
“Emotional Proximity” and “Spatial Proximity”: Higher Relationship Quality and Nearer Distance Both Strengthen Scratch Contagion in Tibetan Macaques


Abstract
:Simple Summary
Abstract
1. Introduction
- Predictions 1 and 2
- Prediction 3
- Prediction 4
2. Materials and Methods
2.1. Study Site and Subjects
2.2. Data Collection and Behavioral Definition
2.3. Data Analysis
2.3.1. Determination of Behavioral Contagion
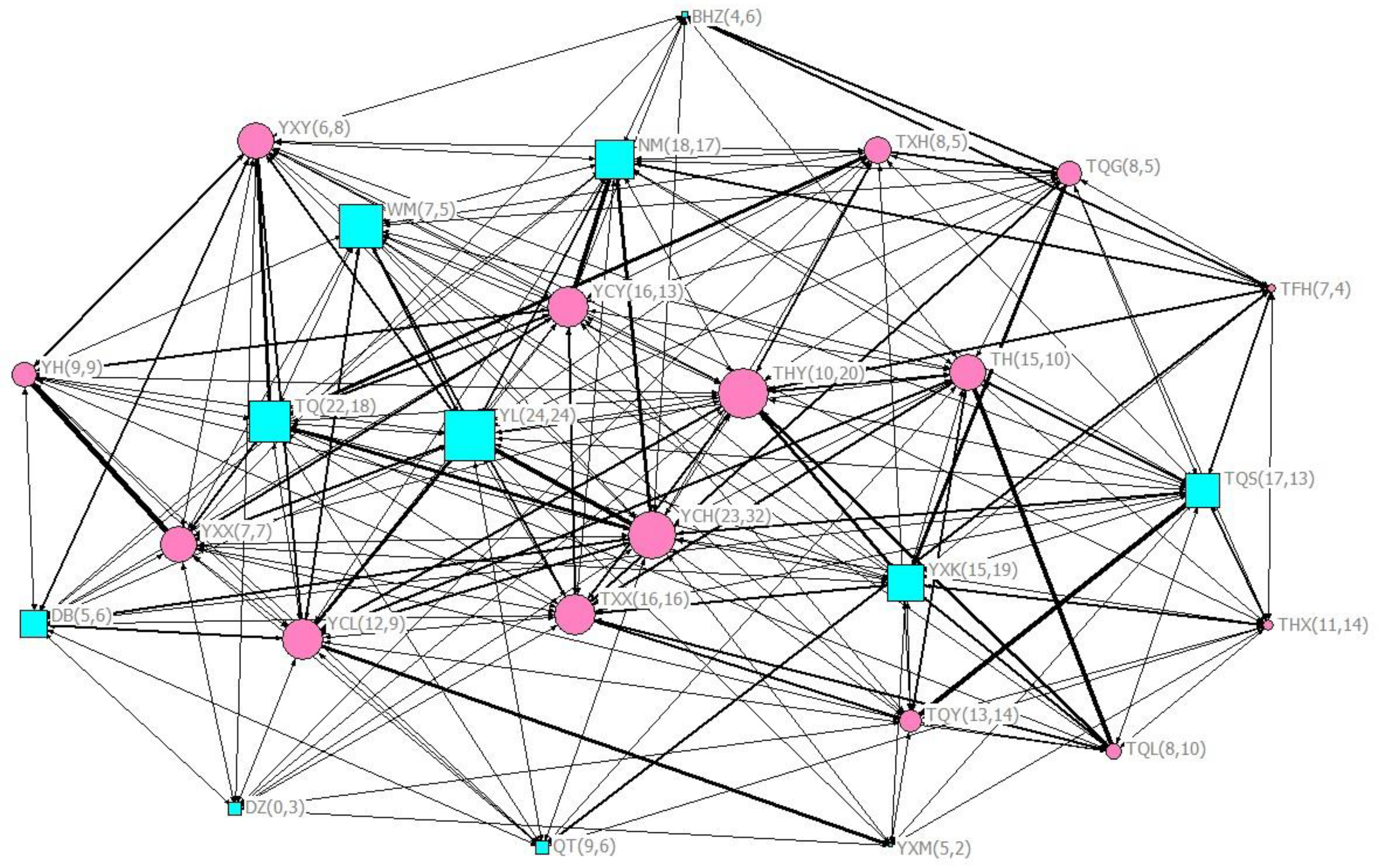
2.3.2. Relationship Quality
2.4. Statistical Analysis
3. Results
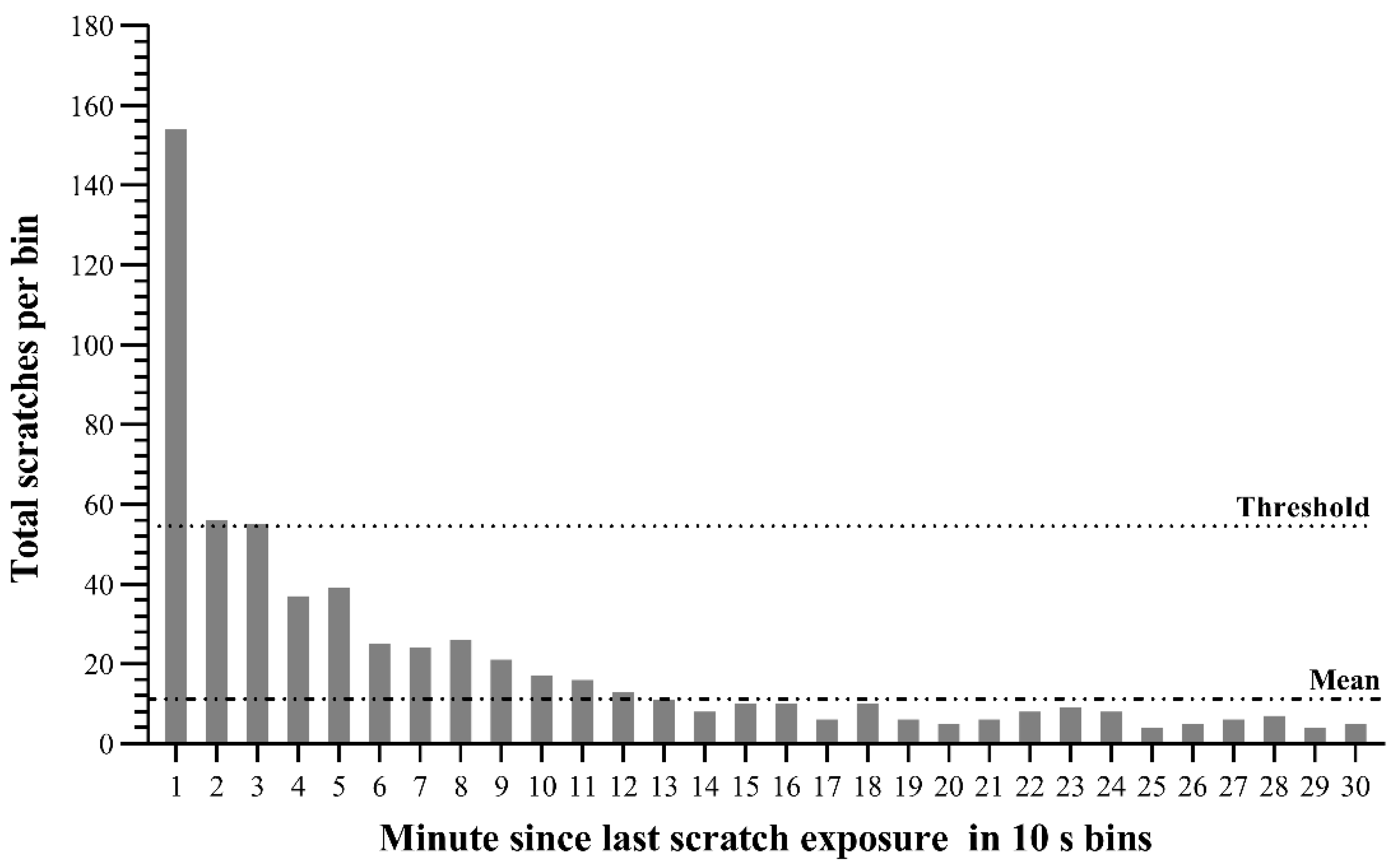
3.1. Scratch Contagion Analysis
3.2. Factors Affecting Frequency of Scratch Contagion
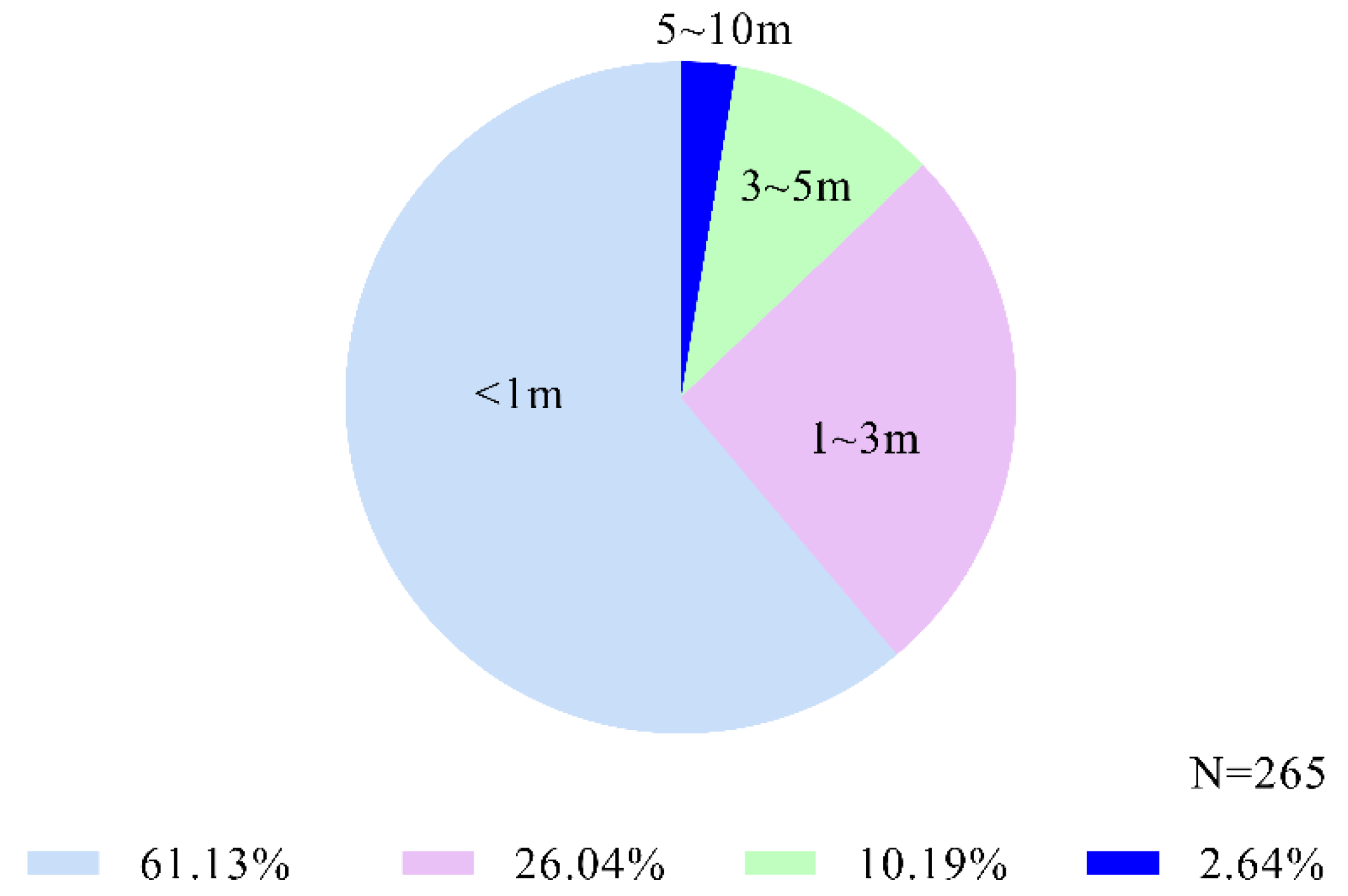
3.3. Strength of Scratch Contagion Changed with Distance between Expresser and Observer
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McDougall, P.L.; Ruckstuhl, K.E. Doing what your neighbour does: Neighbour proximity, familiarity and postural alignment increase behavioural mimicry. Anim. Behav. 2018, 135, 177–185. [Google Scholar] [CrossRef]
- Duranton, C.; Gaunet, F. Behavioural synchronization from an ethological perspective: Overview of its adaptive value. Adapt. Behav. 2016, 24, 181–191. [Google Scholar] [CrossRef]
- Chartrand, T.L.; Bargh, J.A. The chameleon effect: The perception–behavior link and social interaction. J. Pers. Soc. Psychol. 1999, 76, 893. [Google Scholar] [CrossRef]
- Lakin, J.L.; Jefferis, V.E.; Cheng, C.M.; Chartrand, T.L. The chameleon effect as social glue: Evidence for the evolutionary significance of nonconscious mimicry. J. Nonverbal Behav. 2003, 27, 145–162. [Google Scholar] [CrossRef]
- Zentall, T.R. Imitation by animals: How do they do it? Curr. Dir. Psychol. Sci. 2003, 12, 91–95. [Google Scholar] [CrossRef]
- Brooker, J.S.; Webb, C.E.; Clay, Z. Primate Empathy: A Flexible and Multi-Componential Phenomenon; Cambridge University Press: Cambridge, UK, 2021. [Google Scholar]
- Anderson, J.R.; Myowa-Yamakoshi, M.; Matsuzawa, T. Contagious yawning in chimpanzees. Proc. R. Soc. Lond. B 2004, 271 (Suppl. S6), S468–S470. [Google Scholar] [CrossRef]
- Campbell, M.W.; De Waal, F.B. Ingroup-outgroup bias in contagious yawning by chimpanzees supports link to empathy. PLoS ONE 2011, 6, e18283. [Google Scholar] [CrossRef] [PubMed]
- Campbell, M.W.; Carter, J.D.; Proctor, D.; Eisenberg, M.L.; de Waal, F.B. Computer animations stimulate contagious yawning in chimpanzees. Proc. R. Soc. Lond. B 2009, 276, 4255–4259. [Google Scholar] [CrossRef] [PubMed]
- Massen, J.J.; Vermunt, D.A.; Sterck, E.H. Male yawning is more contagious than female yawning among chimpanzees (Pan troglodytes). PLoS ONE 2012, 7, e40697. [Google Scholar] [CrossRef] [PubMed]
- Palagi, E.; Leone, A.; Mancini, G.; Ferrari, P.F. Contagious yawning in gelada baboons as a possible expression of empathy. Proc. Natl. Acad. Sci. USA 2009, 106, 19262–19267. [Google Scholar] [CrossRef]
- Palagi, E.; Norscia, I.; Demuru, E. Yawn contagion in humans and bonobos: Emotional affinity matters more than species. PeerJ 2014, 2, e519. [Google Scholar] [CrossRef] [PubMed]
- Demuru, E.; Palagi, E. In bonobos yawn contagion is higher among kin and friends. PLoS ONE 2012, 7, e49613. [Google Scholar] [CrossRef] [PubMed]
- Preston, S.D.; de Waal, F. The Communication of Emotions and the Possibility of Empathy in Animals; Oxford University Press: Oxford, UK, 2002; pp. 284–308. [Google Scholar]
- Norscia, I.; Demuru, E.; Palagi, E. She more than he: Gender bias supports the empathic nature of yawn contagion in Homo sapiens. Roy. Soc. Open Sci. 2016, 3, 150459. [Google Scholar] [CrossRef] [PubMed]
- Amici, F.; Aureli, F.; Call, J. Response facilitation in the four great apes: Is there a role for empathy? Primates 2014, 55, 113–118. [Google Scholar] [CrossRef]
- Paukner, A.; Anderson, J.R. Video-induced yawning in stumptail macaques (Macaca arctoides). Biol. Lett. 2006, 2, 36–38. [Google Scholar] [CrossRef]
- Reddy, R.B.; Krupenye, C.; MacLean, E.L.; Hare, B. No evidence for contagious yawning in lemurs. Anim. Cogn. 2016, 19, 889–898. [Google Scholar] [CrossRef]
- Massen, J.J.; Gallup, A.C. Why contagious yawning does not (yet) equate to empathy. Neurosci. Biobehav. Rev. 2017, 80, 573–585. [Google Scholar] [CrossRef]
- de Waal, F.B. Putting the altruism back into altruism: The evolution of empathy. Annu. Rev. Psychol. 2008, 59, 279–300. [Google Scholar] [CrossRef]
- Palagi, E.; Norscia, I.; Cordoni, G. Lowland gorillas (Gorilla gorilla gorilla) failed to respond to others’ yawn: Experimental and naturalistic evidence. J. Comp. Psychol. 2019, 133, 406. [Google Scholar] [CrossRef]
- Van Berlo, E.; Díaz-Loyo, A.P.; Juárez-Mora, O.E.; Kret, M.E.; Massen, J.J. Experimental evidence for yawn contagion in orangutans (Pongo pygmaeus). Sci. Rep. 2020, 10, 22251. [Google Scholar] [CrossRef]
- Miller, M.L.; Gallup, A.C.; Vogel, A.R.; Vicario, S.M.; Clark, A.B. Evidence for contagious behaviors in budgerigars (Melopsittacus undulatus): An observational study of yawning and stretching. Behav. Processes 2012, 89, 264–270. [Google Scholar] [CrossRef] [PubMed]
- Laméris, D.W.; van Berlo, E.; Sterck, E.H.; Bionda, T.; Kret, M.E. Low relationship quality predicts scratch contagion during tense situations in orangutans (Pongo pygmaeus). Am. J. Primatol. 2020, 82, e23138. [Google Scholar] [CrossRef] [PubMed]
- Maestripieri, D.; Schino, G.; Aureli, F.; Troisi, A. A modest proposal: Displacement activities as an indicator of emotions in primates. Anim. Behav. 1992, 44, 967–979. [Google Scholar] [CrossRef]
- Troisi, A. Ethological research in clinical psychiatry: The study of nonverbal behavior during interviews. Neurosci. Biobehav. Rev. 1999, 23, 905–913. [Google Scholar] [CrossRef]
- Gallup, A.C. On the link between emotional contagion and contagious yawning. Neurosci. Biobehav. Rev. 2021, 121, 18–19. [Google Scholar] [CrossRef] [PubMed]
- Duranton, C.; Bedossa, T.; Gaunet, F. Pet dogs synchronize their walking pace with that of their owners in open outdoor areas. Anim. Cogn. 2018, 21, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Rauchbauer, B.; Majdandzic, J.; Stieger, S.; Lamm, C. The Modulation of Mimicry by Ethnic Group-Membership and Emotional Expressions. PLoS ONE 2016, 11, e0161064. [Google Scholar] [CrossRef]
- Whitehouse, J.; Micheletta, J.; Waller, B.M. Stress behaviours buffer macaques from aggression. Sci. Rep. 2017, 7, 11083. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.X.; Li, J.H.; Xia, D.P.; Zhu, Y.; Wang, X.; Zhang, D. Influence of dominance rank and affiliation relationships on self-directed behavior in female Tibetan macaques (Macaca thibetana). Zool. Res. 2014, 35, 214–221. [Google Scholar] [CrossRef]
- Li, J.H. The Tibetan Macaque Society: A Field Study; Anhui University Press: Hefei, China, 1999. [Google Scholar]
- Berman, C.M.; Ionica, C.S.; Li, J. Dominance style among Macaca thibetana on Mt. Huangshan, China. Int. J. Primatol. 2004, 25, 1283–1312. [Google Scholar] [CrossRef]
- Wright, K.R.; Mayhew, J.A.; Sheeran, L.K.; Funkhouser, J.A.; Wagner, R.S.; Sun, L.X.; Li, J.H. Playing it cool: Characterizing social play, bout termination, and candidate play signals of juvenile and infant Tibetan macaques (Macaca thibetana). Zool. Res. 2018, 39, 272–283. [Google Scholar] [CrossRef] [PubMed]
- Xia, D.; Li, J.; Garber, P.A.; Sun, L.; Zhu, Y.; Sun, B. Grooming reciprocity in female tibetan macaques macaca thibetana. Am. J. Primatol. 2012, 74, 569–579. [Google Scholar] [CrossRef] [PubMed]
- Berman, C.M.; Li, J.-H. Impact of translocation, provisioning and range restriction on a group of Macaca thibetana. Int. J. Primatol. 2002, 23, 383–397. [Google Scholar] [CrossRef]
- Berman, C.M.; Ogawa, H.; Ionica, C.; Yin, H.; Li, J. Variation in kin bias over time in a group of Tibetan macaques at Huangshan, China: Contest competition, time constraints or risk response? Behaviour 2008, 145, 863–896. [Google Scholar] [CrossRef]
- Wang, X.; Xia, D.P.; Sun, L.; Garber, P.A.; Kyes, R.C.; Sheeran, L.K.; Sun, B.H.; Li, B.W.; Li, J.H. Infant attraction: Why social bridging matters for female leadership in Tibetan macaques. Curr. Zool. 2020, 66, 635–642. [Google Scholar] [CrossRef] [PubMed]
- Altmann, J. Observational study of behavior: Sampling methods. Behaviour 1974, 49, 227–267. [Google Scholar] [CrossRef] [PubMed]
- Berman, C.M.; Ionica, C.S.; Dorner, M.; Li, J. Postconflict affiliation between former opponents in Macaca thibetana on Mt. Huangshan, China. Int. J. Primatol. 2006, 27, 827–854. [Google Scholar] [CrossRef]
- Massen, J.J.; Slipogor, V.; Gallup, A.C. An Observational Investigation of Behavioral Contagion in Common Marmosets (Callithrix jacchus): Indications for Contagious Scent-Marking. Front. Psychol. 2016, 7, 1190. [Google Scholar] [CrossRef]
- Silk, J.B.; Altmann, J.; Alberts, S.C.; Sociobiology. Social relationships among adult female baboons (Papio cynocephalus) I. Variation in the strength of social bonds. Behav. Ecol. Sociobiol. 2006, 61, 183–195. [Google Scholar] [CrossRef]
- Silk, J.; Cheney, D.; Seyfarth, R. A practical guide to the study of social relationships. Evol. Anthropol. 2013, 22, 213–225. [Google Scholar] [CrossRef]
- Sueur, C.; Petit, O. Signals use by leaders in Macaca tonkeana and Macaca mulatta: Group-mate recruitment and behaviour monitoring. Anim. Cogn. 2010, 13, 239–248. [Google Scholar] [CrossRef] [PubMed]
- Gammell, M.P.; De Vries, H.; Jennings, D.J.; Carlin, C.o.M.; Hayden, T.J. David’s score: A more appropriate dominance ranking method than Clutton-Brock et al.’s index. Anim. Behav. 2003, 66, 601–605. [Google Scholar] [CrossRef]
- Gallese, V.; Fadiga, L.; Fogassi, L.; Rizzolatti, G. Action recognition in the premotor cortex. Brain 1996, 119 Pt 2, 593–609. [Google Scholar] [CrossRef] [PubMed]
- Preston, S.D.; de Waal, F.B. Empathy: Its ultimate and proximate bases. Behav. Brain. Sci. 2002, 25, 1–20; discussion 20–71. [Google Scholar] [CrossRef] [PubMed]
- Feneran, A.N.; O’Donnell, R.; Press, A.; Yosipovitch, G.; Cline, M.; Dugan, G.; Papoiu, A.D.P.; Nattkemper, L.A.; Chan, Y.H.; Shively, C.A. Monkey See, Monkey Do: Contagious Itch in Nonhuman Primates. Acta Derm-Venereol 2013, 93, 27–29. [Google Scholar] [CrossRef] [PubMed]
- Scopa, C.; Palagi, E. Mimic me while playing! Social tolerance and rapid facial mimicry in macaques (Macaca tonkeana and Macaca fuscata). J. Comp. Psychol. 2016, 130, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Whitehouse, J.; Micheletta, J.; Kaminski, J.; Waller, B.M. Macaques attend to scratching in others. Anim. Behav. 2016, 122, 169–175. [Google Scholar] [CrossRef]
- Paukner, A.; Suomi, S.J.; Visalberghi, E.; Ferrari, P.F. Capuchin monkeys display affiliation toward humans who imitate them. Science 2009, 325, 880–883. [Google Scholar] [CrossRef]
- Paukner, A.; Anderson, J.R.; Borelli, E.; Visalberghi, E.; Ferrari, P.F. Macaques (Macaca nemestrina) recognize when they are being imitated. Biol. Lett. 2005, 1, 219–222. [Google Scholar] [CrossRef]
- Bailenson, J.N.; Yee, N. Digital chameleons: Automatic assimilation of nonverbal gestures in immersive virtual environments. Psychol. Sci. 2005, 16, 814–819. [Google Scholar] [CrossRef] [PubMed]
- Dunbar, R.I.; Shultz, S. Bondedness and sociality. Behaviour 2010, 147, 775–803. [Google Scholar]
- Tang, Z.; Wang, X.; Wu, M.; Chen, S.; Li, J. Tibetan Macaques with Higher Social Centrality and More Relatives Emit More Frequent Visual Communication in Collective Decision-Making. Animals 2021, 11, 876. [Google Scholar] [CrossRef] [PubMed]
- Lewis, L.S.; Kano, F.; Stevens, J.M.; DuBois, J.G.; Call, J.; Krupenye, C. Bonobos and chimpanzees preferentially attend to familiar members of the dominant sex. Anim. Behav. 2021, 177, 193–206. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Catalog | Definition |
|---|---|
| Behavioral contagion | Self-scratching and yawning of individuals whose own behavior is repeated by other individuals for some time; the former is called the expresser, and the latter is called the observer. |
| Self-scratching | Movement of the hand or foot during which the fingertips are drawn across fur or skin. Not recorded as self-scratching if accompanied by fiddling with the fur carefully. |
| Yawning | Brief gaping movement of the mouth. Not recorded as yawning if accompanied by aggressive signals such as eye flash or canine whetting. |
| Some other social behaviors | |
| Social grooming | One individual orally or manually manipulates the fur of another. |
| Contact sitting | Two or more individuals are sitting or huddling in close body contact lasting more than 5 s. |
| Proximity (<1/3/5 m) | Two or more individuals keep a sitting or lying posture within a certain distance. In this study, the distances of 1, 3 and 5 m were recorded, respectively. |
| Aggression | An individual stares, hits on the ground, chases or orbits another individual. |
| Submission | An individual is attacked by another, but quickly leaves or flees in the opposite direction. |
| Factors | Coefficients | SE | Z | p |
|---|---|---|---|---|
| Relationship quality | 0.019 | 0.002 | 8.49 | <0.001 |
| Kinship | −0.328 | 0.203 | −1.62 | 0.105 |
| Sex-combination | 0.128 | 0.142 | 0.901 | 0.368 |
| Rank distance | 0.125 | 0.301 | 0.415 | 0.367 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.-H.; Wang, X.; Chen, M.-M.; Tai, Y.-M.; Li, J.-H. “Emotional Proximity” and “Spatial Proximity”: Higher Relationship Quality and Nearer Distance Both Strengthen Scratch Contagion in Tibetan Macaques. Animals 2022, 12, 2151. https://doi.org/10.3390/ani12162151
Zhang Y-H, Wang X, Chen M-M, Tai Y-M, Li J-H. “Emotional Proximity” and “Spatial Proximity”: Higher Relationship Quality and Nearer Distance Both Strengthen Scratch Contagion in Tibetan Macaques. Animals. 2022; 12(16):2151. https://doi.org/10.3390/ani12162151
Chicago/Turabian StyleZhang, Yu-Heng, Xi Wang, Meng-Meng Chen, Yi-Mei Tai, and Jin-Hua Li. 2022. "“Emotional Proximity” and “Spatial Proximity”: Higher Relationship Quality and Nearer Distance Both Strengthen Scratch Contagion in Tibetan Macaques" Animals 12, no. 16: 2151. https://doi.org/10.3390/ani12162151