
Genomic Organization of Microsatellites and LINE-1-like Retrotransposons: Evolutionary Implications for Ctenomys minutus (Rodentia: Ctenomyidae) Cytotypes

 , ,
, ,  ,
,  , ,
, ,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Chromosomal Preparations
2.3. Probe Preparation and Fluorescence In Situ Hybridization (FISH)
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- López-Flores, I.; Garrido-Ramos, M.A. The Repetitive DNA Content of Eukaryotic Genomes. Genome Dyn. 2012, 7, 1–28. [Google Scholar] [CrossRef]
- Liehr, T. Repetitive Elements in Humans. Int. J. Mol. Sci. 2021, 22, 2072. [Google Scholar] [CrossRef]
- Gibbs, R.A.; Weinstock, G.M.; Metzker, M.L.; Muzny, D.M.; Sodergren, E.J.; Scherer, S.; Scott, G.; Steffen, D.; Worley, K.C.; Burch, P.E.; et al. Genome Sequence of the Brown Norway Rat Yields Insights into Mammalian Evolution. Nature 2004, 428, 493–520. [Google Scholar] [CrossRef]
- de Koning, A.P.J.; Gu, W.; Castoe, T.A.; Batzer, M.A.; Pollock, D.D. Repetitive Elements May Comprise Over Two-Thirds of the Human Genome. PLoS Genet. 2011, 7, e1002384. [Google Scholar] [CrossRef]
- Biet, E.; Sun, J.-S.; Dutreix, M. Conserved Sequence Preference in DNA Binding among Recombination Proteins: An Effect of SsDNA Secondary Structure. Nucleic Acids Res. 1999, 27, 596–600. [Google Scholar] [CrossRef]
- Liu, Z.; Li, P.; Kocabas, A.; Karsi, A.; Ju, Z. Microsatellite-Containing Genes from the Channel Catfish Brain: Evidence of Trinucleotide Repeat Expansion in the Coding Region of Nucleotide Excision Repair Gene RAD23B. Biochem. Biophys. Res. Commun. 2001, 289, 317–324. [Google Scholar] [CrossRef]
- Kidwell, M.G. Transposable Elements and the Evolution of Genome Size in Eukaryotes. Genetica 2002, 115, 49–63. [Google Scholar] [CrossRef]
- Li, Y.; Korol, A.B.; Fahima, T.; Beiles, A.; Nevo, E. Microsatellites: Genomic Distribution, Putative Functions and Mutational Mechanisms: A Review. Mol. Ecol. 2002, 11, 2453–2465. [Google Scholar] [CrossRef]
- De Oliveira, T.D.; Kretschmer, R.; Bertocchi, N.A.; Degrandi, T.M.; De Oliveira, E.H.C.; De Cioffi, M.B.; Garnero, A.D.V.; Gunski, R.J. Genomic Organization of Repetitive DNA in Woodpeckers (Aves, Piciformes): Implications for Karyotype and ZW Sex Chromosome Differentiation. PLoS ONE 2017, 12, e0169987. [Google Scholar] [CrossRef]
- Martins, C. Chromosomes and Repetitive DNAs: A Contribution to the Knowledge of the Fish Genome. In Fish Cytogenetics; Pisano, E., Ozouf-Costaz, C., Foresti, F., Kapoor, B., Eds.; Science Publisher: New York, NY, USA, 2007; pp. 421–432. [Google Scholar]
- Komissarov, A.S.; Gavrilova, E.V.; Demin, S.J.; Ishov, A.M.; Podgornaya, O.I. Tandemly Repeated DNA Families in the Mouse Genome. BMC Genom. 2011, 12, 531. [Google Scholar] [CrossRef]
- Pokorná, M.; Kratochvíl, L.; Kejnovský, E. Microsatellite Distribution on Sex Chromosomes at Different Stages of Heteromorphism and Heterochromatinization in Two Lizard Species (Squamata: Eublepharidae: Coleonyx elegans and Lacertidae: Eremias velox). BMC Genet. 2011, 12, 90. [Google Scholar] [CrossRef] [PubMed]
- Cioffi, M.d.B.; Bertollo, L.A.C. Chromosomal Distribution and Evolution of Repetitive DNAs in Fish. Repetitive DNA 2012, 7, 197–221. [Google Scholar] [CrossRef]
- Paço, A.; Adega, F.; Chaves, R. LINE-1 Retrotransposons: From ‘Parasite’ Sequences to Functional Elements. J. Appl. Genet. 2014, 56, 133–145. [Google Scholar] [CrossRef] [PubMed]
- Bertocchi, N.A.; de Oliveira, T.D.; del Valle Garnero, A.; Coan, R.L.B.; Gunski, R.J.; Martins, C.; Torres, F.P. Distribution of CR1-like Transposable Element in Woodpeckers (Aves Piciformes): Z Sex Chromosomes Can Act as a Refuge for Transposable Elements. Chromosom. Res. 2018, 26, 333–343. [Google Scholar] [CrossRef] [PubMed]
- Wicker, T.; Sabot, F.; Hua-Van, A.; Bennetzen, J.L.; Capy, P.; Chalhoub, B.; Flavell, A.; Leroy, P.; Morgante, M.; Panaud, O.; et al. A Unified Classification System for Eukaryotic Transposable Elements. Nat. Rev. Genet. 2007, 8, 973–982. [Google Scholar] [CrossRef]
- Burton, F.H.; Loeb, D.D.; Voliva, C.F.; Martin, S.L.; Edgell, M.H.; Hutchison, C.A. Conservation throughout Mammalia and Extensive Protein-Encoding Capacity of the Highly Repeated DNA Long Interspersed Sequence One. J. Mol. Biol. 1986, 187, 291–304. [Google Scholar] [CrossRef]
- Boissinot, S.; Chevret, P.; Furano, A.V. L1 (LINE-1) Retrotransposon Evolution and Amplification in Recent Human History. Mol. Biol. Evol. 2000, 17, 915–928. [Google Scholar] [CrossRef]
- Furano, A.; Duvernell, D.; Boissinot, S. L1 (LINE-1) Retrotransposon Diversity Differs Dramatically between Mammals and Fish. Trends Genet. 2004, 20, 9–14. [Google Scholar] [CrossRef]
- Bratthauer, G.L.; Cardiff, R.D.; Fanning, T.G. Expression of LINE-1 Retrotransposons in Human Breast Cancer. Cancer 1994, 73, 2333–2336. [Google Scholar] [CrossRef]
- Esnault, C.; Maestre, J.; Heidmann, T. Human LINE Retrotransposons Generate Processed Pseudogenes. Nat. Genet. 2000, 24, 363–367. [Google Scholar] [CrossRef]
- Ovchinnikov, I.; Troxel, A.B.; Swergold, G.D. Genomic Characterization of Recent Human LINE-1 Insertions: Evidence Supporting Random Insertion. Genome Res. 2001, 11, 2050–2058. [Google Scholar] [CrossRef] [PubMed]
- Acosta, M.J.; Marchal, J.A.; Fernández-Espartero, C.H.; Bullejos, M.; Sánchez, A. Retroelements (LINEs and SINEs) in Vole Genomes: Differential Distribution in the Constitutive Heterochromatin. Chromosom. Res. 2008, 16, 949–959. [Google Scholar] [CrossRef]
- Dobigny, G.; Ozouf-Costaz, C.; Waters, P.D.; Bonillo, C.; Coutanceau, J.P.; Volobouev, V. LINE-1 Amplification Accompanies Explosive Genome Repatterning in Rodents. Chromosom. Res. 2005, 12, 787–793. [Google Scholar] [CrossRef]
- Appels, R.; Morris, R.; Gill, B.S.; May, C.E. Chromosome Morphology and Number. In Chromosome Biology; Springer: Boston, MA, USA, 1998; pp. 67–84. [Google Scholar]
- Teta, P.; D’Elía, G. Uncovering the Species Diversity of Subterranean Rodents at the End of the World: Three New Species of Patagonian Tuco-Tucos (Rodentia, Hystricomorpha, Ctenomys). PeerJ 2020, 8, e9259. [Google Scholar] [CrossRef] [PubMed]
- Reig, O.A.; Bush, C.; Ortells, M.O.; Contreras, J.R. An Overview of Evolution, Systematic, Population Biology, Cytogenetics, Molecular Biology and Speciation in Ctenomys. In Evolution of Subterranean Mammals at the Organismal and Molecular Level; Nevo, E., Reig, O.A., Eds.; Alan R. Liss: New York, NY, USA, 1990; pp. 71–96. [Google Scholar]
- Freitas, T.R.O. Family Ctenomyidae (Tuco-Tucos). In Handbook of the Mammals of the World. Lagomorphs and Rodents I; Wilson, D., Lacer, T., Mittermeier, R., Eds.; Lynx Edicions Publications: Barcelona, Spain, 2016; Volume 6, pp. 498–534. ISBN 978-84-941892-3-4. [Google Scholar]
- Freitas, T.R.O. De Chromosome Polymorphism in Ctenomys minutus (Rodentia-Octodontidae). Braz. J. Genet. 1997, 20, 1–7. [Google Scholar] [CrossRef]
- Massarini, A.; Mizrahi, D.; Tiranti, S.; Toloza, A.; Luna, F.; Schleich, C.E. Extensive Chromosomal Variation in Ctenomys talarum talarum from the Atlantic Coast of Buenos Aires Province, Argentina (Rodentia: Octodontidae). Mastozool. Neotrop. 2002, 9, 199–207. [Google Scholar]
- Freygang, C.C.; Marinho, J.R.; de Freitas, T.R.O. New Karyotypes and Some Considerations about the Chromosomal Diversification of Ctenomys minutus (Rodentia: Ctenomyidae) on the Coastal Plain of the Brazilian State of Rio Grande Do Sul. Genetica 2004, 121, 125–132. [Google Scholar] [CrossRef]
- Freitas, T.R.O. Ctenomys lami: The Highest Chromosome Variability in Ctenomys (Rodentia, Ctenomyidae) Due to a Centric Fusion/Fission and Pericentric Inversion System. Acta Theriol. 2007, 52, 171–180. [Google Scholar] [CrossRef]
- Fernandes, F.; Fernández-Stolz, G.; Lopes, C.; Freitas, T. The Conservation Status of the Tuco-Tucos, Genus Ctenomys (Rodentia: Ctenomyidae), in Southern Brazil. Braz. J. Biol. 2007, 67, 839–847. [Google Scholar] [CrossRef]
- Freitas, T.R.O. Geographic Distribution and Conservation of Four Species of the Genus Ctenomys in Southern Brazil. Stud. Neotrop. Fauna Environ. 1995, 30, 53–59. [Google Scholar] [CrossRef]
- Gava, A.; de Freitas, T.R.O. Characterization of a Hybrid Zone Between Chromosomally Divergent Populations of Ctenomys minutus (Rodentia: Ctenomyidae). J. Mammal. 2002, 83, 843–851. [Google Scholar] [CrossRef]
- Gava, A.; Freitas, T.R.O. De Microsatellite Analysis of a Hybrid Zone Between Chromosomally Divergent Populations of Ctenomys minutus from Southern Brazil (Rodentia: Ctenomyidae). J. Mammal. 2004, 85, 1201–1206. [Google Scholar] [CrossRef]
- Lopes, C.M.; Ximenes, S.S.F.; Gava, A.; de Freitas, T.R.O. The Role of Chromosomal Rearrangements and Geographical Barriers in the Divergence of Lineages in a South American Subterranean Rodent (Rodentia: Ctenomyidae: Ctenomys minutus). Heredity 2013, 111, 293–305. [Google Scholar] [CrossRef] [PubMed]
- Freitas, T.R.O. De Cytogenetics Status of Four Ctenomys Species in the South of Brazil. Genetica 2006, 126, 227–235. [Google Scholar] [CrossRef]
- Gava, A.; Freitas, T.R.O. Inter and Intra-Specific Hybridization in Tuco-Tucos (Ctenomys) from Brazilian Coastal Plains (Rodentia: Ctenomyidae). Genetica 2003, 119, 11–17. [Google Scholar] [CrossRef]
- Fornel, R.; Cordeiro-Estrela, P.; de Freitas, T.R.O. Skull Shape and Size Variation within and between mendocinus and torquatus Groups in the Genus Ctenomys (Rodentia: Ctenomyidae) in Chromosomal Polymorphism Context. Genet. Mol. Biol. 2018, 41, 263–272. [Google Scholar] [CrossRef]
- Ditcharoen, S.; Bertollo, L.A.C.; Ráb, P.; Hnátková, E.; Molina, W.F.; Liehr, T.; Tanomtong, A.; Triantaphyllidis, C.; Ozouf-Costaz, C.; Tongnunui, S.; et al. Genomic Organization of Repetitive DNA Elements and Extensive Karyotype Diversity of Silurid Catfishes (Teleostei: Siluriformes): A Comparative Cytogenetic Approach. Int. J. Mol. Sci. 2019, 20, 3545. [Google Scholar] [CrossRef]
- Sember, A.; De Oliveira, E.A.; Ráb, P.; Bertollo, L.A.C.; De Freitas, N.L.; Viana, P.F.; Yano, C.F.; Hatanaka, T.; Marinho, M.M.F.; De Moraes, R.L.R.; et al. Centric Fusions behind the Karyotype Evolution of Neotropical Nannostomus pencilfishes (Characiforme, Lebiasinidae): First Insights from a Molecular Cytogenetic Perspective. Genes 2020, 11, 91. [Google Scholar] [CrossRef]
- Rossi, M.S.; Pesce, C.G.; Reig, O.A.; Kornblihtt, A.R.; Zorzópulos, J. Retroviral-like Features in the Monomer of the Major Satellite DNA from the South American Rodents of the Genus Ctenomys. Mitochondrial DNA 1993, 3, 379–381. [Google Scholar] [CrossRef]
- Novello, A.; Cortinas, M.N.; Suárez, M.; Musto, H. Cytogenetic and Molecular Analysis of the Satellite DNA of the Genus Ctenomys (Rodentia Octodontidae) from Uruguay. Chromosom. Res. 1996, 4, 335–339. [Google Scholar] [CrossRef]
- Sikes, R.S. The animal Care and use committee of the american Society of mammalogists 2016 Guidelines of the American Society of Mammalogists for the Use of Wild Mammals in Research and Education. J. Mammal. 2016, 97, 663–688. [Google Scholar] [CrossRef] [PubMed]
- Verma, R.; Babu, A. Human Chromosomes: Principles & Techniques. Mol. Reprod. Dev. 1996, 43, 134. [Google Scholar] [CrossRef]
- Kubat, Z.; Hobza, R.; Vyskot, B.; Kejnovsky, E. Microsatellite Accumulation on the Y Chromosome in Silene latifolia. Genome 2008, 356, 350–356. [Google Scholar] [CrossRef] [PubMed]
- Doyle, J.J.; Doyle, J.L. A Rapid DNA Isolation Procedure for Small Quantities of Fresh Leaf Tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Casavant, N.C.; Scott, L.; Cantrell, M.A.; Wiggins, L.E.; Baker, R.J.; Wichman, H.A. The End of the LINE? Lack of Recent L1 Activity in a Group of South American Rodents. Genetics 2000, 154, 1809–1817. [Google Scholar] [CrossRef]
- Richard, G.-F.; Kerrest, A.; Dujon, B. Comparative Genomics and Molecular Dynamics of DNA Repeats in Eukaryotes. Microbiol. Mol. Biol. Rev. 2008, 72, 686–727. [Google Scholar] [CrossRef]
- Liehr, T.; Kosayakova, N.; Schröder, J.; Ziegler, M.; Kreskowski, K.; Pohle, B.; Bhatt, S.; Theuss, L.; Wilhelm, K.; Weise, A.; et al. Evidence for Correlation of Fragile Sites and Chromosomal Breakpoints in Carriers of Constitutional Balanced Chromosomal Rearrangements. Balk. J. Med. Genet. 2011, 14, 13–16. [Google Scholar] [CrossRef]
- Ruiz-Herrera, A.; Castresana, J.; Robinson, T.J. Is Mammalian Chromosomal Evolution Driven by Regions of Genome Fragility? Genome Biol. 2006, 7, R115. [Google Scholar] [CrossRef]
- Ferguson-Smith, M.A.; Trifonov, V. Mammalian Karyotype Evolution. Nat. Rev. Genet. 2007, 8, 950–962. [Google Scholar] [CrossRef]
- Fan, Y.; Newman, T.; Linardopoulou, E.; Trask, B.J. Gene Content and Function of the Ancestral Chromosome Fusion Site in Human Chromosome 2q13-2q14.1 and Paralogous Regions. Genome Res. 2002, 12, 1663–1672. [Google Scholar] [CrossRef]
- Kehrer-Sawatzki, H.; Schreiner, B.; Tänzer, S.; Platzer, M.; Müller, S.; Hameister, H. Molecular Characterization of the Pericentric Inversion That Causes Differences between Chimpanzee Chromosome 19 and Human Chromosome 17. Am. J. Hum. Genet. 2002, 71, 375–388. [Google Scholar] [CrossRef] [PubMed]
- Locke, D.P.; Archidiacono, N.; Misceo, D.; Cardone, M.F.; Deschamps, S.; Roe, B.; Rocchi, M.; Eichler, E.E. Refinement of a Chimpanzee Pericentric Inversion Breakpoint to a Segmental Duplication Cluster. Genome Biol. 2003, 4, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Kehrer-Sawatzki, H.; Sandig, C.A.; Goidts, V.; Hameister, H. Breakpoint Analysis of the Pericentric Inversion between Chimpanzee Chromosome 10 and the Homologous Chromosome 12 in Humans. Cytogenet. Genome Res. 2005, 108, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Leffak, M. Break-Induced Replication Links Microsatellite Expansion to Complex Genome Rearrangements. BioEssays 2017, 39, 1700025. [Google Scholar] [CrossRef] [PubMed]
- Balzano, E.; Pelliccia, F.; Giunta, S. Genome (in)Stability at Tandem Repeats. Semin. Cell Dev. Biol. 2021, 113, 97–112. [Google Scholar] [CrossRef]
- Gadgil, R.Y.; Romer, E.J.; Goodman, C.C.; Dean Rider, S.; Damewood, F.J.; Barthelemy, J.R.; Shin-Ya, K.; Hanenberg, H.; Leffak, M. Replication Stress at Microsatellites Causes DNA Double-Strand Breaks and Break-Induced Replication. J. Biol. Chem. 2020, 295, 15378–15397. [Google Scholar] [CrossRef] [PubMed]
- Feng, W.; Di Rienzi, S.C.; Raghuraman, M.K.; Brewer, B.J. Replication Stress-Induced Chromosome Breakage Is Correlated with Replication Fork Progression and Is Preceded by Single-Stranded DNA Formation. G3 Genes Genomes Genet. 2011, 1, 327–335. [Google Scholar] [CrossRef]
- Glover, T.W. Common Fragile Sites. Cancer Lett. 2006, 232, 4–12. [Google Scholar] [CrossRef]
- Schwartz, M.; Zlotorynski, E.; Kerem, B. The Molecular Basis of Common and Rare Fragile Sites. Cancer Lett. 2006, 232, 13–26. [Google Scholar] [CrossRef]
- Yakovchuk, P.; Protozanova, E.; Frank-kamenetskii, M.D. Base-Stacking and Base-Pairing Contributions into Thermal Stability of the DNA Double Helix. Nucleic Acids Res. 2006, 34, 564–574. [Google Scholar] [CrossRef]
- Volff, J.; Körting, C.; Meyer, A.; Schartl, M. Evolution and Discontinuous Distribution of Rex3 Retrotransposons in Fish. Mol. Biol. Evol. 2001, 18, 427–431. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, D.C.; Porto-Foresti, F.; Oliveira, C.; Foresti, F. Transposable Elements as a Potential Source for Understanding the Fish Genome. Mob. Genet. Elements 2011, 1, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Araújo, N.P.; Sena, R.S.; Bonvicino, C.R.; Kuhn, G.C.S.; Svartman, M. SINE-B1 Distribution and Chromosome Rearrangements in the South American Proechimys gr. goeldii (Echimyidae, Rodentia). Cytogenet. Genome Res. 2021, 161, 6–13. [Google Scholar] [CrossRef] [PubMed]
- Vieira-da-Silva, A.; Adega, F.; Guedes-Pinto, H.; Chaves, R. LINE-1 Distribution in Six Rodent Genomes Follow a Species-Specific Pattern. J. Genet. 2016, 95, 21–33. [Google Scholar] [CrossRef]
- Slamovits, C.H.; Cook, J.A.; Lessa, E.P.; Susana Rossi, M. Recurrent Amplifications and Deletions of Satellite DNA Accompanied Chromosomal Diversification in South American Tuco-Tucos (Genus Ctenomys, Rodentia: Octodontidae): A Phylogenetic Approach. Mol. Biol. Evol. 2001, 18, 1708–1719. [Google Scholar] [CrossRef]
- Erickson, I.K.; Cantrell, M.A.; Scott, L.; Wichman, H.A. Retrofitting the Genome: L1 Extinction Follows Endogenous Retroviral Expansion in a Group of Muroid Rodents. J. Virol. 2011, 85, 12315–12323. [Google Scholar] [CrossRef]
- Feschotte, C.; Pritham, E.J. Mobile DNA: Genomes under the Influence. Genome Biol. 2006, 7, 12315–12323. [Google Scholar] [CrossRef]
- Waminal, N.E.; Pellerin, R.J.; Kang, S.H.; Kim, H.H. Chromosomal Mapping of Tandem Repeats Revealed Massive Chromosomal Rearrangements and Insights Into Senna tora Dysploidy. Front. Plant Sci. 2021, 12, 629898. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
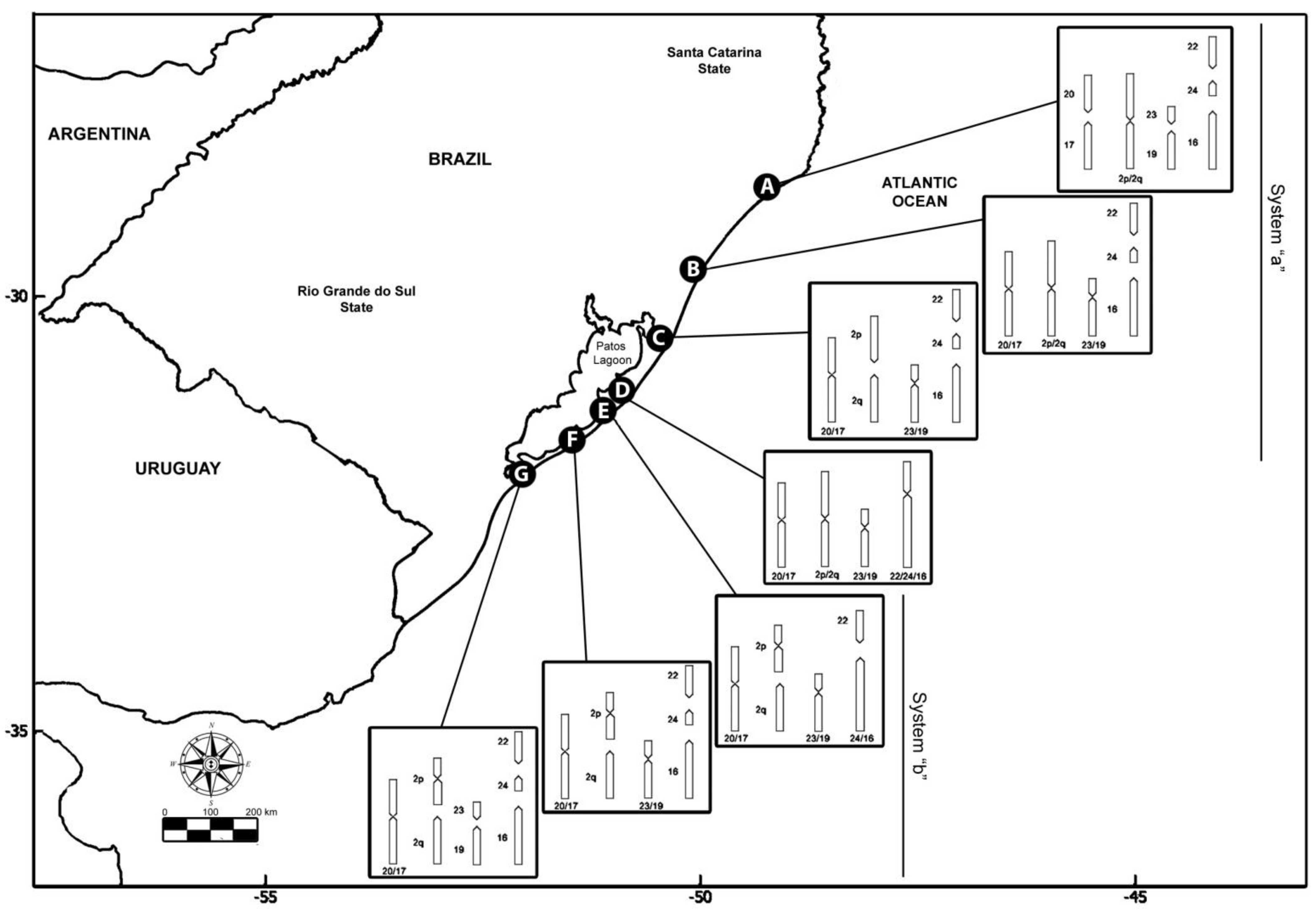
| Cytotype | 2n | Karyotype | NF | Fusion | Fission |
|---|---|---|---|---|---|
| Cytotype A | 50 a | 28 sm + 20 a; XY | 76 | Standard | |
| Cytotype B | 46 a | 32 sm + 12 a; XY | 76 | 20/17; 23/19 | - |
| Cytotype C | 48 a | 30 sm + 16 a; XY | 76 | 20/17; 23/19 | 2p; 2q |
| Cytotype D | 42 | 36 sm + 6 a; XY | 78 | 20/17; 23/19; 22/24/16 | - |
| Cytotype E | 46 b | 32 sm + 12 a; XY | 76 | 20/17; 23/19; 24/16 | 2p *; 2q |
| Cytotype F | 48 b | 30 sm + 16 a; XY | 76 | 20/17; 23/19 | 2p *; 2q |
| Cytotype G | 50 b | 28 sm + 20 a; XY | 76 | 20/17 | 2p *; 2q |
| Cytotype | Individuals/Sex | Locality | Geographic Coordinate |
|---|---|---|---|
| A | 1 ♀ | Jaguaruna (SC), Brazil | 28°41′53.02″ S 49°01′33.86″ W |
| B | 2 ♀ | Praia do Barro (RS), Brazil | 29°42′14.86″ S 49°58′51.86″ W |
| C | 2 ♂♀ | Bacupari (RS), Brazil | 30°28′41.01″ S 50°27′13.92″ W |
| D | 2 ♀ | Mostardas (RS), Brazil | 31°06′17″ S 50°55′20″ W |
| E | 2 ♂♀ | Tavares (RS), Brazil | 31°17′58.9″ S 51°05′47.6″ W |
| F | 2 ♀ | Bojuru (RS), Brazil | 31°39′10.7″ S 51°26′14.8″ W |
| G | 2 ♀ | São José do Norte (RS), Brazil | 32°04′34.47″ S 52°02′31.47″ W |
| Cytotypes | Motif Probes | |||||||
|---|---|---|---|---|---|---|---|---|
| (A)30 | (C)30 | (CA)15 | (CAC)10 | (CAG)10 | (CGG)10 | (GA)15 | (GAG)10 | |
| Cyt A | Terminal and centromeric blocks on all chromosomes | Terminal blocks on all chromosomes and interstitial block on chr 1 and 8 | Terminal and centromeric blocks on large chromosomes | Terminal blocks on all chromosomes and centromeric blocks on large chromosomes | Terminal and centromeric blocks on most chromosomes | Terminal blocks on large chromosomes and interstitial block on chr 8 | Terminal blocks on all chromosomes | Terminal and centromeric blocks on all chromosomes |
| Cyt B | Terminal and centromeric blocks on all chromosomes | Terminal blocks on most chromosomes and interstitial block on chr 8 | Terminal and centromeric blocks on all chromosomes | Terminal blocks on some chromosomes | Terminal blocks on some chromosomes | Terminal blocks on two chromosome pairs and interstitial block on chr 8 | Terminal and interstitial blocks on all chromosomes | Terminal and interstitial blocks on some chromosomes |
| Cyt C | Terminal block on large chromosomes and centromeric block on all chromosomes | Terminal blocks on ten chromosome pairs and interstitial block on chr 1 and 8 | Large blocks on all chromosomes | Terminal blocks on some chromosomes | Terminal and interstitial blocks on some chromosomes | Terminal blocks on some chromosomes and interstitial block on chr 8 | Terminal and interstitial blocks on some chromosomes | Terminal and interstitial blocks on some chromosomes |
| Cyt D | Terminal and centromeric blocks on large chromosomes | Terminal blocks on all chromosomes and interstitial block on chr 8 | Large blocks on larges chromosomes | Terminal and interstitial blocks on some chromosomes | Terminal and interstitial blocks on some chromosomes | Terminal blocks on ten chromosome pairs and interstitial block on chr 8 | Terminal and interstitial blocks on all chromosomes | Terminal and interstitial blocks on all chromosomes |
| Cyt E | Terminal block on some chromosomes and centromeric block on some large chromosome | Terminal blocks on all chromosomes and interstitial block on chr 8 | Large blocks in the q arm on all chromosomes | Terminal and proximal blocks on all chromosomes | Terminal and interstitial blocks on some chromosomes | Terminal blocks on four chromosome pairs and interstitial block on chr 8 | Terminal and interstitial blocks on all chromosomes | Terminal and interstitial blocks on some chromosomes |
| Cyt F | Terminal block on some chromosomes and centromeric block on large chromosome | Terminal blocks on eight chromosome pairs and interstitial block on chr 1 and 8 | Terminal blocks on some chromosomes | Terminal blocks on all chromosomes | Terminal blocks on some chromosomes | Interstitial block on chr 8 | Terminal blocks on some chromosomes | Terminal blocks on some chromosomes |
| Cyt G | Terminal block on some chromosomes and centromeric block on some large chromosomes | Terminal blocks on six chromosome pairs and interstitial block on chr 8 | Terminal and centromeric blocks on all chromosomes | Terminal and proximal blocks on all chromosomes | Terminal and proximal blocks on all chromosomes | Terminal blocks on four chromosome pairs and interstitial block on chr 8 | Terminal and proximal blocks on all chromosomes | Terminal block on all chromosomes and centromeric blocks on chr 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Oliveira, T.D.; Bertocchi, N.A.; Kretschmer, R.; de Oliveira, E.H.C.; Cioffi, M.d.B.; Liehr, T.; de Freitas, T.R.O. Genomic Organization of Microsatellites and LINE-1-like Retrotransposons: Evolutionary Implications for Ctenomys minutus (Rodentia: Ctenomyidae) Cytotypes. Animals 2022, 12, 2091. https://doi.org/10.3390/ani12162091
de Oliveira TD, Bertocchi NA, Kretschmer R, de Oliveira EHC, Cioffi MdB, Liehr T, de Freitas TRO. Genomic Organization of Microsatellites and LINE-1-like Retrotransposons: Evolutionary Implications for Ctenomys minutus (Rodentia: Ctenomyidae) Cytotypes. Animals. 2022; 12(16):2091. https://doi.org/10.3390/ani12162091
Chicago/Turabian Stylede Oliveira, Thays Duarte, Natasha Avila Bertocchi, Rafael Kretschmer, Edivaldo H. C. de Oliveira, Marcelo de Bello Cioffi, Thomas Liehr, and Thales R. O. de Freitas. 2022. "Genomic Organization of Microsatellites and LINE-1-like Retrotransposons: Evolutionary Implications for Ctenomys minutus (Rodentia: Ctenomyidae) Cytotypes" Animals 12, no. 16: 2091. https://doi.org/10.3390/ani12162091