
B Chromosomes in Psalidodon scabripinnis (Characiformes, Characidae) Species Complex

 , , ,
, , ,  , , and
, , and
Abstract
:Simple Summary
Abstract
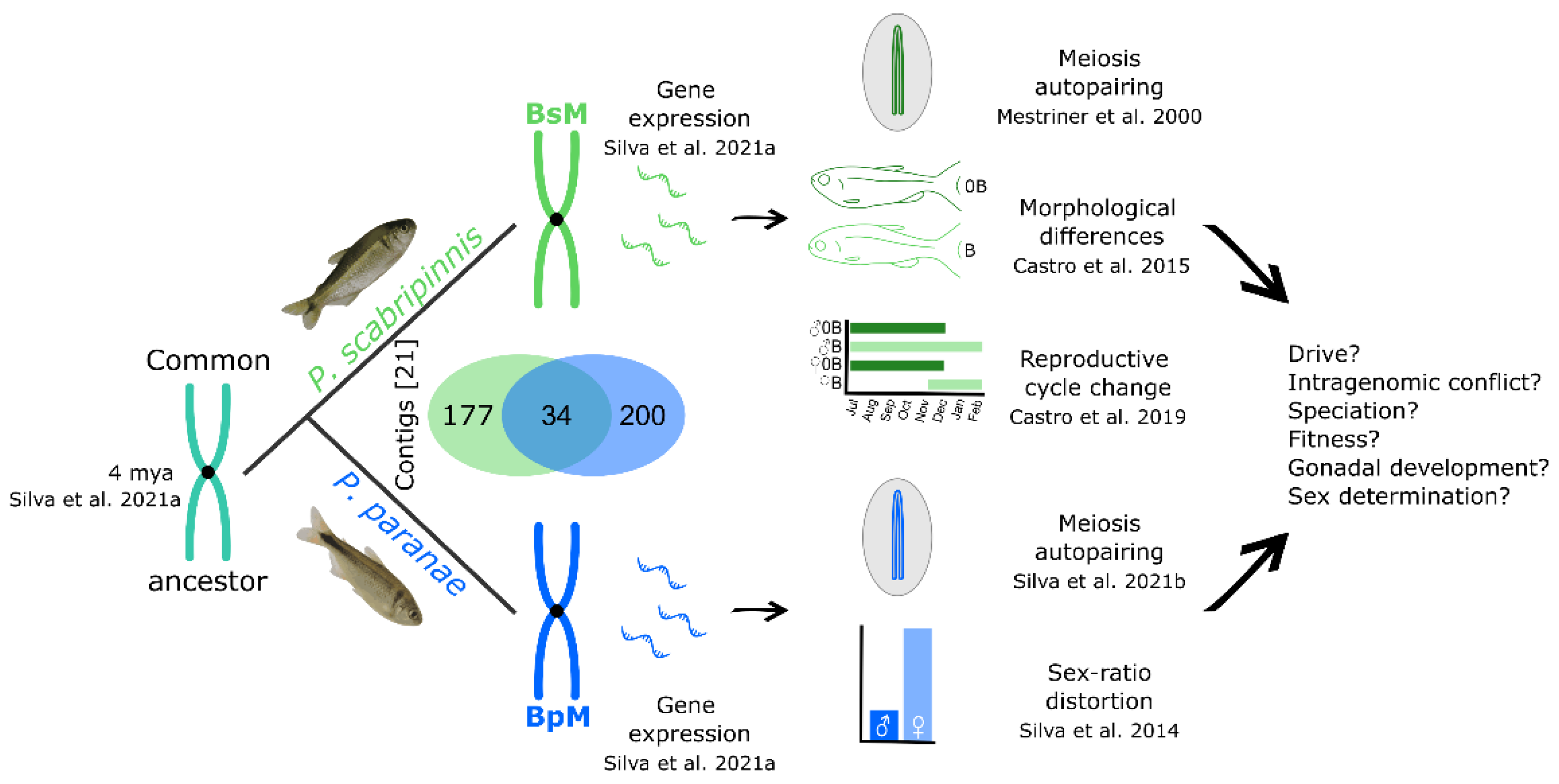
1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
2. Materials and Methods
3. B Chromosomes in Psalidodon scabripinnis Complex
3.1. Origin
3.2. Predominance in Females
3.3. Geographic Variation
4. Transmission of B Chromosomes
5. Structure of B Chromosomes
6. B-Genes
7. Expression and Effects of B-Genes
8. Meiotic Behavior of B Chromosomes
9. B Chromosome and Speciation
10. Psalidodon scabripinnis as a Study Model
11. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Darlington, C.D.; Upcott, M.B. The Activity of Inert Chromosomes in Zea mays. J. Genet. 1941, 41, 275–296. [Google Scholar] [CrossRef]
- Artoni, R.F.; Vicari, M.R.; Endler, A.L.; Cavallaro, Z.I.; de Jesus, C.M.; de Almeida, M.C.; Moreira-Filho, O.; Bertollo, L.A.C. Banding Pattern of A and B Chromosomes of Prochilodus lineatus (Characiformes, Prochilodontidae), with Comments on B Chromosomes Evolution. Genetica 2006, 127, 277. [Google Scholar] [CrossRef] [PubMed]
- Pansonato-Alves, J.C.; Serrano, É.A.; Utsunomia, R.; Camacho, J.P.M.; Silva, G.J.d.C.; Vicari, M.R.; Artoni, R.F.; Oliveira, C.; Foresti, F. Single Origin of Sex Chromosomes and Multiple Origins of B Chromosomes in Fish Genus Characidium. PLoS ONE 2014, 9, e107169. [Google Scholar] [CrossRef] [PubMed]
- McAllister, B.F.; Werren, J.H. Hybrid Origin of a B Chromosome (PSR) in the Parasitic Wasp Nasonia vitripennis. Chromosoma 1997, 106, 243–253. [Google Scholar] [CrossRef]
- Tosta, V.C.; Marthe, J.B.; Tavares, M.G.; Fernandes-Salomão, T.M.; Pompolo, S.G.; Recco-Pimentel, S.M.; Perfectti, F.; Campos, L.A.O.; Camacho, J.P.M. Possible Introgression of B Chromosomes between Bee Species (Genus Partamona). Cytogenet. Genome Res. 2014, 144, 220–226. [Google Scholar] [CrossRef]
- Camacho, J.P.M.; Sharbel, T.F.; Beukeboom, L.W. B-Chromosome Evolution. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2000, 355, 163–178. [Google Scholar] [CrossRef]
- Dos Santos, L.P.; Francisco, C.M.; Júnior, E.O.C.; Castro, J.P.; Utsunomia, R.; Morelli, S.; Porto-Foresti, F.; Foresti, F.; Artoni, R.F. Chromosomal Instability and Origin of B Chromosomes in the Amazonian Glass Tetra Moenkhausia oligolepis (Günther, 1864) (Characiformes, Characidae). Cytogenet. Genome Res. 2021, 161, 249–256. [Google Scholar] [CrossRef]
- Silva, D.M.Z.A.; Pansonato-Alves, J.C.; Utsunomia, R.; Araya-Jaime, C.; Ruiz-Ruano, F.J.; Daniel, S.N.; Hashimoto, D.T.; Oliveira, C.; Camacho, J.P.M.; Porto-Foresti, F.; et al. Delimiting the Origin of a B Chromosome by FISH Mapping, Chromosome Painting and DNA Sequence Analysis in Astyanax paranae (Teleostei, Characiformes). PLoS ONE 2014, 9, e94896. [Google Scholar] [CrossRef]
- Serrano, É.A.; Utsunomia, R.; Scudeller, P.S.; Oliveira, C.; Foresti, F. Origin of B Chromosomes in Characidium alipioi (Characiformes, Crenuchidae) and Its Relationship with Supernumerary Chromosomes in Other Characidium Species. Comp. Cytogenet. 2017, 11, 81–95. [Google Scholar] [CrossRef]
- Serrano-Freitas, É.A.; Silva, D.M.Z.A.; Ruiz-Ruano, F.J.; Utsunomia, R.; Araya-Jaime, C.; Oliveira, C.; Camacho, J.P.M.; Foresti, F. Satellite DNA Content of B Chromosomes in the Characid Fish Characidium gomesi Supports Their Origin from Sex Chromosomes. Mol. Genet. Genom. 2020, 295, 195–207. [Google Scholar] [CrossRef]
- Blavet, N.; Yang, H.; Su, H.; Solanský, P.; Douglas, R.N.; Karafiátová, M.; Šimková, L.; Zhang, J.; Liu, Y.; Hou, J.; et al. Sequence of the Supernumerary B Chromosome of Maize Provides Insight into Its Drive Mechanism and Evolution. Proc. Natl. Acad. Sci. USA 2021, 118, e2104254118. [Google Scholar] [CrossRef] [PubMed]
- Banaei-Moghaddam, A.M.; Meier, K.; Karimi-Ashtiyani, R.; Houben, A. Formation and Expression of Pseudogenes on the B Chromosome of Rye. Plant Cell 2013, 25, 2536–2544. [Google Scholar] [CrossRef] [PubMed]
- Valente, G.T.; Conte, M.A.; Fantinatti, B.E.A.; Cabral-de-Mello, D.C.; Carvalho, R.F.; Vicari, M.R.; Kocher, T.D.; Martins, C. Origin and Evolution of B Chromosomes in the Cichlid Fish Astatotilapia latifasciata Based on Integrated Genomic Analyses. Mol. Biol. Evol. 2014, 31, 2061–2072. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Ruano, F.J.; Navarro-Domínguez, B.; López-León, M.D.; Cabrero, J.; Camacho, J.P.M. Evolutionary Success of a Parasitic B Chromosome Rests on Gene Content. bioRxiv 2019, 683417. [Google Scholar] [CrossRef]
- Martis, M.M.; Klemme, S.; Banaei-Moghaddam, A.M.; Blattner, F.R.; Macas, J.; Schmutzer, T.; Scholz, U.; Gundlach, H.; Wicker, T.; Šimková, H.; et al. Selfish Supernumerary Chromosome Reveals Its Origin as a Mosaic of Host Genome and Organellar Sequences. Proc. Natl. Acad. Sci. USA 2012, 109, 13343–13346. [Google Scholar] [CrossRef] [PubMed]
- Houben, A.; Banaei-Moghaddam, A.M.; Klemme, S.; Timmis, J.N. Evolution and Biology of Supernumerary B Chromosomes. Cell. Mol. Life Sci. 2014, 71, 467–478. [Google Scholar] [CrossRef]
- Lui, R.L.; Traldi, J.B.; Blanco, D.R.; Margarido, V.P.; Mariotto, S.; Centofante, L.; Artoni, R.F.; Moreira Filho, O. Possible Common Origin of B Chromosomes in Neotropical Fish (Siluriformes, Auchenipteridae) Reinforced by Repetitive DNA Mapping. Braz. Arch. Biol. Technol. 2021, 64, e21190494. [Google Scholar] [CrossRef]
- Nascimento, C.N.; Troy, W.P.; Alves, J.C.P.; Carvalho, M.L.; Oliveira, C.; Foresti, F. Molecular Cytogenetic Analyses Reveal Extensive Chromosomal Rearrangements and Novel B Chromosomes in Moenkhausia (Teleostei, Characidae). Genet. Mol. Biol. 2020, 43, e20200027. [Google Scholar] [CrossRef]
- Stornioli, J.H.F.; Goes, C.A.G.; Calegari, R.M.; dos Santos, R.Z.; Giglio, L.M.; Foresti, F.; Oliveira, C.; Penitente, M.; Porto-Foresti, F.; Utsunomia, R. The B Chromosomes of Prochilodus lineatus (Teleostei, Characiformes) Are Highly Enriched in Satellite DNAs. Cells 2021, 10, 1527. [Google Scholar] [CrossRef]
- Milani, D.; Ruiz-Ruano, F.J.; Camacho, J.P.M.; Cabral-de-Mello, D.C. Out of Patterns, the Euchromatic B Chromosome of the Grasshopper Abracris flavolineata Is Not Enriched in High-Copy Repeats. Heredity 2021, 127, 475–483. [Google Scholar] [CrossRef]
- Silva, D.M.Z.A.; Ruiz-Ruano, F.J.; Utsunomia, R.; Martín-Peciña, M.; Castro, J.P.; Freire, P.P.; Carvalho, R.F.; Hashimoto, D.T.; Suh, A.; Oliveira, C.; et al. Long-Term Persistence of Supernumerary B Chromosomes in Multiple Species of Astyanax Fish. BMC Biol. 2021, 19, 52. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Domínguez, B.; Ruiz-Ruano, F.J.; Cabrero, J.; Corral, J.M.; López-León, M.D.; Sharbel, T.F.; Camacho, J.P.M. Protein-Coding Genes in B Chromosomes of the Grasshopper Eyprepocnemis plorans. Sci. Rep. 2017, 7, 45200. [Google Scholar] [CrossRef] [PubMed]
- Dalla Benetta, E.; Akbari, O.S.; Ferree, P.M. Sequence Expression of Supernumerary B Chromosomes: Function or Fluff? Genes 2019, 10, 123. [Google Scholar] [CrossRef] [PubMed]
- Kinsella, C.M.; Ruiz-Ruano, F.J.; Dion-Côté, A.-M.; Charles, A.J.; Gossmann, T.I.; Cabrero, J.; Kappei, D.; Hemmings, N.; Simons, M.J.P.; Camacho, J.P.M.; et al. Programmed DNA Elimination of Germline Development Genes in Songbirds. Nat. Commun. 2019, 10, 5468. [Google Scholar] [CrossRef]
- Ma, W.; Gabriel, T.S.; Martis, M.M.; Gursinsky, T.; Schubert, V.; Vrána, J.; Doležel, J.; Grundlach, H.; Altschmied, L.; Scholz, U.; et al. Rye B Chromosomes Encode a Functional Argonaute-like Protein with in Vitro Slicer Activities Similar to Its A Chromosome Paralog. New Phytol. 2017, 213, 916–928. [Google Scholar] [CrossRef]
- Dalla Benetta, E.; Antoshechkin, I.; Yang, T.; Nguyen, H.Q.M.; Ferree, P.M.; Akbari, O.S. Genome Elimination Mediated by Gene Expression from a Selfish Chromosome. Sci. Adv. 2020, 6, eaaz9808. [Google Scholar] [CrossRef]
- Yoshida, K.; Terai, Y.; Mizoiri, S.; Aibara, M.; Nishihara, H.; Watanabe, M.; Kuroiwa, A.; Hirai, H.; Hirai, Y.; Matsuda, Y.; et al. B Chromosomes Have a Functional Effect on Female Sex Determination in Lake Victoria Cichlid Fishes. PLoS Genet. 2011, 7, e1002203. [Google Scholar] [CrossRef]
- Clark, F.E.; Kocher, T.D. Changing Sex for Selfish Gain: B Chromosomes of Lake Malawi Cichlid Fish. Sci. Rep. 2019, 9, 20213. [Google Scholar] [CrossRef]
- Ruban, A.; Schmutzer, T.; Wu, D.D.; Fuchs, J.; Boudichevskaia, A.; Rubtsova, M.; Pistrick, K.; Melzer, M.; Himmelbach, A.; Schubert, V.; et al. Supernumerary B Chromosomes of Aegilops speltoides Undergo Precise Elimination in Roots Early in Embryo Development. Nat. Commun. 2020, 11, 2764. [Google Scholar] [CrossRef]
- Vea, I.M.; de la Filia, A.G.; Jaron, K.S.; Mongue, A.J.; Ruiz-Ruano, F.J.; Barlow, S.E.J.; Nelson, R.; Ross, L. The B Chromosome of Pseudococcus viburni: A Selfish Chromosome That Exploits Whole-Genome Meiotic Drive. bioRxiv 2021. [Google Scholar] [CrossRef]
- D’Ambrosio, U.; Alonso-Lifante, M.P.; Barros, K.; Kovařík, A.; Mas de Xaxars, G.; Garcia, S. B-Chrom: A Database on B-Chromosomes of Plants, Animals and Fungi. New Phytol. 2017, 216, 635–642. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, R.A.; Martins-Santos, I.C.; Dias, A.L. B Chromosomes: An Update about Their Occurrence in Freshwater Neotropical Fishes (Teleostei). J. Fish Biol. 2008, 72, 1907–1932. [Google Scholar] [CrossRef]
- Oliveira, C.; Foresti, F.; Hilsdorf, A.W.S. Genetics of Neotropical Fish: From Chromosomes to Populations. Fish Physiol. Biochem. 2009, 35, 81–100. [Google Scholar] [CrossRef] [PubMed]
- Moreira-Filho, O.; Galetti Jr, P.M.; Bertollo, L.A.C. B Chromosomes in the Fish Astyanax scabripinnis (Characidae, Tetragonopterinae): An Overview in Natural Populations. Cytogenet. Genome Res. 2004, 106, 230–234. [Google Scholar] [CrossRef] [PubMed]
- Daniel, S.N.; Hashimoto, D.T.; Pansonato-Alves, J.C.; Foresti, F.; Porto-Foresti, F. Cytogenetic Characterization of Distinct B Chromosomes in a Population of the Fish Astyanax bockmanni (Teleostei, Characiformes). Caryologia 2012, 65, 229–233. [Google Scholar] [CrossRef]
- Moreira-Filho, O.; Bertollo, L.A.C. Astyanax scabripinnis (Pisces; Characidae): A “Species Complex”. Braz. J. Genet. 1991, 14, 331–357. [Google Scholar]
- Terán, G.E.; Benitez, M.F.; Mirande, J.M. Opening the Trojan Horse: Phylogeny of Astyanax, Two New Genera and Resurrection of Psalidodon (Teleostei: Characidae). Zool. J. Linn. Soc. 2020, 190, 1217–1234. [Google Scholar] [CrossRef]
- Mestriner, C.A.; Galetti, P.M., Jr.; Valentini, S.R.; Ruiz, I.R.G.; Abel, L.D.S.; Moreira-Filho, O.; Camacho, J.P.M. Structural and Functional Evidence That a B Chromosome in the Characid Fish Astyanax scabripinnis Is an Isochromosome. Heredity 2000, 85, 1–9. [Google Scholar] [CrossRef]
- Silva, D.M.Z.A.; Araya-Jaime, C.; Yamashita, M.; Vidal, M.R.; Oliveira, C.; Porto-Foresti, F.; Artoni, R.F.; Foresti, F. Meiotic Self-Pairing of the Psalidodon (Characiformes, Characidae) Iso-B Chromosome: A Successful Perpetuation Mechanism. Genet. Mol. Biol. 2021, 44, e20210084. [Google Scholar] [CrossRef]
- Castro, J.P.; Hattori, R.S.; Yoshinaga, T.T.; Silva, D.M.Z.A.; Ruiz-Ruano, F.J.; Foresti, F.; Santos, M.H.; de Almeida, M.C.; Moreira-Filho, O.; Artoni, R.F. Differential Expression of Genes Related to Sexual Determination Can Modify the Reproductive Cycle of Astyanax scabripinnis (Characiformes: Characidae) in B Chromosome Carrier Individuals. Genes 2019, 10, 909. [Google Scholar] [CrossRef]
- Castro, J.P.; Moura, M.O.; Moreira-Filho, O.; Shibatta, O.A.; Santos, M.H.; Nogaroto, V.; Vicari, M.R.; de Almeida, M.C.; Artoni, R.F. Diversity of the Astyanax scabripinnis Species Complex (Teleostei: Characidae) in the Atlantic Forest, Brazil: Species Limits and Evolutionary Inferences. Rev. Fish Biol. Fish. 2015, 25, 231–244. [Google Scholar] [CrossRef]
- Salvador, L.B.; Moreira-Filho, O. B Chromosomes in Astyanax scabripinnis (Pisces, Characidae). Heredity 1992, 69, 50–56. [Google Scholar] [CrossRef]
- Vicente, V.E.; Moreira-Filho, O.; Camacho, J.P.M. Sex-Ratio Distortion Associated with the Presence of a B Chromosome in Astyanax scabripinnis (Teleostei, Characidae). Cytogenet. Genome Res. 1996, 74, 70–75. [Google Scholar] [CrossRef] [PubMed]
- Maistro, E.L.; Foresti, F.; Oliveira, C. R- and G-Band Patterns in Astyanax scabripinnis paranae (Pisces, Characiformes, Characidae). Genet. Mol. Biol. 1999, 22, 201–204. [Google Scholar] [CrossRef]
- Maistro, E.L.; Oliveira, C.; Foresti, F. Cytogenetic Characterization of a Supernumerary Chromosome Segment and of B-Chromosomes in Astyanax scabripinnis (Teleostei, Characidae). Genetica 2000, 110, 177. [Google Scholar] [CrossRef]
- Silva, D.M.Z.A.; Utsunomia, R.; Ruiz-Ruano, F.J.; Daniel, S.N.; Porto-Foresti, F.; Hashimoto, D.T.; Oliveira, C.; Camacho, J.P.M.; Foresti, F. High-Throughput Analysis Unveils a Highly Shared Satellite DNA Library among Three Species of Fish Genus Astyanax. Sci. Rep. 2017, 7, 12726. [Google Scholar] [CrossRef]
- Néo, D.M.; Bertollo, L.A.C.; Filho, O.M. Morphological Differentiation and Possible Origin of B Chromosomes in Natural Brazilian Population of Astyanax scabripinnis (Pisces, Characidae). Genetica 2000, 108, 211–215. [Google Scholar] [CrossRef]
- Ferro, D.A.M.; Moreira-Filho, O.; Bertollo, L.A.C. B Chromosome Polymorphism in the Fish, Astyanax scabripinnis. Genetica 2003, 119, 147–153. [Google Scholar] [CrossRef]
- Rocon-Stange, E.A.; Almeida-Toledo, L.F. Supernumerary B Chromosomes Restricted to Males in Asyanax scabripinnis (Pisces, Characidae). Braz. J. Genet. 1993, 16, 601–615. [Google Scholar]
- Imarazene, B.; Du, K.; Beille, S.; Jouanno, E.; Feron, R.; Pan, Q.; Torres-Paz, J.; Lopez-Roques, C.; Castinel, A.; Gil, L.; et al. A Supernumerary “B-Sex” Chromosome Drives Male Sex Determination in the Pachón Cavefish, Astyanax mexicanus. Curr. Biol. 2021, 31, 4800–4809.e9. [Google Scholar] [CrossRef]
- Maistro, E.L.; Foresti, F.; Oliveira, C. New Occurrence of a Macro B-Chromosome in Astyanax scabripinnis paranae (Pisces, Characiformes, Characidae). Braz. J. Genet. 1994, 17, 153–156. [Google Scholar]
- Porto-Foresti, F.; Oliveira, C.; Maistro, E.L.; Foresti, F. Estimated Frequency of B-Chromosomes and Population Density of Astyanax scabripinnis paranae in a Small Stream. Braz. J. Genet. 1997, 20, 377–380. [Google Scholar] [CrossRef]
- Goes, C.A.G.; Silva, D.M.Z.A.; Utsunomia, R.; do Nascimento, N.F.; Yasui, G.S.; Senhorini, J.A.; Hashimoto, D.T.; Artoni, R.F.; Foresti, F.; Porto-Foresti, F. Sex-Dependent Inheritance of B Chromosomes in Psalidodon paranae (Teleostei, Characiformes) Revealed by Directed Crossings. Zebrafish 2021, 18, 363–368. [Google Scholar] [CrossRef] [PubMed]
- Néo, D.M.; Filho, O.M.; Camacho, J.P.M. Altitudinal Variation for B Chromosome Frequency in the Characid Fish Astyanax scabripinnis. Heredity 2000, 85, 136–141. [Google Scholar] [CrossRef] [PubMed]
- Caramaschi, E.P. Distribuição da Ictiofauna de Riachos das Bacias do Tietê e do Paranapanema, Junto Ao Divisor de Água (Botucatu, SP). Ph.D. Thesis, Universidade Fedral Sao Carlos, São Paulo, Brazil, 1986. [Google Scholar]
- Santos, N.M.; Ferreira-Neto, M.; Artoni, R.F.; Vicari, M.R.; Bakkali, M.; de Oliveira, C.; Foresti, F. A Comparative Structural Cytogenetic Study in Three Allopatric Populations of Astyanax scabripinnis (Teleostei: Characidae). Zool. Curitiba 2012, 29, 159–166. [Google Scholar] [CrossRef]
- Schemczssen-Graeff, Z.; Barbosa, P.; Castro, J.P.; Silva, M.; de Almeida, M.C.; Moreira-Filho, O.; Artoni, R.F. Dynamics of Replication and Nuclear Localization of the B Chromosome in Kidney Tissue Cells in Astyanax scabripinnis (Teleostei: Characidae). Zebrafish 2020, 17, 147–152. [Google Scholar] [CrossRef]
- Cabrero, J.; Martín-Peciña, M.; Ruiz-Ruano, F.J.; Gómez, R.; Camacho, J.P.M. Post-Meiotic B Chromosome Expulsion, during Spermiogenesis, in Two Grasshopper Species. Chromosoma 2017, 126, 633–644. [Google Scholar] [CrossRef]
- Gernand, D.; Rutten, T.; Varshney, A.; Rubtsova, M.; Prodanovic, S.; Brüß, C.; Kumlehn, J.; Matzk, F.; Houben, A. Uniparental Chromosome Elimination at Mitosis and Interphase in Wheat and Pearl Millet Crosses Involves Micronucleus Formation, Progressive Heterochromatinization, and DNA Fragmentation. Plant Cell 2005, 17, 2431–2438. [Google Scholar] [CrossRef]
- Clark, F.E.; Akera, T. Unravelling the Mystery of Female Meiotic Drive: Where We Are. Open Biol. 2021, 11, 210074. [Google Scholar] [CrossRef]
- Raemaekers, T.; Ribbeck, K.; Beaudouin, J.; Annaert, W.; Van Camp, M.; Stockmans, I.; Smets, N.; Bouillon, R.; Ellenberg, J.; Carmeliet, G. NuSAP, a Novel Microtubule-Associated Protein Involved in Mitotic Spindle Organization. J. Cell Biol. 2003, 162, 1017–1029. [Google Scholar] [CrossRef]
- Nie, J.; Wang, H.; He, F.; Huang, H. Nusap1 Is Essential for Neural Crest Cell Migration in Zebrafish. Protein Cell 2010, 1, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Qian, Z.; Li, Y.; Ma, J.; Xue, Y.; Xi, Y.; Hong, L.; Dai, X.; Zhang, Y.; Ji, X.; Chen, Y.; et al. Prognostic Value of NUSAP1 in Progression and Expansion of Glioblastoma Multiforme. J. Neuro-Oncol. 2018, 140, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Akera, T.; Trimm, E.; Lampson, M.A. Molecular Strategies of Meiotic Cheating by Selfish Centromeres. Cell 2019, 178, 1132–1144.e10. [Google Scholar] [CrossRef] [PubMed]
- Vicari, M.R.; de Mello Pistune, H.F.; Castro, J.P.; de Almeida, M.C.; Bertollo, L.A.C.; Moreira-Filho, O.; Camacho, J.P.M.; Artoni, R.F. New Insights on the Origin of B Chromosomes in Astyanax scabripinnis Obtained by Chromosome Painting and FISH. Genetica 2011, 139, 1073. [Google Scholar] [CrossRef] [PubMed]
- Silva, D.M.Z.A.; Daniel, S.N.; Camacho, J.P.M.; Utsunomia, R.; Ruiz-Ruano, F.J.; Penitente, M.; Pansonato-Alves, J.C.; Hashimoto, D.T.; Oliveira, C.; Porto-Foresti, F.; et al. Origin of B Chromosomes in the Genus Astyanax (Characiformes, Characidae) and the Limits of Chromosome Painting. Mol. Genet. Genom. MGG 2016, 291, 1407–1418. [Google Scholar] [CrossRef]
- Fernandes, C.A.; Martins-Santos, I.C. Chromosomal Location of 5S and 18S RRNA Genes in Three Sympatric Cytotypes of Astyanax scabripinnis (Characiformes, Characidae) from the Ivaí River Basin, State of Paraná, Brazil. Caryologia 2006, 59, 253–259. [Google Scholar] [CrossRef]
- Ferro, D.A.M.; Néo, D.M.; Moreira-Filho, O.; Bertollo, L.A.C. Nucleolar Organizing Regions, 18S and 5S RDNA in Astyanax scabripinnis (Pisces, Characidae): Populations Distribution and Functional Diversity. Genetica 2000, 110, 55. [Google Scholar] [CrossRef]
- Barbosa, P.; Leal, E.V.; Silva, M.; de Almeida, M.C.; Moreira-Filho, O.; Artoni, R.F. Variability and Evolutionary Implications of Repetitive DNA Dynamics in Genome of Astyanax scabripinnis (Teleostei, Characidae). Comp. Cytogenet. 2017, 11, 143–162. [Google Scholar] [CrossRef]
- Garrido-Ramos, M.A. Satellite DNA: An Evolving Topic. Genes 2017, 8, 230. [Google Scholar] [CrossRef]
- Carmello, B.O.; Coan, R.L.B.; Cardoso, A.L.; Ramos, E.; Fantinatti, B.E.A.; Marques, D.F.; Oliveira, R.A.; Valente, G.T.; Martins, C. The HnRNP Q-like Gene Is Retroinserted into the B Chromosomes of the Cichlid Fish Astatotilapia latifasciata. Chromosome Res. 2017, 25, 277–290. [Google Scholar] [CrossRef]
- Jones, R.N.; Rees, H. B Chromosomes; Academic Press: London, UK, 1982. [Google Scholar]
- Burt, A.; Trivers, R. Genes in Conflict: The Biology of Selfish Genetic Elements; Harvard University Press: Cambridge, MA, USA, 2009; ISBN 978-0-674-02911-8. [Google Scholar]
- Castro, J.P.; Hattori, R.S.; Yoshinaga, T.T.; Silva, D.M.Z.A.; Foresti, F.; Santos, M.H.; Almeida, M.C.; Artoni, R.F. Differential Expression of Dmrt1 in Astyanax scabripinnis (Teleostei, Characidade) Is Correlated with B Chromosome Occurrence. Zebrafish 2019, 16, 182–188. [Google Scholar] [CrossRef] [PubMed]
- Rajendiran, P.; Jaafar, F.; Kar, S.; Sudhakumari, C.; Senthilkumaran, B.; Parhar, I.S. Sex Determination and Differentiation in Teleost: Roles of Genetics, Environment, and Brain. Biology 2021, 10, 973. [Google Scholar] [CrossRef] [PubMed]
- Herpin, A.; Schartl, M. Plasticity of Gene-Regulatory Networks Controlling Sex Determination: Of Masters, Slaves, Usual Suspects, Newcomers, and Usurpators. EMBO Rep. 2015, 16, 1260–1274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbosa, P.; Schemczssen-Graeff, Z.; Marques, A.; da Silva, M.; Favero, G.M.; Sobreiro, B.P.; de Almeida, M.C.; Moreira-Filho, O.; de Silva, D.M.Z.A.; Porto-Foresti, F.; et al. Silencing of Transposable Elements Mediated by 5-MC and Compensation of the Heterochromatin Content by Presence of B Chromosomes in Astyanax Scabripinnis. Cells 2021, 10, 1162. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, P.; de Oliveira, L.A.; Pucci, M.B.; Santos, M.H.; Moreira-Filho, O.; Vicari, M.R.; Nogaroto, V.; de Almeida, M.C.; Artoni, R.F. Identification and Chromosome Mapping of Repetitive Elements in the Astyanax scabripinnis (Teleostei: Characidae) Species Complex. Genetica 2015, 143, 55–62. [Google Scholar] [CrossRef]
- Cornelio, D.; Castro, J.P.; Santos, M.H.; Vicari, M.R.; de Almeida, M.C.; Moreira-Filho, O.; Camacho, J.P.M.; Artoni, R.F. Hermaphroditism Can Compensate for the Sex Ratio in the Astyanax scabripinnis Species Complex (Teleostei: Characidae): Expanding the B Chromosome Study Model. Rev. Fish Biol. Fish. 2017, 27, 681–689. [Google Scholar] [CrossRef]
- Castro, J.P.; Moura, M.O.; Moreira-Filho, O.; Shibatta, O.A.; Santos, M.H.; Nogaroto, V.; Vicari, M.R.; de Almeida, M.C.; Artoni, R.F. Evidence of Incipient Speciation in Astyanax scabripinnis Species Complex (Teleostei: Characidae). Neotrop. Ichthyol. 2014, 12, 429–438. [Google Scholar] [CrossRef]
- Pauls, E.; Bertollo, L.A.C. Evidence for a System of Supernumerary Chromosomes in Prochilodus scrofa Steindachner, 1881 (Pisces, Prochilodontidae). Caryologia 1983, 36, 307–314. [Google Scholar] [CrossRef]
- Dias, A.L.; Foresti, F.; Oliveira, C. Synapsis in Supernumerary Chromosomes of Prochilodus lineatus (Teleostei: Prochilodontidae). Caryologia 1998, 51, 105–113. [Google Scholar] [CrossRef]
- Basheva, E.A.; Torgasheva, A.A.; Sakaeva, G.R.; Bidau, C.; Borodin, P.M. A- and B-Chromosome Pairing and Recombination in Male Meiosis of the Silver Fox (Vulpes vulpes L., 1758, Carnivora, Canidae). Chromosome Res. 2010, 18, 689–696. [Google Scholar] [CrossRef]
- Serrano, É.A.; Araya-Jaime, C.; Suárez-Villota, E.Y.; Oliveira, C.; Foresti, F. Meiotic Behavior and H3K4m Distribution in B Chromosomes of Characidium gomesi (Characiformes, Crenuchidae). Comp. Cytogenet. 2016, 10, 255–268. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.; Voloshin, O.; Potapova, S.; Camerini-Otero, R.D. Meiotic Knockdown and Complementation Reveals Essential Role of RAD51 in Mouse Spermatogenesis. Cell Rep. 2017, 18, 1383–1394. [Google Scholar] [CrossRef] [PubMed]
- Kneitz, B.; Cohen, P.E.; Avdievich, E.; Zhu, L.; Kane, M.F.; Hou, H.; Kolodner, R.D.; Kucherlapati, R.; Pollard, J.W.; Edelmann, W. MutS Homolog 4 Localization to Meiotic Chromosomes Is Required for Chromosome Pairing during Meiosis in Male and Female Mice. Genes Dev. 2000, 14, 1085–1097. [Google Scholar] [CrossRef]
- Foe, V.E. Does the Pachytene Checkpoint, a Feature of Meiosis, Filter Out Mistakes in Double-Strand DNA Break Repair and as a Side-Effect Strongly Promote Adaptive Speciation? Integr. Org. Biol. 2022, 4, obac008. [Google Scholar] [CrossRef]
- Klassmann, J.P.; dos Santos, I.C.M. Karyotype Diversity in Astyanax scabripinnis (Pisces, Characiformes) from the São Francisco River Basin, Brazil. Cytologia 2017, 82, 199–203. [Google Scholar] [CrossRef]
- Fernandes, C.A.; Martins-Santos, I.C. Sympatric Occurrence of Three Cytotypes and Four Morphological Types of B Chromosomes of Asyanax scabripinnis (Pisces, Characiformes) in the River Ivaí Basin, State of Paraná, Brazil. Genetica 2005, 124, 301–306. [Google Scholar] [CrossRef]
- Limeira, D.M.; Santos, M.H.; Mateus, R.P.; de Almeida, M.C.; Artoni, R.F. Genetic Variability in a Population of Astyanax scabripinnis: Recent Bottleneck and the Possible Influence of Individuals with B Chromosomes. Acta Sci. Biol. Sci. 2019, 41, e47323. [Google Scholar] [CrossRef]
- Gray, S.; Cohen, P.E. Control of Meiotic Crossovers: From Double-Strand Break Formation to Designation. Annu. Rev. Genet. 2016, 50, 175–210. [Google Scholar] [CrossRef]
- Manhart, C.M.; Alani, E. Roles for Mismatch Repair Family Proteins in Promoting Meiotic Crossing Over. DNA Repair 2016, 38, 84–93. [Google Scholar] [CrossRef]
- Li, X.; Schimenti, J.C. Mouse Pachytene Checkpoint 2 (Trip13) Is Required for Completing Meiotic Recombination but Not Synapsis. PLoS Genet. 2007, 3, e130. [Google Scholar] [CrossRef]
- Queller, D.C.; Strassmann, J.E. Evolutionary Conflict. Annu. Rev. Ecol. Evol. Syst. 2018, 49, 73–93. [Google Scholar] [CrossRef]
- Dufresnes, C.; Crochet, P.-A. Sex Chromosomes as Supergenes of Speciation: Why Amphibians Defy the Rules? Philos. Trans. R. Soc. B Biol. Sci. 2022, 377, 20210202. [Google Scholar] [CrossRef] [PubMed]
- Rhie, A.; McCarthy, S.A.; Fedrigo, O.; Damas, J.; Formenti, G.; Koren, S.; Uliano-Silva, M.; Chow, W.; Fungtammasan, A.; Kim, J.; et al. Towards Complete and Error-Free Genome Assemblies of All Vertebrate Species. Nature 2021, 592, 737–746. [Google Scholar] [CrossRef]
- Aga, M.; Yamano, N.; Kumamoto, T.; Frank, J.; Onitsuka, M.; Omasa, T. Construction of a Gene Knockout CHO Cell Line Using a Simple Gene Targeting Method. BMC Proc. 2015, 9, P2. [Google Scholar] [CrossRef]
- Fu, X.; Li, N.; Lai, Y.; Luo, X.; Wang, Y.; Shi, C.; Huang, Z.; Wu, S.; Su, J. A Novel Fish Cell Line Derived from the Brain of Chinese Perch Siniperca Chuatsi: Development and Characterization. J. Fish Biol. 2015, 86, 32–45. [Google Scholar] [CrossRef] [PubMed]
- Dehler, C.E.; Boudinot, P.; Martin, S.A.M.; Collet, B. Development of an Efficient Genome Editing Method by CRISPR/Cas9 in a Fish Cell Line. Mar. Biotechnol. 2016, 18, 449–452. [Google Scholar] [CrossRef]
- Periyasamy, T.; Xiao Joe, J.T.; Lu, M.-W. Cloning and Expression of Malabar Grouper (Epinephelus malabaricus) ADAR1 Gene in Response to Immune Stimulants and Nervous Necrosis Virus. Fish Shellfish Immunol. 2017, 71, 116–126. [Google Scholar] [CrossRef]
- Collet, B.; Collins, C.; Lester, K. Engineered Cell Lines for Fish Health Research. Dev. Comp. Immunol. 2018, 80, 34–40. [Google Scholar] [CrossRef]
- Trifonov, V.A.; Dementyeva, P.V.; Larkin, D.M.; O’Brien, P.C.; Perelman, P.L.; Yang, F.; Ferguson-Smith, M.A.; Graphodatsky, A.S. Transcription of a Protein-Coding Gene on B Chromosomes of the Siberian Roe Deer (Capreolus pygargus). BMC Biol. 2013, 11, 90. [Google Scholar] [CrossRef]
- Alves, A.L.; Martins-Santos, I.C. Cytogenetics Studies in Two Populations of Astyanax scabripinnis with 2n=48 Chromosomes (Teleostei, Characidae). Cytologia 2002, 67, 117–122. [Google Scholar] [CrossRef]
- Araujo, A.S.; Morelli, S. Estudo Cariotípico Da População de Astyanax scabripinnis (Pisces, Characidae) Da Nascente Do Córrego Jataí, Uberlândia-MG. Genet Mol. Biol. 2000, 23, 60. [Google Scholar]
- Mizoguchi, S.M.H.N.; Martins-Santos, I.C. Macro- and Microchromosomes B in Females of Astyanax scabripinnis (Pisces, Characidae). Hereditas 1997, 127, 249–253. [Google Scholar] [CrossRef]
- Souza, I.L.; Moreira-Filho, O. Cytogenetic Diversity in the Astyanax scabripinnis Species Complex (Pisces, Characidae). Cytologia 1995, 60, 1–11. [Google Scholar] [CrossRef]
- Fauaz, G. Natural Triploidy and B Chromosomes in the Neotropical Fish Genus Astyanax (Characidae). Braz. J. Genet. 1994, 7, 639–645. [Google Scholar]
- Goes, C.A.G.; Silva, D.M.Z.A.; Utsunomia, R.; Yasui, G.S.; Artoni, R.F.; Foresti, F.; Porto-Foresti, F. Establishment of Rapid and Non-Invasive Protocols to Identify B-Carrying Individuals of Psalidodon paranae. Genet. Mol. Biol. 2021, 44, e20200003. [Google Scholar] [CrossRef]
- Abelini, E.; Martins-Santos, I.C.; Fernandes, C.A. Cytogenetic Analysis in Three Species from Genus Astyanax (Pisces; Characiformes) with a New Occurrence of B Chromosome in Astyanax paranae. Caryologia 2014, 67, 160–171. [Google Scholar] [CrossRef]
- Maistro, E.L.; Lúcia Dias, A.; Foresti, F.; Oliveira, C.; Filho, O.M. Natural Triploidy in Astyanax scabripinnis (Pisces, Characidae) and Simultaneous Occurrence of Macro B-Chromosomes. Caryologia 1994, 47, 233–239. [Google Scholar] [CrossRef]
- Maistro, E.L.; Foresti, F.; Oliveira, C.; de Almeida-Toledo, L.F. Occurrence of Macro B Chromosomes in Astyanax scabripinnis paranae (Pisces, Characiformes, Characidae). Genetica 1992, 87, 101–106. [Google Scholar] [CrossRef]
- Hashimoto, D.T.; Ferguson-Smith, M.A.; Rens, W.; Foresti, F.; Porto-Foresti, F. Chromosome Mapping of H1 Histone and 5S RRNA Gene Clusters in Three Species of Astyanax (Teleostei, Characiformes). Cytogenet. Genome Res. 2011, 134, 64–71. [Google Scholar] [CrossRef]
- Daniel, S.N.; Penitente, M.; Hashimoto, D.T.; Silva, D.M.Z.A.; Foresti, F.; Porto-Foresti, F. New Insights into Karyotypic Relationships Among Populations of Astyanax bockmanni (Teleostei, Characiformes) of Different Watersheds. Zebrafish 2015, 12, 181–188. [Google Scholar] [CrossRef]
- Ahmad, S.F.; Jehangir, M.; Cardoso, A.L.; Wolf, I.R.; Margarido, V.P.; Cabral-de-Mello, D.C.; O’Neill, R.; Valente, G.T.; Martins, C. B Chromosomes of Multiple Species Have Intense Evolutionary Dynamics and Accumulated Genes Related to Important Biological Processes. BMC Genom. 2020, 21, 656. [Google Scholar] [CrossRef] [PubMed]
- Torres-Mariano, A.R.; Morelli, S. B Chromosomes in a Population of Astyanax eigenmanniorum (Characiformes, Characidae) from the Araguari River Basin (Uberlândia, MG, Brazil). Genet. Mol. Biol. 2008, 31, 246–249. [Google Scholar] [CrossRef]
- Moreira-Filho, O.; Fenocchio, A.S.; Pastori, M.C.; Bertollo, L.A.C. Occurrence of a Metacentric Macrochromosome B in Different Species of the Genus Astyanax (Pisces, Characidae, Tetragonopterinae). Cytologia 2001, 66, 59–64. [Google Scholar] [CrossRef]
- Fazoli, L.C.; Silva, V. de A.B. da; Portela-Castro, A.L. de B.; Júnior, H.F.J. Chromosome Characterization of Astyanax Sp B (Characidae, Tetragonopterinae), an Endemic Species of the Iguaçu River, Paraná, Brazil. Cytologia 2003, 68, 389–394. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, D.M.Z.A.; Castro, J.P.; Goes, C.A.G.; Utsunomia, R.; Vidal, M.R.; Nascimento, C.N.; Lasmar, L.F.; Paim, F.G.; Soares, L.B.; Oliveira, C.; et al. B Chromosomes in Psalidodon scabripinnis (Characiformes, Characidae) Species Complex. Animals 2022, 12, 2174. https://doi.org/10.3390/ani12172174
Silva DMZA, Castro JP, Goes CAG, Utsunomia R, Vidal MR, Nascimento CN, Lasmar LF, Paim FG, Soares LB, Oliveira C, et al. B Chromosomes in Psalidodon scabripinnis (Characiformes, Characidae) Species Complex. Animals. 2022; 12(17):2174. https://doi.org/10.3390/ani12172174
Chicago/Turabian StyleSilva, Duílio M. Z. A., Jonathan P. Castro, Caio A. G. Goes, Ricardo Utsunomia, Mateus R. Vidal, Cristiano N. Nascimento, Lucas F. Lasmar, Fabilene G. Paim, Letícia B. Soares, Claudio Oliveira, and et al. 2022. "B Chromosomes in Psalidodon scabripinnis (Characiformes, Characidae) Species Complex" Animals 12, no. 17: 2174. https://doi.org/10.3390/ani12172174