A.; Arndt, C. Effect of forage-to-concentrate ratio in dairy cow diets on emission of methane, carbon dioxide, and ammonia, lactation performance, and manure excretion. J. Dairy Sci. 2011, 94, 3081–3093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machado, L.; Magnusson, M.; Paul, N.A.; Kinley, R.; de Nys, R.; Tomkins, N. Identification of bioactives from the red seaweed Asparagopsis taxiformis that promote antimethanogenic activity in vitro. J. Appl. Phycol. 2016, 28, 3117–3126. [Google Scholar] [CrossRef]
- Muizelaar, W.; Groot, M.; van Duinkerken, G.; Peters, R.; Dijkstra, J. Safety and Transfer Study: Transfer of Bromoform Present in Asparagopsis taxiformis to Milk and Urine of Lactating Dairy Cows. Foods 2021, 10, 584. [Google Scholar] [CrossRef]
- Kinley, R.D.; Martinez-Fernandez, G.; Matthews, M.K.; de Nys, R.; Magnusson, M.; Tomkins, N.W. Mitigating the carbon footprint and improving productivity of ruminant livestock agriculture using a red seaweed. J. Clean. Prod. 2020, 259, 120836. [Google Scholar] [CrossRef]
- Bhuyan, M.; Islam, A.; Sayeed, M.; Sharif, M.; Islam, M. Seaweed: A Powerful Tool for Climate Change Mitigation That Provides Various Ecological Services; Springer: Berlin/Heidelberg, Germany, 2021; pp. 159–191. [Google Scholar]
- Duarte, C.M.; Wu, J.; Xiao, X.; Bruhn, A.; Krause-Jensen, D. Can Seaweed Farming Play a Role in Climate Change Mitigation and Adaptation? Front. Mar. Sci. 2017, 4, 100. [Google Scholar] [CrossRef] [Green Version]
- Langton, R.; Augyte, S.; Price, N.; Forster, J.; Noji, T.; Grebe, G.; Gelais, A.; Byron, C.J. An Ecosystem Approach to the Culture of Seaweed; National Oceanic and Atmospheric Administration: Washington, DC, USA, 2019.
- Ku-Vera, J.C.; Jiménez-Ocampo, R.; Valencia-Salazar, S.S.; Montoya-Flores, M.D.; Molina-Botero, I.C.; Arango, J.; Gómez-Bravo, C.A.; Aguilar-Pérez, C.F.; Solorio-Sánchez, F.J. Role of Secondary Plant Metabolites on Enteric Methane Mitigation in Ruminants. Front. Vet. Sci. 2020, 7, 584. [Google Scholar] [CrossRef]
- Wanapat, M.; Chanthakhoun, V.; Phesatcha, K.; Kang, S. Influence of mangosteen peel powder as a source of plant secondary compounds on rumen microorganisms, volatile fatty acids, methane and microbial protein synthesis in swamp buffaloes. Livest. Sci. 2014, 162, 126–133. [Google Scholar] [CrossRef]
- Anantasook, N.; Wanapat, M.; Cherdthong, A. Manipulation of ruminal fermentation and methane production by supplementation of rain tree pod meal containing tannins and saponins in growing dairy steers. J. Anim. Physiol. Anim. Nutr. 2014, 98, 50–55. [Google Scholar] [CrossRef]
- Abdalla, A.L.; Louvandini, H.; Sallam, S.M.A.H.; Bueno, I.C.; Tsai, S.M.; Figueira, A.V. In vitro evaluation, in vivo quantification, and microbial diversity studies of nutritional strategies for reducing enteric methane production. Trop. Anim. Health Prod. 2012, 44, 953–964. [Google Scholar] [CrossRef]
- Cardoso-Gutierrez, E.; Aranda-Aguirre, E.; Robles-Jimenez, L.E.; Castelán-Ortega, O.A.; Chay-Canul, A.J.; Foggi, G.; Angeles-Hernandez, J.C.; Vargas-Bello-Pérez, E.; González-Ronquillo, M. Effect of tannins from tropical plants on methane production from ruminants: A systematic review. Vet. Anim. Sci. 2021, 14, 100214. [Google Scholar] [CrossRef]
- Piluzza, G.; Sulas, L.; Bullitta, S. Tannins in forage plants and their role in animal husbandry and environmental sustainability: A review. Grass Forage Sci. 2013, 69, 32–48, Epub ahead of print 1 March 2014. [Google Scholar] [CrossRef]
- Shirmohammadli, Y.; Efhamisisi, D.; Pizzi, A.P. Tannins as a sustainable raw material for green chemistry: A review. Ind. Crops Prod. 2018, 126, 316–332. [Google Scholar] [CrossRef]
- Jayanegara, A.; Wina, E.; Takahashi, J. Meta-analysis on Methane Mitigating Properties of Saponin-rich Sources in the Rumen: Influence of Addition Levels and Plant Sources. Asian Australas. J. Anim. Sci. 2014, 27, 1426–1435. [Google Scholar] [CrossRef]
- Wanapat, M.; Kang, S.; Polyorach, S. Development of feeding systems and strategies of supplementation to enhance rumen fermentation and ruminant production in the tropics. J. Anim. Sci. Biotechnol. 2013, 4, 32. [Google Scholar] [CrossRef] [Green Version]
- Jayanegara, A.; Leiber, F.; Kreuzer, M. Meta-analysis of the relationship between dietary tannin level and methane formation in ruminants from in vivo and in vitro experiments. J. Anim. Physiol. Anim. Nutr. 2012, 96, 365–375. [Google Scholar] [CrossRef]
- Rai, S.; Acharya-Siwakoti, E.; Kafle, A.; Devkota, H.P.; Bhattarai, A. Plant-Derived Saponins: A Review of Their Surfactant Properties and Applications. Sci 2021, 3, 44. [Google Scholar] [CrossRef]
- Xu, M.; Wan, Z.; Yang, X. Recent Advances and Applications of Plant-Based Bioactive Saponins in Colloidal Multiphase Food Systems. Molecules 2021, 26, 6075. [Google Scholar] [CrossRef]
- Rossi, C.A.S.; Grossi, S.; Dell’Anno, M.; Compiani, C.; Rossi, L. Effect of a Blend of Essential Oils, Bioflavonoids and Tannins on In Vitro Methane Production and In Vivo Production Efficiency in Dairy Cows. Animals 2022, 12, 728. [Google Scholar] [CrossRef]
- Manh, N.S.; Wanapat, M.; Uriyapongson, S.; Khejornsart, P.; Chanthakhoun, V. Effect of eucalyptus (Camaldulensis) leaf meal powder on rumen fermentation characteristics in cattle fed on rice straw. Afr. J. Agric. Res. 2012, 7, 2142–2148. [Google Scholar]
- Belanche, A.; Newbold, C.J.; Morgavi, D.P.; Bach, A.; Zweifel, B.; Yáñez-Ruiz, D.R. A Meta-analysis Describing the Effects of the Essential oils Blend Agolin Ruminant on Performance, Rumen Fermentation and Methane Emissions in Dairy Cows. Animals 2020, 10, E620. [Google Scholar] [CrossRef]
- Brice, R.M.; Dele, P.A.; Ike, K.A.; Shaw, Y.A.; Olangunju, L.K.; Orimaye, O.E.; Subedi, K.; Anele, U.Y. Effects of Essential Oil Blends on In Vitro Apparent and Truly Degradable Dry Matter, Efficiency of Microbial Production, Total Short-Chain Fatty Acids and Greenhouse Gas Emissions of Two Dairy Cow Diets. Animals 2022, 12, 2185. [Google Scholar] [CrossRef]
- Visch, W.; Kononets, M.; Hall, P.O.J.; Nylund, G.M.; Pavia, H. Environmental impact of kelp (Saccharina latissima) aquaculture. Mar. Pollut. Bull. 2020, 155, 110962. [Google Scholar] [CrossRef]
- Kim, J.; Stekoll, M.; Yarish, C. Opportunities, challenges and future directions of open-water seaweed aquaculture in the United States. Phycologia 2019, 58, 446–461. [Google Scholar] [CrossRef]
- Gerretsen, I. The Remarkable Power of Australian Kelp. Available online: https://www.bbc.com/future/article/20210406-how-kelp-can-help-solve-climate-change (accessed on 14 September 2022).
- Beauchemin, K.A.; Ungerfeld, E.M.; Eckard, R.J.; Wang, M. Review: Fifty years of research on rumen methanogenesis: Lessons learned and future challenges for mitigation. Anim. Int. J. Anim. Biosci. 2020, 14, S2–S16. [Google Scholar] [CrossRef] [Green Version]
- Machado, L.; Kinley, R.D.; Magnusson, M.; de Nys, R.; Tomkins, N.W. The potential of macroalgae for beef production systems in Northern Australia. J. Appl. Phycol. 2015, 27, 2001–2005. [Google Scholar] [CrossRef]
- Stefenoni, H.A.; Räisänen, S.E.; Cueva, S.F.; Wason, D.E.; Lage, C.F.A.; Melgar, A.; Fetter, M.E.; Smith, P.; Hennessy, M.; Vecchiarelli, B.; et al. Effects of the macroalga Asparagopsis taxiformis and oregano leaves on methane emission, rumen fermentation, and lactational performance of dairy cows. J. Dairy Sci. 2021, 104, 4157–4173. [Google Scholar] [CrossRef]
- Roque, B.M.; Salwen, J.K.; Kinley, R.; Kebreab, E. Inclusion of Asparagopsis armata in lactating dairy cows’ diet reduces enteric methane emission by over 50 percent. J. Clean. Prod. 2019, 234, 132–138. [Google Scholar] [CrossRef]
- Li, X.; Norman, H.C.; Kinley, R.D.; Laurence, M.; Wilmot, M.; Bender, H.; Nus, R.; Tomkins, N. Asparagopsis taxiformis decreases enteric methane production from sheep. Anim. Prod. Sci. 2016, 58, 681–688. [Google Scholar] [CrossRef]
- García-Poza, S.; Leandro, A.; Cotas, C.; Cotas, J.; Marques, J.C.; Pereira, L.; Gonçalves, A.M.M. The Evolution Road of Seaweed Aquaculture: Cultivation Technologies and the Industry 4.0. Int. J. Environ. Res. Public Health 2020, 17, 6528. [Google Scholar] [CrossRef] [PubMed]
- Food and Agriculture Organization of the United Nations. Towards Blue Transformation. In The State of World Fisheries and Aquaculture; Aquaculture Production; FAO: Rome, Italy, 2022. [Google Scholar] [CrossRef]
- Nilsson, J.; Martin, M. Exploratory environmental assessment of large-scale cultivation of seaweed used to reduce enteric methane emissions. Sustain. Prod. Consum. 2022, 30, 413–423. [Google Scholar] [CrossRef]
- Cai, J.; Lovatelli, A.; Aguilar-Manjarrez, J.; Cornish, L.; Dabbadie, L.; Desrochers, A.; Diffey, S.; Garrido Gamarro, E.; Geehan, J.; Hurtado, A.; et al. Seaweeds and Microalgae: An Overview for Unlocking Their Potential in Global Aquaculture Development; FAO: Rome, Italy, 2021. [Google Scholar] [CrossRef]
- Charrier, B.; Abreu, M.H.; Araujo, R.; Bruhn, A.; Coates, J.; De Clerck, O.; Katsaros, C.; Robaina, R.R.; Wichard, T. Furthering knowledge of seaweed growth and development to facilitate sustainable aquaculture. New Phytol. 2017, 216, 967–975. [Google Scholar] [CrossRef] [Green Version]
- Duarte, C.M.; Bruhn, A.; Krause-Jensen, D. A seaweed aquaculture imperative to meet global sustainability targets. Nat. Sustain. 2022, 5, 185–193. [Google Scholar] [CrossRef]
- Cobellis, G.; Trabalza, M.; Yu, Z. Critical evaluation of essential oils as rumen modifiers in ruminant nutrition: A review. Sci. Total Environ. 2016, 545–546, 556–568. [Google Scholar] [CrossRef]
- Benetel, G.; dos Silva, T.S.; Fagundes, G.M.; Welter, K.C.; Melo, F.A.; Lobo, A.A.G.; Muir, J.P.; Bueno, I.C.S. Essential Oils as In Vitro Ruminal Fermentation Manipulators to Mitigate Methane Emission by Beef Cattle Grazing Tropical Grasses. Molecules 2022, 27, 2227. [Google Scholar] [CrossRef]
- Fraser, G.R.; Chaves, A.V.; Wang, Y.; McAllister, T.A.; Beauchemin, K.A.; Benchaar, C. Assessment of the Effects of Cinnamon Leaf Oil on Rumen Microbial Fermentation Using Two Continuous Culture Systems1. J. Dairy Sci. 2007, 90, 2315–2328. [Google Scholar] [CrossRef] [Green Version]
- Hodjatpanah-montazeri, A.; Mesgaran, M.; Vakili, A.; Tahmasebi, A.M. Effect of essential oils of various plants as microbial modifier to alter corn silage fermentation and in vitro methane production. Iran. J. Appl. Anim. Sci. 2016, 6, 269–276. [Google Scholar]
- Montoya-Flores, M.D.; Molina-Botero, I.C.; Arango, J.; Romano-Muñoz, J.L.; Solorio-Sánchez, F.; Aguilar-Pérez, C.F.; Ku-Vera, J.C. Effect of Dried Leaves of Leucaena leucocephala on Rumen Fermentation, Rumen Microbial Population, and Enteric Methane Production in Crossbred Heifers. Animals 2020, 10, 300. [Google Scholar] [CrossRef] [Green Version]
- Suybeng, B.; Charmley, E.; Gardiner, C.P.; Malau-Aduli, B.S.; Malau-Aduli, A.E.O. Methane Emissions and the Use of Desmanthus in Beef Cattle Production in Northern Australia. Animals 2019, 9, 542. [Google Scholar] [CrossRef] [Green Version]
- ZhiHua, F.; YuFeng, C.; YanXia, G.; QiuFeng, L.; JianGuo, L. Effect of Gross Saponin of Tribulus terrestris on ruminal fermentation and methane production in vitro. J. Anim. Vet. Adv. 2012, 11, 2121–2125. [Google Scholar]
- Guo, Y.Q.; Liu, J.-X.; Lu, Y.; Zhu, W.Y.; Denman, S.E.; McSweeney, C.S. Effect of tea saponin on methanogenesis, microbial community structure and expression of mcrA gene, in cultures of rumen micro-organisms. Lett. Appl. Microbiol. 2008, 47, 421–426. [Google Scholar] [CrossRef]
- Holtshausen, L.; Chaves, A.V.; Beauchemin, K.A.; McGinn, S.M.; McAllister, T.A.; Odongo, N.E.; Cheeke, P.R.; Benchaar, C. Feeding saponin-containing Yucca schidigera and Quillaja saponaria to decrease enteric methane production in dairy cows. J. Dairy Sci. 2009, 92, 2809–2821. [Google Scholar] [CrossRef]
- Seradj, A.R.; Abecia, L.; Crespo, J.; Villalba, D.; Fondevila, M.; Balcells, J. The effect of Bioflavex® and its pure flavonoid components on in vitro fermentation parameters and methane production in rumen fluid from steers given high concentrate diets. Anim. Feed Sci. Technol. 2014, 197, 85–91. [Google Scholar] [CrossRef] [Green Version]
- Stoldt, A.-K.; Derno, M.; Das, G.; Weitzel, J.M.; Wolffram, S.; Metges, C.C. Effects of rutin and buckwheat seeds on energy metabolism and methane production in dairy cows. J. Dairy Sci. 2016, 99, 2161–2168. [Google Scholar] [CrossRef] [Green Version]
- Cui, K.; Guo, X.D.; Tu, Y.; Zhang, N.F.; Ma, T.; Diao, Q.Y. Effect of dietary supplementation of rutin on lactation performance, ruminal fermentation and metabolism in dairy cows. J. Anim. Physiol. Anim. Nutr. 2015, 99, 1065–1073. [Google Scholar] [CrossRef]
- Marie, A. Cinnamon Benefits + Side Effects. HEALabel. 2022. Available online: https://www.healabel.com/cinnamon-benefits/ (accessed on 14 September 2022).
- Menggala, S.R.; Vanhove, W.; Muhammad, D.R.A.; Hendri, J.; Speelman, S.; Van Dammen, P. Sustainable Harvesting of Cinnamomum burmannii (Nees & T. Nees) Blume in Kerinci Regency, Indonesia. Sustainability 2019, 11, 6709. [Google Scholar]
- Marie, A. Oregano Benefits + Side Effects. HEALabel. 2022. Available online: https://www.healabel.com/oregano-benefits/ (accessed on 14 September 2022).
- Mar’atusholikha, V.; Widiatmaka Firmansyah, I. Sustainability of garlic cultivation at Tegal Regency, Central Java Province, Indonesia. IOP Conf. Ser. Earth Environ. Sci. 2019, 399, 012027. [Google Scholar] [CrossRef]
- Your Bottle of Eucalyptus Oil Is Helping Save Our Planet Here’s How. Bosistos. Available online: https://www.bosistos.com.au/blogs/blog/your-bottle-of-eucalyptus-oil-is-helping-save-our-planet-heres-how (accessed on 14 September 2022).
- ME. The Eco-Friendliness of Eucalyptus Floors: An In-Depth Look. The Greener Living Blog. 2018. Available online: https://www.ambientbp.com/blog/how-eco-friendly-are-eucalyptus-floors (accessed on 14 September 2022).
- Villar, M.L. The Effect of Dietary Nitrate and Canola Oil Alone or In Combination on Fermentation, Digesta Kinetics and Methane Emissions from Cattle. Anim. Feed Sci. Technol. 2019, 259, 114294. [Google Scholar] [CrossRef]
- Bayat, A.R.; Tapio, I.; Vilkki, J.; Shingfield, K.J.; Leskinen, H. Plant oil supplements reduce methane emissions and improve milk fatty acid composition in dairy cows fed grass silage-based diets without affecting milk yield. J. Dairy Sci. 2018, 101, 1136–1151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramin, M.; Chagas, J.C.; Smidt, H.; Exposito, R.G.; Krizsan, S.J. Enteric and Fecal Methane Emissions from Dairy Cows Fed Grass or Corn Silage Diets Supplemented with Rapeseed Oil. Animals 2021, 11, 1322. [Google Scholar] [CrossRef] [PubMed]
- Chagas, J.C.; Ramin, M.; Exposito, R.G.; Smidt, H.; Krizsan, S.J. Effect of a Low-Methane Diet on Performance and Microbiome in Lactating Dairy Cows Accounting for Individual Pre-Trial Methane Emissions. Animals 2021, 11, 2597. [Google Scholar] [CrossRef] [PubMed]
- Poulsen, M.; Schwab, C.; Borg Jensen, B.; Engberg, R.M.; Spang, A.; Canibe, N.; Højberg, O.; Milinovich, G.; Fragner, L.; Schleper, C.; et al. Methylotrophic methanogenic Thermoplasmata implicated in reduced methane emissions from bovine rumen. Nat. Commun. 2013, 4, 1428. [Google Scholar] [CrossRef]
- Bowen, J.M.; Cormican, P.; Lister, S.J.; McCabe, M.S.; Duthie, C.-A.; Roehe, R.; Dewhurst, R.J. Links between the rumen microbiota, methane emissions and feed efficiency of finishing steers offered dietary lipid and nitrate supplementation. PLoS ONE 2020, 15, e0231759. [Google Scholar] [CrossRef]
- Difford, G.F.; Lassen, J.; Løvendahl, P. Interchangeability between methane measurements in dairy cows assessed by comparing precision and agreement of two non-invasive infrared methods. Comput. Electron. Agric. 2016, 124, 220–226. [Google Scholar] [CrossRef] [Green Version]
- Kittelmann, S.; Pinares-Patiño, C.S.; Seedorf, H.; Kirk, M.R.; Ganesh, S.; McEwan, J.C.; Janssen, P.H. Two Different Bacterial Community Types Are Linked with the Low-Methane Emission Trait in Sheep. PLoS ONE 2014, 9, e103171. [Google Scholar] [CrossRef]
- Garnsworthy, P.C.; Craigon, J.; Hernandez-Medrano, J.H.; Saunderns, N. Variation among individual dairy cows in methane measurements made on farm during milking. J. Dairy Sci. 2012, 95, 3181–3189. [Google Scholar] [CrossRef] [Green Version]
- Lassen, J.; Difford, G.F. Review: Genetic and genomic selection as a methane mitigation strategy in dairy cattle. Animal 2020, 14, s473–s483. [Google Scholar] [CrossRef]
- Pszczola, M.; Rzewuska, K.; Mucha, S.; Strabel, T. Heritability of methane emissions from dairy cows over a lactation measured on commercial farms. J. Anim. Sci. 2017, 95, 4813–4819. [Google Scholar] [CrossRef]
- Manzanilla-Pech, C.I.V.; L⊘vendahl, P.; Gordo, D.M.; Difford, G.F.; Pryce, J.E.; Schenkel, F.; Wegmann, S.; Miglior, F.; Chud, T.C.; Moate, P.J.; et al. Breeding for reduced methane emission and feed-efficient Holstein cows: An international response. J. Dairy Sci. 2021, 104, 8983–9001. [Google Scholar] [CrossRef]
- Donoghue, K.A.; Bird-Gardiner, T.; Arthur, P.F. Genetic and phenotypic variance and covariance components for methane emission and postweaning traits in Angus cattle. J. Anim. Sci. 2016, 94, 1438–1445. [Google Scholar] [CrossRef]
- Richardson, C.M.; Nguyen, T.T.T.; Abdelsayed, M.; Moate, P.J.; Williams, S.R.O.; Chud, T.C.S.; Schenkel, F.S.; Goddard, M.E.; van den Berg, I.; Cocks, B.G. Genetic parameters for methane emission traits in Australian dairy cows. J. Dairy Sci. 2021, 104, 539–549. [Google Scholar] [CrossRef]
- Hayes, B.J.; Donoghue, K.A.; Reich, C.M.; Mason, B.A.; Bird-Gardiner, T.; Herd, R.M.; Arthur, P.F. Genomic heritabilities and genomic estimated breeding values for methane traits in Angus cattle. J. Anim Sci. 2016, 94, 902–908. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Method | Short Elucidation |
|---|---|
| Respiration chambers (RC) | There are two types of RC: closed-circuit and open-circuit [25]. While closed-circuit systems are practically never used nowadays, open-circuit chambers are the most often exploited type, with varying degrees of complexity [25,47,48]. Individual animals are typically kept in chambers between 2 to 7 days, and CH4 emissions are estimated based on gas flow and changes in gas concentrations between the air coming in and out [10,25,48,49,50]. The chamber approach is expensive in terms of both investment and labor, and it has been accused of affecting feeding behavior. In trials employing transparent chambers, however, no impacts on dry matter intake (DMI) were observed [49]. However, only one cow may be tested at a time, and each test requires several hours in the respiration chamber, limiting research efficiency [10,18,25]. Almost in all studies, a single gas analyzer was used to measure in and out methane concentrations, often for two or more chambers [10,18,42]. When Garnsworthy et al. compared several different methods of GHG measuring, it was found that respiration chambers were the most accurate method. None of the correlations with other methods exceeded 0.90 [10]. |
| Sulfur hexafluoride (SF6) tracer technique | The method is invasive—the cow must consume a bolus carrying the tracer, and the SF6 tracer is an exceptionally potent greenhouse gas [18,42]. The underlying premise is that the rate of SF6 gas release from the rumen is calculated in order to calculate the CH4 emission measurement [25,51]. The approach is suited for enclosed and free-roaming animals, and it involves inserting a permeation tube with a known SF6 gas release rate into the animal’s reticulorumen [49,52]. A tube hooked to a halter and connected to an evacuated canister worn around the animal’s neck or back is the basic premise behind this technique. Near the animal’s nostrils, the air is taken for testing purposes. It takes roughly 24 h for the canister to fill up between 50% and 70% because of an airflow restriction provided by a capillary tube. Methane emission rates are calculated by multiplying the predetermined SF6 release rate by the canister methane to SF6 concentration ratio [10,25]. The technique is more beneficial when evaluating CH4 emissions from individual animals. Wearing the device and daily handling to change canisters may affect animal behavior and feed intake. The sulfur hexafluoride (SF6) tracer technique is far less intrusive than breathing chambers because cows remain in the herd [42]. |
| Spot sampling technique/ Gas-flux quantification system | The cornerstone of spot sampling approaches is the collection of acceptable short-term breath data for emission measurements. The techniques employ spot measurements of exhaled CH4 during milking or feeding. These procedures are typically automated, noninvasive, and non-intrusive, allowing for a high rate of animal throughput [45,49]. Breath sampling is taken during milking and feeding. The feed bin could be at a concentrate feeding station or an autonomous milking station [18,52,53]. These methods are referred to as “sniffer methods” since they use devices originally developed to detect harmful gas leaks. Air is sampled near the animal’s nostrils using a tube attached to a feed bin and immediately connected to a gas analyzer [10,18]. Methane concentrations measured during a sample visit of 3 to 10 min can be expressed as the overall mean or the mean of eructation peaks. Breath-sampling approaches provide substantial advantages over other methods for large-scale measurement of methane emissions by individual animals [20,25]. Some patented methods, such as the GreenFeed system, work in the same method as sniffer methods [25]. The GF method is based on the idea that a single animal’s daily average CH4 emission can be estimated by combining multiple short-term Methane emission measurements obtained throughout the day [25]. Breath-sampling procedures are noninvasive because animals are unaware of the apparatus and are in their natural environment after it is deployed. Animals continue their normal schedule, which includes milking and feeding; therefore, no animal training, handling, or dietary changes are required. Although more sophisticated gas analyzers are available, the equipment is quite inexpensive, and the operating costs are insignificant [20,45]. |
| Carbon dioxide as a tracer to estimate daily methane emission | The methane and carbon dioxide ratio technique predicts CH4 output by certain species by forecasting carbon emissions and sensing methane and carbon dioxide concentrations [18,53]. This method demands knowledge about the ration’s consumption, energy content, and heat increase [54]. Using the CO2 technique does not consider the difference in CH4 emissions between efficient and inefficient cows; according to Huhtanen et al., Researchers found a strong correlation between the efficiency of low and high-efficiency cows. The technique overestimated the amount of CH4 produced by productive cows while underestimating the amount produced by ineffective cows [55]. Because it is so easy to apply to many animals, the standard error of means can be reduced [25]. |
| Infrared ray spectroscopy, laser technique | Lasers have long been applied for gas detection in environmental monitoring, air quality monitoring, security, and health care [25]. Hand-held gas detectors for remote measurements of column density for methane-containing gases. It is based on infrared (IR) absorption spectroscopy. It uses a collimated semiconductor laser as an excitation source and wavelength modulation spectroscopy’s second harmonic detection to establish a methane concentration measurement [25,56]. Methane concentration measurements are performed manually using a portable instrument around 1–3 m away from the animal. The data acquisition sequence comprises small spans of 2–4 min. The resulting data is a series of peaks representing the animal’s breathing cycle [56,57]. The laser methane detector (LMD) can be used in the animal’s natural environment; however, a constraint is required during the measuring process to ensure accuracy. Results can be affected by factors such as the distance from the animal, the angle of pointing, the animal’s motion and moving direction, the airflow movement, and temperature in the barn since the LMD measures methane in the plume issuing from the animal’s nostrils [58]. |
| Face mask (FM) method | The method for spot samplings of respiratory exchange and CH4 emissions is based on animals trained to remain in sternal recumbency for 30 min measurement periods taken every 2–3 h, with up to 7 measures per day [25]. In terms of assessing gas exchange and changes in the exhaled CH4 concentration, the basis of this method is identical to that of RC. It consists of a mass flow controller, gas sampling unit, and CH4 emission analyzer attached to each face mask. Gas readings are corrected for humidity, lag time, drift, and CH4 emission (mL/min) changes for each period [25,59]. The FM approach is less expensive and simpler than SF6 or RC. Its mobility allows it to measure multiple areas in order to collect CH4 emissions [25,60]. |
| Component | Methane Reducing Effect | Influence on Animal Health | Sustainability |
|---|---|---|---|
| Algae, bromoform | ↓ 45–99% [23,37,98] | Bromoform can be excreted in urine and milk [99]. Weight increase was observed [23,100]. | Seaweed raises water pH, hence mitigating ocean acidification (suitable habitat). Emits trace that degrades the ozone layer, dampens wave energy during storms, protects the coast, offers human consumption with biofuels, fertilizer, medicine, and food, animal food supplements [37,101,102,103]. |
| Tannins | ↓ 13–30% [104] | Increase total bacteria and fungi, decrease protozoa, and decrease methanogens [104,105,106,107]. Some decrease fungi but increase methanogens [108]. It can increase weight and production [109]. | They are abundant in many plant species and may be extracted using simple procedures [109,110]. |
| Saponins | ↓ 7–23% [111] | Reduced populations of ruminal ciliate protozoa may accelerate microbial protein flow from the rumen, increasing feed utilization efficiency and decreasing methanogenesis [112]. Decrease protozoa, decrease methanogens [113]. | They are eco-friendly due to their natural nature, biodegradable, and non-toxic, which is critical from an environmental and health standpoint. Saponins obtained from plants can be a sustainable alternative to synthetic surfactants [114,115]. |
| Essential oils | ↓ 8–22% [116] | Alternative feed booster, preserving mineral digestion [117]. Increasing MY, DMI, can improve milk fat and protein composition and decrease somatic cell count [116,118]. Improve the efficiency of microbial production [119]. | When highly concentrated essential oils are used correctly, they may be both ecologically and economically sustainable. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bačėninaitė, D.; Džermeikaitė, K.; Antanaitis, R. Global Warming and Dairy Cattle: How to Control and Reduce Methane Emission. Animals 2022, 12, 2687. https://doi.org/10.3390/ani12192687
Bačėninaitė D, Džermeikaitė K, Antanaitis R. Global Warming and Dairy Cattle: How to Control and Reduce Methane Emission. Animals. 2022; 12(19):2687. https://doi.org/10.3390/ani12192687
Chicago/Turabian StyleBačėninaitė, Dovilė, Karina Džermeikaitė, and Ramūnas Antanaitis. 2022. "Global Warming and Dairy Cattle: How to Control and Reduce Methane Emission" Animals 12, no. 19: 2687. https://doi.org/10.3390/ani12192687