
Individual Variation in the Use of Acoustic Signals to Coordinate Group Movements among Tibetan Macaques (Macaca thibetana)


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site and Subjects
2.2. Data Collection
2.2.1. Behavioral Observations and Definitions
2.2.2. Vocalization Recording and Acoustic Analyses
2.3. Data Analysis
2.4. Statistical Analysis
2.5. Ethics Statement
3. Results
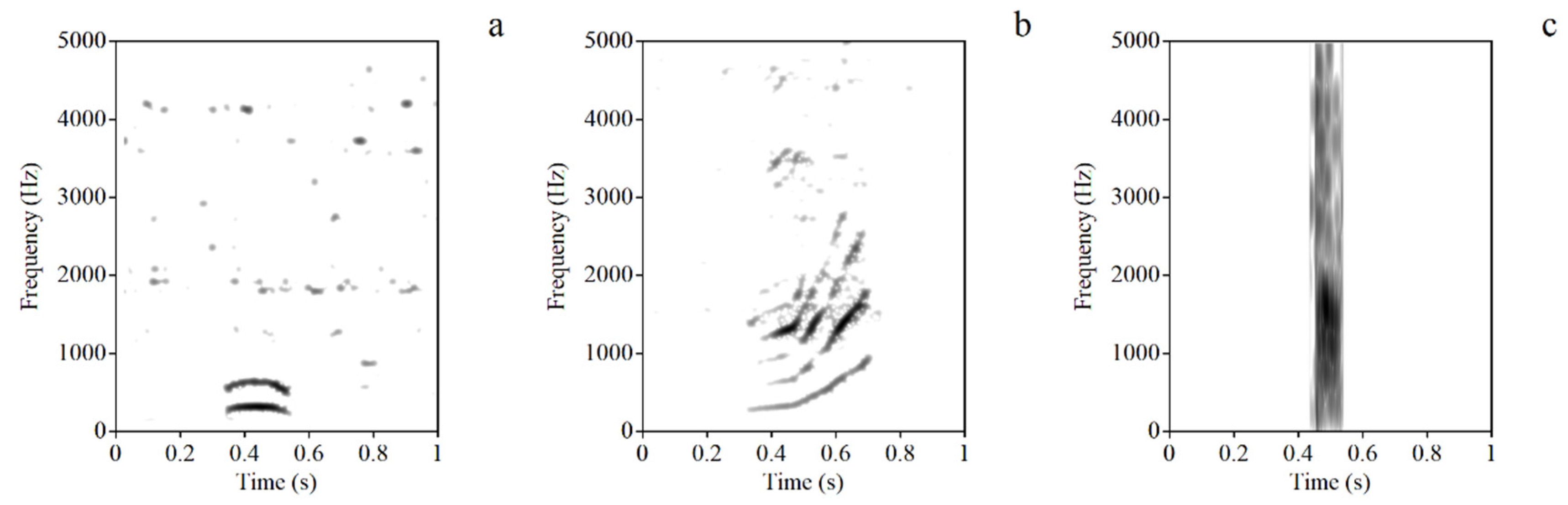
3.1. The Types of Sound Signals during Group Movements
3.2. The Usage of Vocal Communication during Group Movements
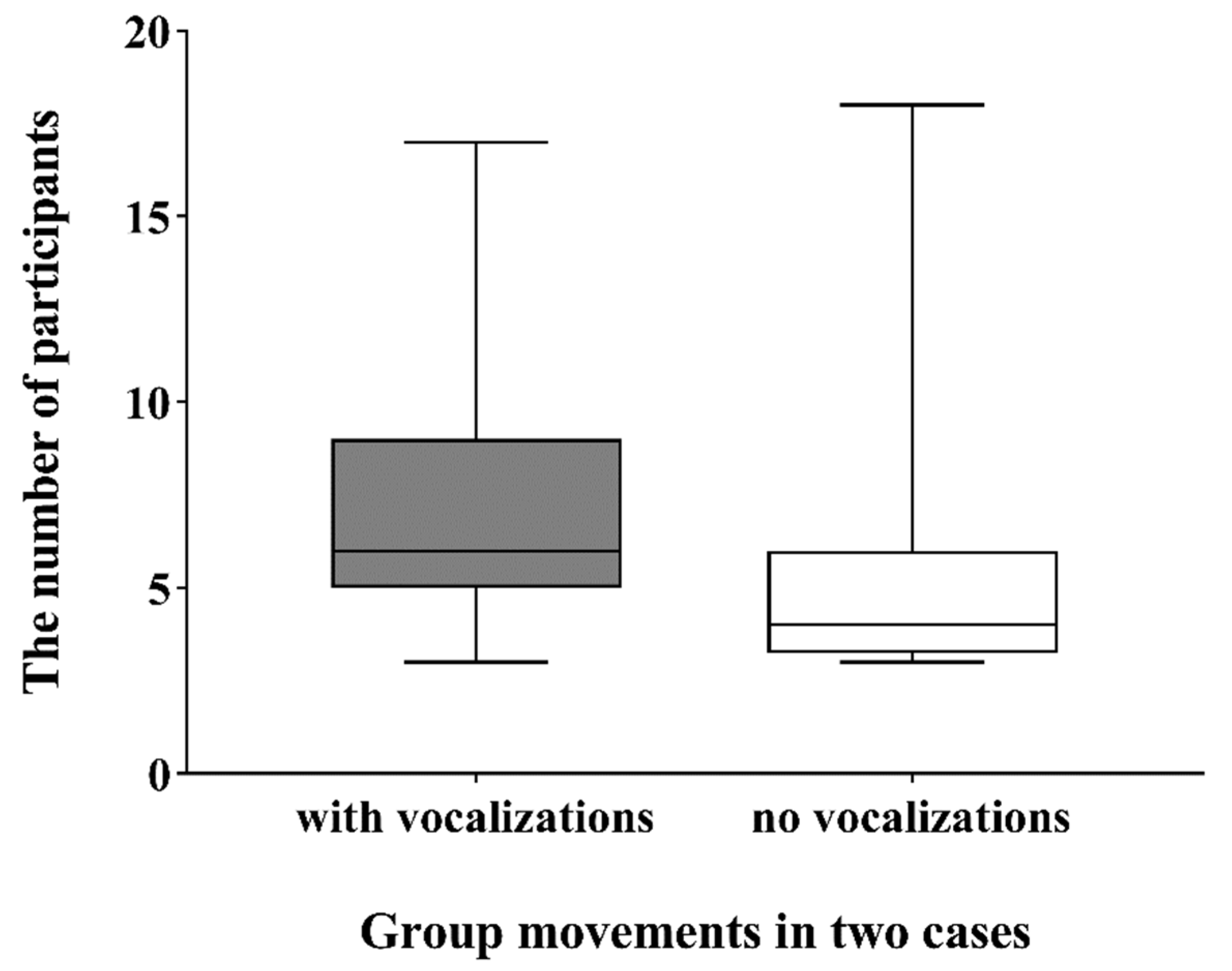
3.3. The Number of Participants during Group Movements
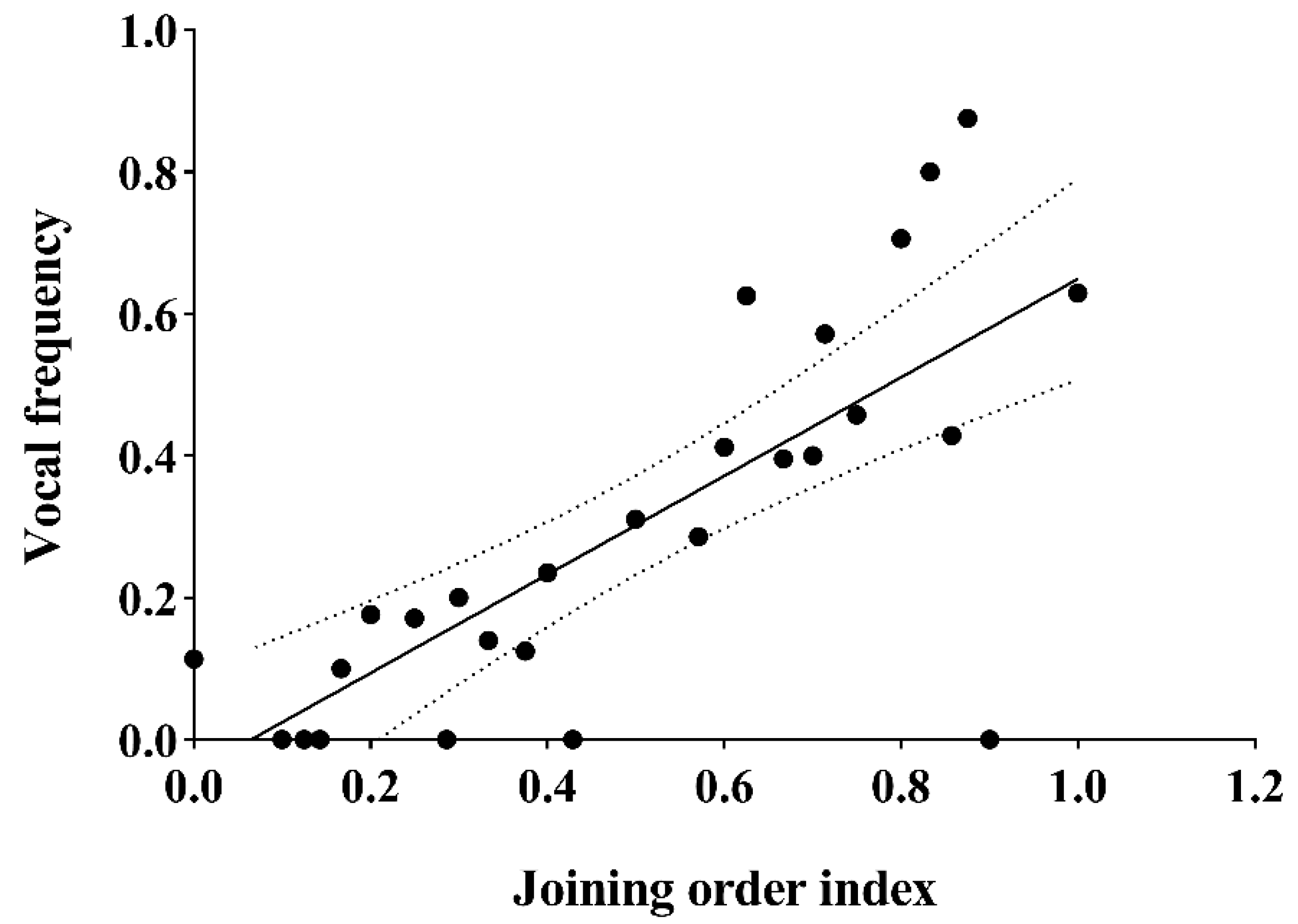
3.4. Joining Order and Vocalization
3.5. Social Factors and Vocalization
4. Discussion
4.1. Recruitment Function of Acoustic Signals
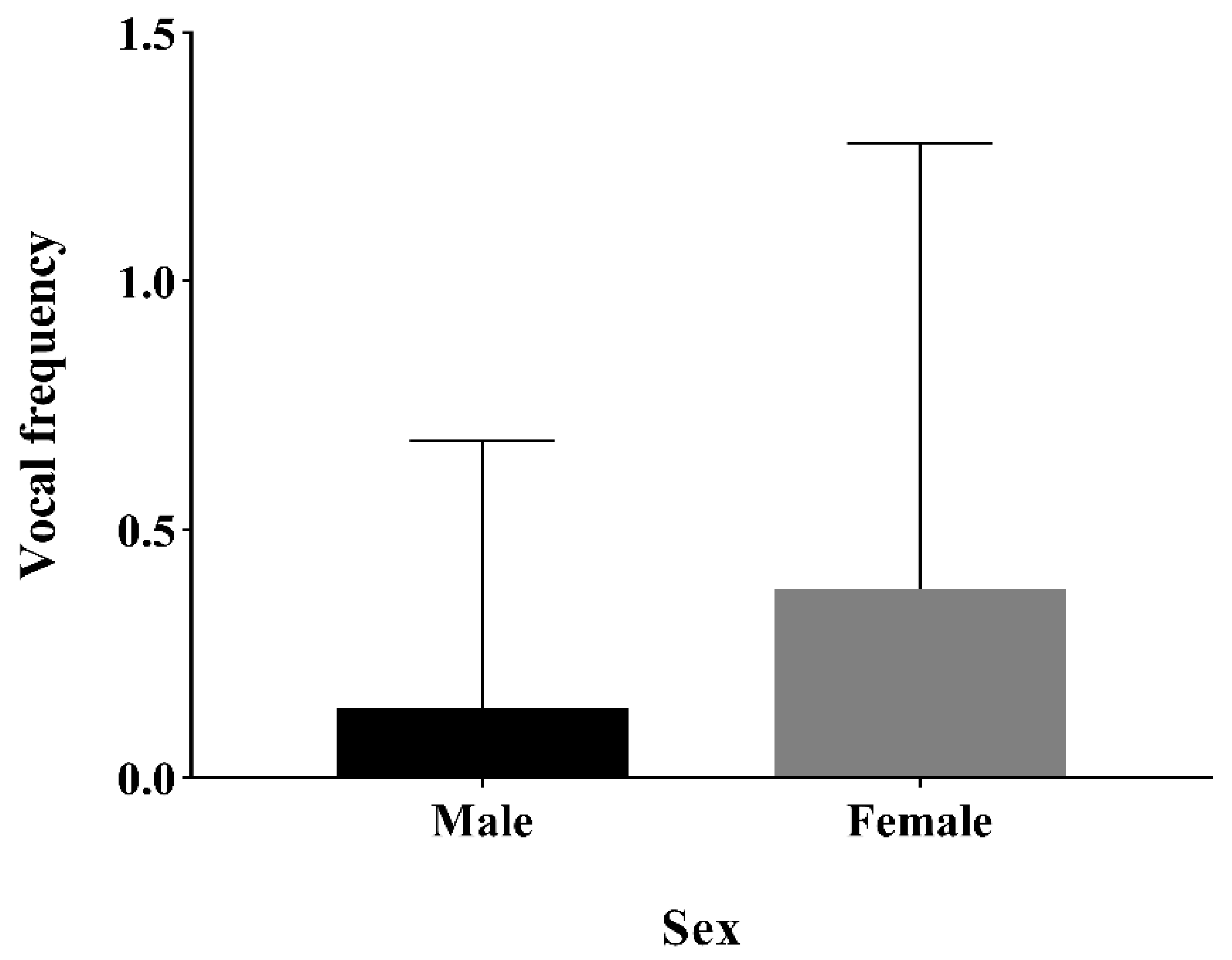
4.2. Influence of Sex and Social Factors on Frequency of Acoustic Signals
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Majolo, B.; de Bortoli Vizioli, A.; Schino, G. Costs and benefits of group living in primates: Group size effects on behaviour and demography. Anim. Behav. 2008, 76, 1235–1247. [Google Scholar] [CrossRef]
- Getz, L.L.; Carter, C.S. Prairie-vole partnerships. Am. Sci. 1996, 84, 56–62. [Google Scholar]
- Shimada, M.; Uno, T.; Nakagawa, N.; Fujita, S.; Izawa, K. Case study of a one-sided attack by multiple troop members on a nontroop adolescent male and the death of Japanese macaques (Macaca fuscata). Aggress. Behav. Off. J. Int. Soc. Res. Aggress. 2009, 35, 334–341. [Google Scholar] [CrossRef] [PubMed]
- Camazine, S.; Deneubourg, J.L.; Franks, N.R.; Sneyd, J.; Theraula, G.; Bonabeau, E. Self-organization in biological systems. In Self-Organization in Biological Systems; Princeton University Press: Princeton, NJ, USA, 2020; pp. 155–202. [Google Scholar]
- Fischer, J.; Zinner, D. Communication and cognition in primate group movement. Int. J. Primatol. 2011, 32, 1279–1295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alma, A.M.; Farji Brener, A.G.; Elizalde, L. Collective response of leaf-cutting ants to the effects of wind on foraging activity. Am. Nat. 2016, 188, 576–581. [Google Scholar] [CrossRef] [Green Version]
- Seltmann, A.; Franz, M.; Majolo, B.; Qarro, M.; Ostner, J.; Schülke, O. Recruitment and monitoring behaviors by leaders predict following in wild Barbary macaques (Macaca sylvanus). Primate Biol. 2016, 3, 23–31. [Google Scholar] [CrossRef] [Green Version]
- Fan, P.L.; Liu, X.C.; Liu, R.S.; Li, F.; Huang, T.P.; Wu, F.; Yao, H.; Liu, D.Z. Vocal repertoire of free-ranging adult golden snub-nosed monkeys (Rhinopithecus roxellana). Am. J. Primatol. 2018, 80, e22869. [Google Scholar] [CrossRef] [Green Version]
- Engesser, S.; Manser, M.B. Collective close calling mediates group cohesion in foraging meerkats via spatially determined differences in call rates. Anim. Behav. 2022, 185, 73–82. [Google Scholar] [CrossRef]
- Gall, G.E.; Manser, M.B. Group cohesion in foraging meerkats: Follow the moving ‘vocal hot spot’. R. Soc. Open Sci. 2017, 4, 170004. [Google Scholar] [CrossRef] [Green Version]
- Walker, R.H.; King, A.J.; McNutt, J.W.; Jordan, N.R. Sneeze to leave: African wild dogs (Lycaon pictus) use variable quorum thresholds facilitated by sneezes in collective decisions. Proc. R. Soc. B Biol. Sci. 2017, 284, 20170347. [Google Scholar] [CrossRef] [Green Version]
- Warrington, M.H.; McDonald, P.G.; Sager, A.K.; Griffith, S.C. The vocal repertoire of the cooperatively breeding Apostlebird (Struthidea cinerea). Emu Austral Ornithol. 2014, 114, 206–221. [Google Scholar] [CrossRef]
- Jürgens, U.; Newman, J.; Zimmermann, E. Current Topics in Primate Vocal Communication; Springer Science & Business Media: New York, NY, USA, 2013. [Google Scholar]
- Fischer, J. Transmission of acquired information in nonhuman primates. In Learning and Memory: A Comprehensive Reference; Elsevier: Oxford, UK, 2008; pp. 299–313. [Google Scholar]
- Sperber, A.L.; Werner, L.M.; Kappeler, P.M.; Fichtel, C. Grunt to go—Vocal coordination of group movements in redfronted lemurs. Ethology 2017, 123, 894–905. [Google Scholar] [CrossRef]
- Campbell, A.F.; Boinski, S. Use of trill vocalizations to coordinate troop movement among white-faced capuchins: A second field test. Behaviour 1995, 132, 875–901. [Google Scholar] [CrossRef]
- Fischer, J.; Hammerschmidt, K. An overview of the Barbary macaque, Macaca sylvanus, vocal repertoire. Folia Primatol. 2002, 73, 32–45. [Google Scholar] [CrossRef]
- Riley, E. The loud call of the Sulawesi Tonkean macaque, Macaca tonkeana. Trop. Biodivers. 2005, 8, 199–209. [Google Scholar]
- Schamberg, I.; Cheney, D.L.; Clay, Z.; Hohmann, G.; Seyfarth, R.M. Bonobos use call combinations to facilitate inter-party travel recruitment. Behav. Ecol. Sociobiol. 2017, 71, 1–8. [Google Scholar] [CrossRef]
- Bernstein, S.K.; Sheeran, L.K.; Wagner, R.S.; Li, J.H.; Koda, H. The vocal repertoire of Tibetan macaques (Macaca thibetana): A quantitative classification. Am. J. Primatol. 2016, 78, 937–949. [Google Scholar] [CrossRef]
- Tang, Z.F.; Wang, X.; Wu, M.Y.; Chen, S.W.; Li, J.H. Tibetan Macaques with Higher Social Centrality and More Relatives Emit More Frequent Visual Communication in Collective Decision-Making. Animals 2021, 11, 876. [Google Scholar] [CrossRef]
- Li, J.H. The Tibetan Macaque Society: A Field Study; Anhui University Press: Heifei, China, 1999. (In Chinese) [Google Scholar]
- Berman, C.M.; Li, J.H. Impact of translocation, provisioning and range restriction on a group of Macaca thibetana. Int. J. Primatol. 2002, 23, 383–397. [Google Scholar] [CrossRef]
- Gammell, M.P.; De Vries, H.; Jennings, D.J.; Carlin, C.o.M.; Hayden, T.J. David’s Score: A More Appropriate Dominance Ranking Method than Clutton-Brock et al.’s Index; Academic Press: Cambridge, MA, USA, 2003; Volume 66, pp. 601–605. [Google Scholar]
- Altmann, J. Observational study of behavior: Sampling methods. Behaviour 1974, 49, 227–266. [Google Scholar] [CrossRef] [Green Version]
- Berman, C.M.; Ionica, C.S.; Li, J.H. Dominance style among Macaca thibetana on Mt. Huangshan, China. Int. J. Primatol. 2004, 25, 1283–1312. [Google Scholar] [CrossRef]
- Rowe, A.K.; Li, J.H.; Sun, L.; Sheeran, L.K.; Wagner, R.S.; Xia, D.P.; Uhey, D.A.; Chen, R. Collective decision making in Tibetan macaques: How followers affect the rules and speed of group movement. Anim. Behav. 2018, 146, 51–61. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Sun, L.; Sheeran, L.K.; Sun, B.H.; Zhang, Q.X.; Zhang, D.; Xia, D.P.; Li, J.H. Social rank versus affiliation: Which is more closely related to leadership of group movements in Tibetan macaques (Macaca thibetana)? Am. J. Primatol. 2016, 78, 816–824. [Google Scholar] [CrossRef] [PubMed]
- Sueur, C.; Petit, O. Shared or unshared consensus decision in macaques? Behav. Process. 2008, 78, 84–92. [Google Scholar] [CrossRef]
- Pyritz, L.W.; King, A.J.; Sueur, C.; Fichtel, C. Reaching a consensus: Terminology and concepts used in coordination and decision-making research. Int. J. Primatol. 2011, 32, 1268–1278. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, A.; Watanabe, K.; Petit, O. Social structure affects initiations of group movements but not recruitment success in Japanese macaques (Macaca fuscata). Int. J. Primatol. 2011, 32, 1311–1324. [Google Scholar] [CrossRef]
- Tenaza, R.; Fitch, H.; Lindburg, D. Vocal behavior of captive Sichuan golden monkeys (Rhinopithecus r. roxellana). Am. J. Primatol. 1988, 14, 1–9. [Google Scholar] [CrossRef]
- Barelli, C.; Boesch, C.; Heistermann, M.; Reichard, U.H. Female white-handed gibbons (Hylobates lar) lead group movements and have priority of access to food resources. Behaviour 2008, 145, 965–981. [Google Scholar] [CrossRef] [Green Version]
- Nishida, T. The social group of wild chimpanzees in the Mahali Mountains. Primates 1968, 9, 167–224. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.X.; Li, J.H.; Xia, D.P.; Zhu, Y.; Wang, X.; Zhang, D. Influence of dominance rank and affiliation relationships on selfdirected behavior in female Tibetan macaques (Macaca thibetana). Zool. Res. 2014, 35, 214. [Google Scholar]
- Sueur, C.; Petit, O. Signals use by leaders in Macaca tonkeana and Macaca mulatta: Group-mate recruitment and behaviour monitoring. Anim. Cogn. 2010, 13, 239–248. [Google Scholar] [CrossRef]
- Bolker, B. Team R: Bbmle: Tools for General Maximum Likelihood Estimation. R package Version 0.9. 2010. Available online: https://www.google.com.hk/url?sa=t&rct=j&q=&esrc=s&source=web&cd=&cad=rja&uact=8&ved=2ahUKEwi9mvrOwdn5AhWJEogKHVPkAbgQFnoECAUQAQ&url=https%3A%2F%2Fcran.r-project.org%2Fweb%2Fpackages%2Fbbmle%2Fbbmle.pdf&usg=AOvVaw0DLh2PwCYlRKL_o5wTIYz1 (accessed on 1 July 2022).
- Meise, K.; Keller, C.; Cowlishaw, G.; Fischer, J. Sources of acoustic variation: Implications for production specificity and call categorization in chacma baboon (Papio ursinus) grunts. J. Acoust. Soc. Am. 2011, 129, 1631–1641. [Google Scholar] [CrossRef] [PubMed]
- Gruber, T.; Zuberbühler, K. Vocal recruitment for joint travel in wild chimpanzees. PLoS ONE 2013, 8, e76073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koski, S.E.; de Vries, H.; van de Kraats, A.; Sterck, E.H. Stability and change of social relationship quality in captive chimpanzees (Pan troglodytes). Int. J. Primatol. 2012, 33, 905–921. [Google Scholar] [CrossRef] [Green Version]
- Fratellone, G.P.; Li, J.H.; Sheeran, L.K.; Wagner, R.S.; Wang, X.; Sun, L.X. Social connectivity among female Tibetan macaques (Macaca thibetana) increases the speed of collective movements. Primates 2019, 60, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Xia, D.P.; Sun, L.; Garber, P.A.; Kyes, R.C.; Sheeran, L.K.; Sun, B.H.; Li, B.W.; Li, J.H. Infant attraction: Why social bridging matters for female leadership in Tibetan macaques. Curr. Zool. 2020, 66, 635–642. [Google Scholar] [CrossRef]
- Wang, X.; Sun, L.X.; Li, J.H.; Xia, D.P.; Sun, B.H.; Zhang, D. Collective movement in the Tibetan macaques (Macaca thibetana): Early joiners write the rule of the game. PLoS ONE 2015, 10, e0127459. [Google Scholar] [CrossRef]
- Bolt, L.M.; Tennenhouse, E. Contact calling behaviour in the male ring-tailed lemur (Lemur catta). Ethology 2017, 123, 614–626. [Google Scholar] [CrossRef]
- Sueur, C.; King, A.J.; Pelé, M.; Petit, O. Fast and accurate decisions as a result of scale-free network properties in two primate species. In Proceedings of the European Conference on Complex Systems 2012; Springer: Berlin/Heidelberg, Germany, 2013; pp. 579–584. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Individual | Sex | Number of Relatives | Social Rank | David’s Score # |
|---|---|---|---|---|
| YL | Male | 0 | 1 | 205.24 |
| YXK | Male | 6 | 2 | 176.94 |
| ZB | Male | 0 | 3 | 137.17 |
| YXX | Female | 6 | 4 | 125.5 |
| YH | Female | 6 | 5 | 82.63 |
| DB | Male | 0 | 6 | 81.58 |
| NM | Male | 0 | 7 | 71.93 |
| TQ | Male | 0 | 8 | 61.68 |
| WM | Male | 0 | 9 | 39.85 |
| YCY | Female | 6 | 10 | 28.62 |
| YXY | Female | 6 | 11 | 25.08 |
| YCH | Female | 0 | 12 | 2.04 |
| TXH | Female | 6 | 13 | −14.6 |
| YCL | Female | 6 | 14 | −23.26 |
| DZ | Male | 0 | 15 | −24.75 |
| TQS | Male | 6 | 16 | −29.32 |
| BHZ | Male | 0 | 17 | −41.48 |
| TH | Female | 6 | 18 | −48 |
| TXX | Female | 6 | 19 | −68.08 |
| TQL | Female | 6 | 20 | −76.17 |
| THY | Female | 2 | 21 | −85.9 |
| TQY | Female | 6 | 22 | −93.89 |
| QT | Male | 0 | 23 | −113.9 |
| THX | Female | 2 | 24 | −138.21 |
| YXM | Male | 6 | 25 | −145.67 |
| TFH | Female | 2 | 26 | −150.35 |
| TQG | Female | 6 | 27 | −153.02 |
| Catalog | Definition |
|---|---|
| Group movement | When no more individuals join the movement within five minutes after the joining of the last one, the number of participants, the initiator included, should be at least 3. |
| Initiator | The individual who first walks over 10 m within 30 s. |
| Follower | The individual who moves over 5 m within 45° along the direction of the initiator. |
| Proximity | At least two individuals maintain the sitting or lying posture within 1 m. |
| Parameters | Definitions |
|---|---|
| Duration | Duration of the entire call (s) |
| Mean f0 | Mean frequency of the fundamental frequency contour (Hz) |
| Min f0 | Minimum frequency of the fundamental frequency contour (Hz) |
| Max f0 | Maximum frequency of the fundamental frequency contour (Hz) |
| Mean AMP | Mean intensity (amplitude) of the entire call (dB) |
| Min AMP | Minimum intensity (amplitude) of the entire call (dB) |
| Max AMP | Maximum intensity (amplitude) of the entire call (dB) |
| Call Type | N | Duration(s) | Mean f0 | Min f0 | Max f0 | Mean AMP | Min AMP | Max AMP |
|---|---|---|---|---|---|---|---|---|
| Coo | 22 | 0.23 ± 0.04 | 282 ± 29 | 244 ± 36 | 303 ± 42 | 53 ± 10 | 43 ± 10 | 56 ± 10 |
| Leap coo | 7 | 0.26 ± 0.11 | 361 ± 15 | 260 ± 60 | 490 ± 45 | 58 ± 6 | 45 ± 5 | 63 ± 6 |
| Bark | 1 | 0.06 | 378 | 360 | 398 | 73 | 68 | 75 |
| Factors | Estimate ± SE | Z | p |
|---|---|---|---|
| Sex | 0.771 ± 0.283 | 2.723 | <0.01 |
| Rank | −0.007 ± 0.013 | 0.498 | 0.619 |
| Relatives | −0.001 ± 0.039 | −0.027 | 0.978 |
| Centrality | 3.361 ± 1.490 | 2.245 | <0.05 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, M.-M.; Zhang, Y.-H.; Tai, Y.-M.; Wang, X. Individual Variation in the Use of Acoustic Signals to Coordinate Group Movements among Tibetan Macaques (Macaca thibetana). Animals 2022, 12, 2149. https://doi.org/10.3390/ani12162149
Chen M-M, Zhang Y-H, Tai Y-M, Wang X. Individual Variation in the Use of Acoustic Signals to Coordinate Group Movements among Tibetan Macaques (Macaca thibetana). Animals. 2022; 12(16):2149. https://doi.org/10.3390/ani12162149
Chicago/Turabian StyleChen, Meng-Meng, Yu-Heng Zhang, Yi-Mei Tai, and Xi Wang. 2022. "Individual Variation in the Use of Acoustic Signals to Coordinate Group Movements among Tibetan Macaques (Macaca thibetana)" Animals 12, no. 16: 2149. https://doi.org/10.3390/ani12162149