
Maternal Supplementation with Polyphenols and Omega-3 Fatty Acids during Pregnancy: Prenatal Effects on Fetal Fatty Acid Composition in the Iberian Pig

 , , , , , and
, , , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethic Statement
2.2. Animals and Experimental Procedures
2.3. Sampling of Fetuses
2.4. Fatty Acid Composition of Plasma and Tissues
2.5. Statistical Analysis
3. Results
3.1. Fat Content of the Different Tissues
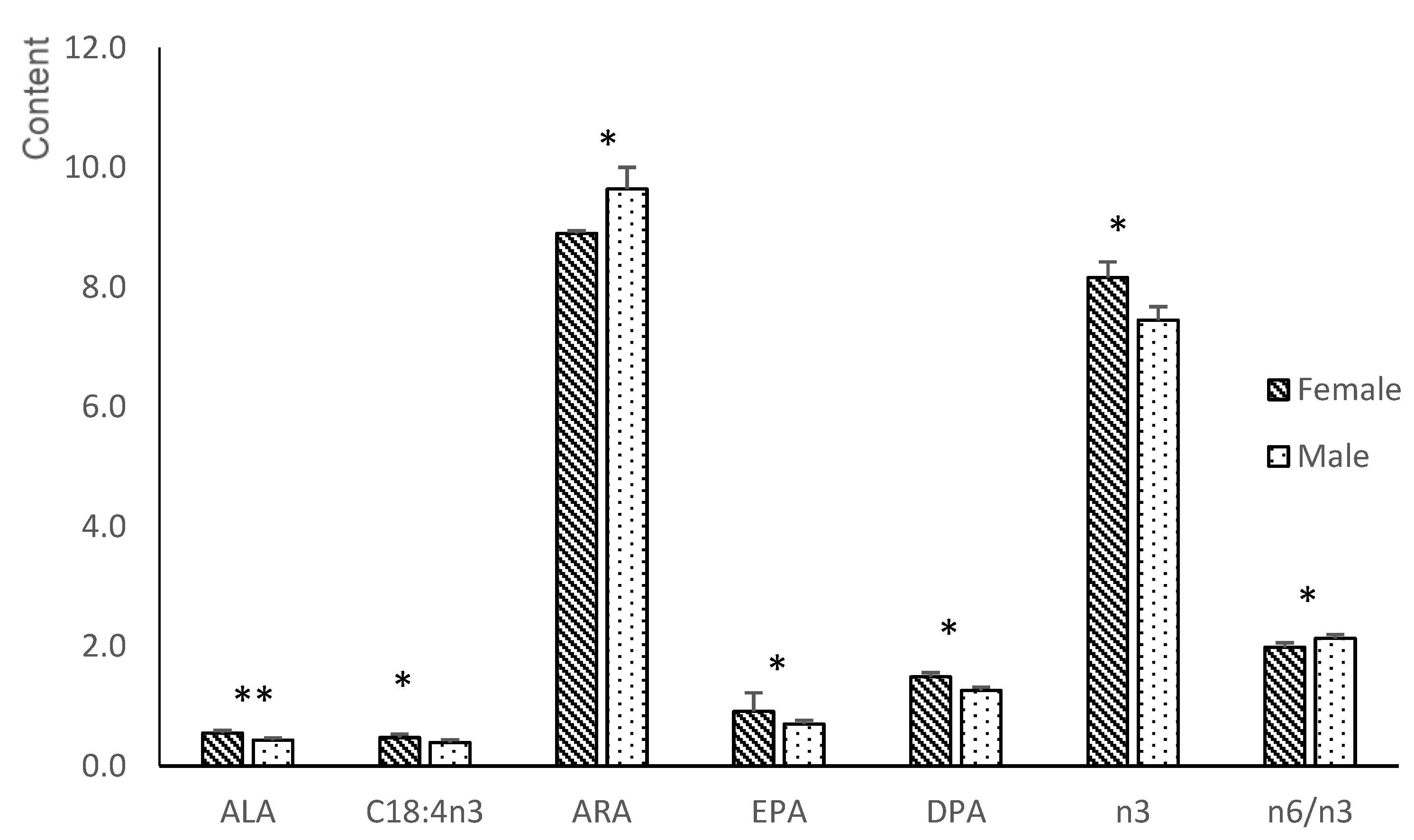
3.2. Fatty Acid Composition of the Plasma of the Gilts and Fetuses
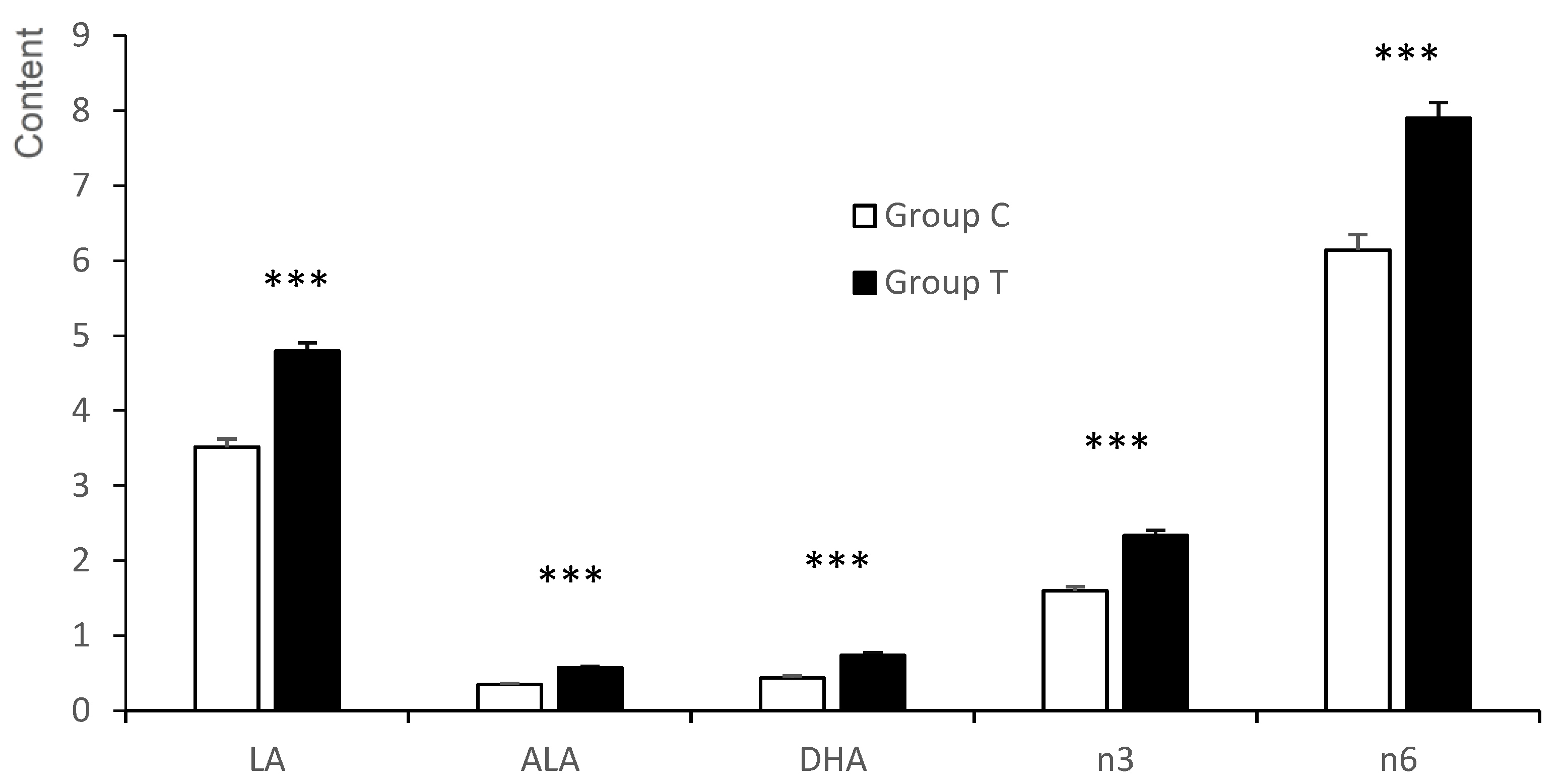
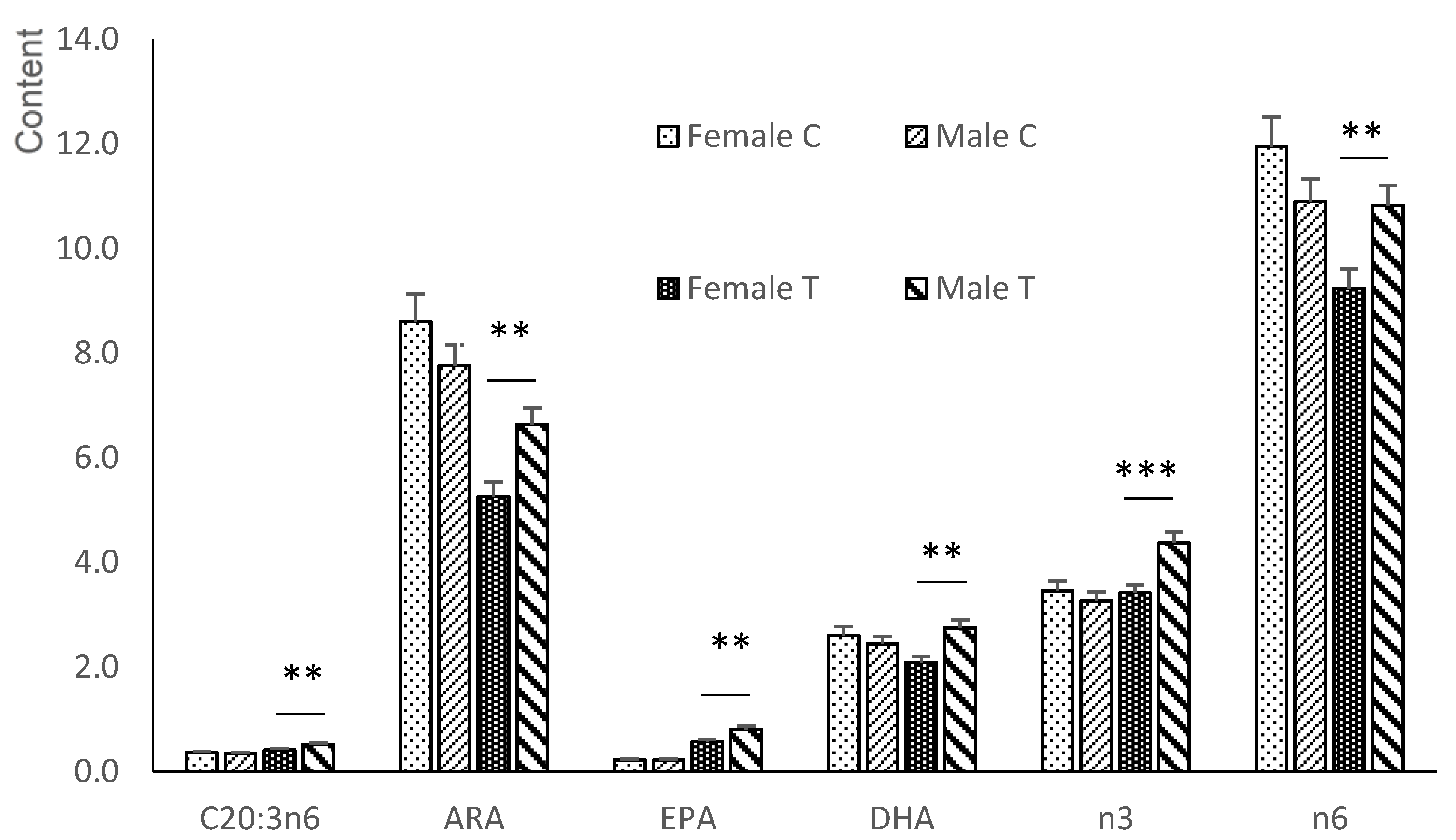
3.3. Fatty Acid Composition of the Longissimus Dorsi Muscle of the Fetuses
3.3.1. Neutral Fraction
3.3.2. Polar Fraction
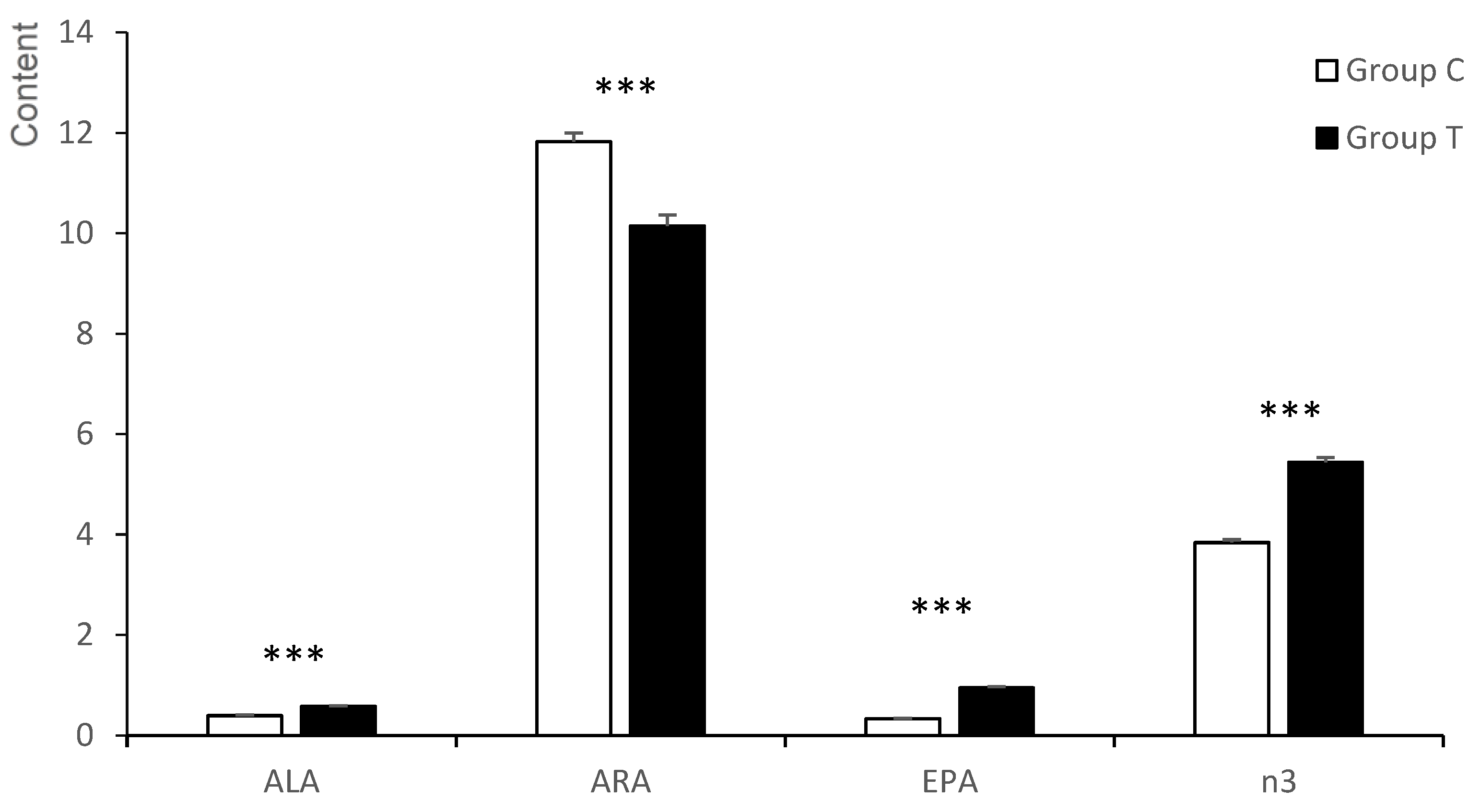
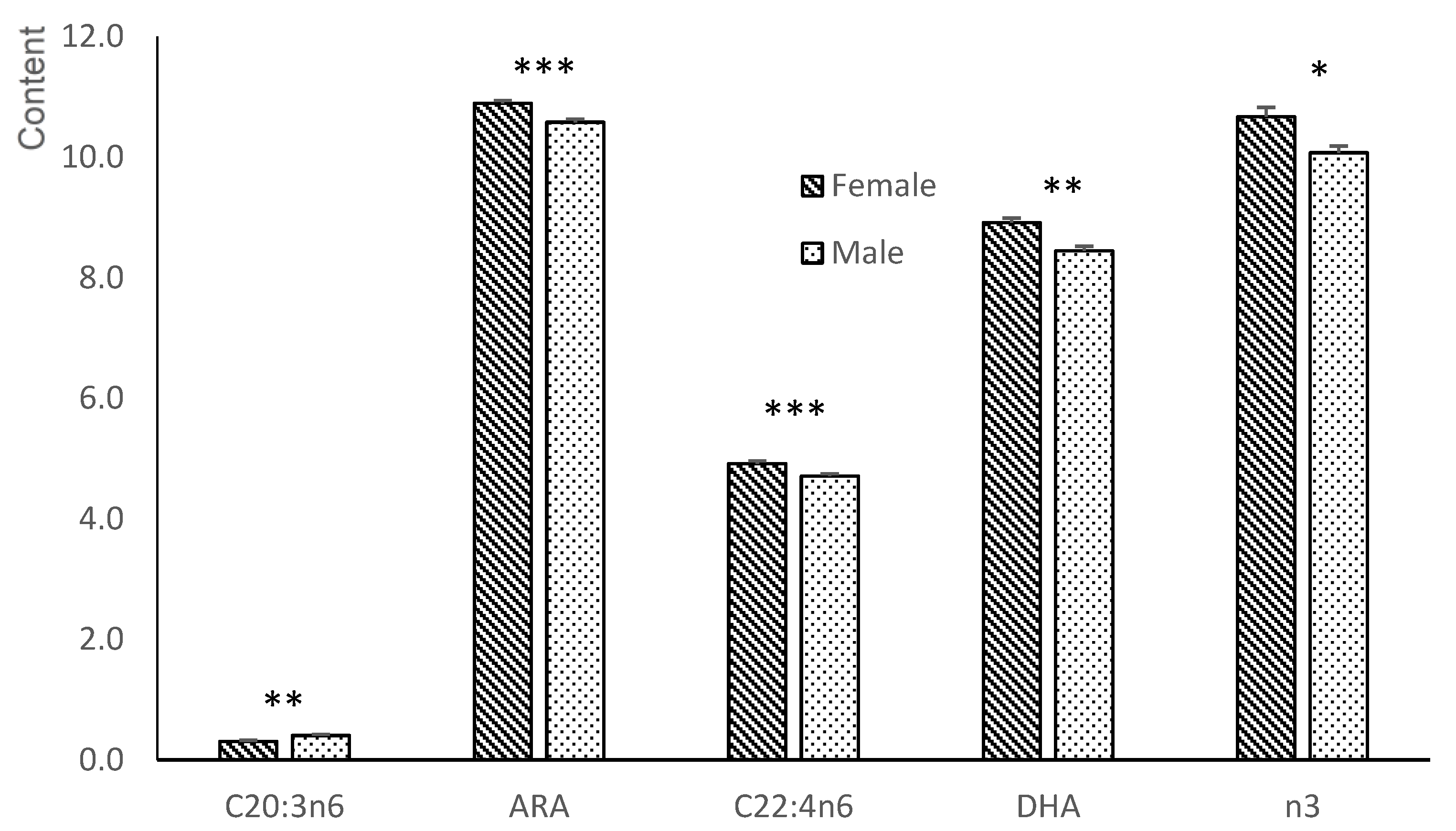
3.4. Fatty Acid Composition of the Liver of the Fetuses
3.4.1. Neutral Fraction
3.4.2. Polar Fraction
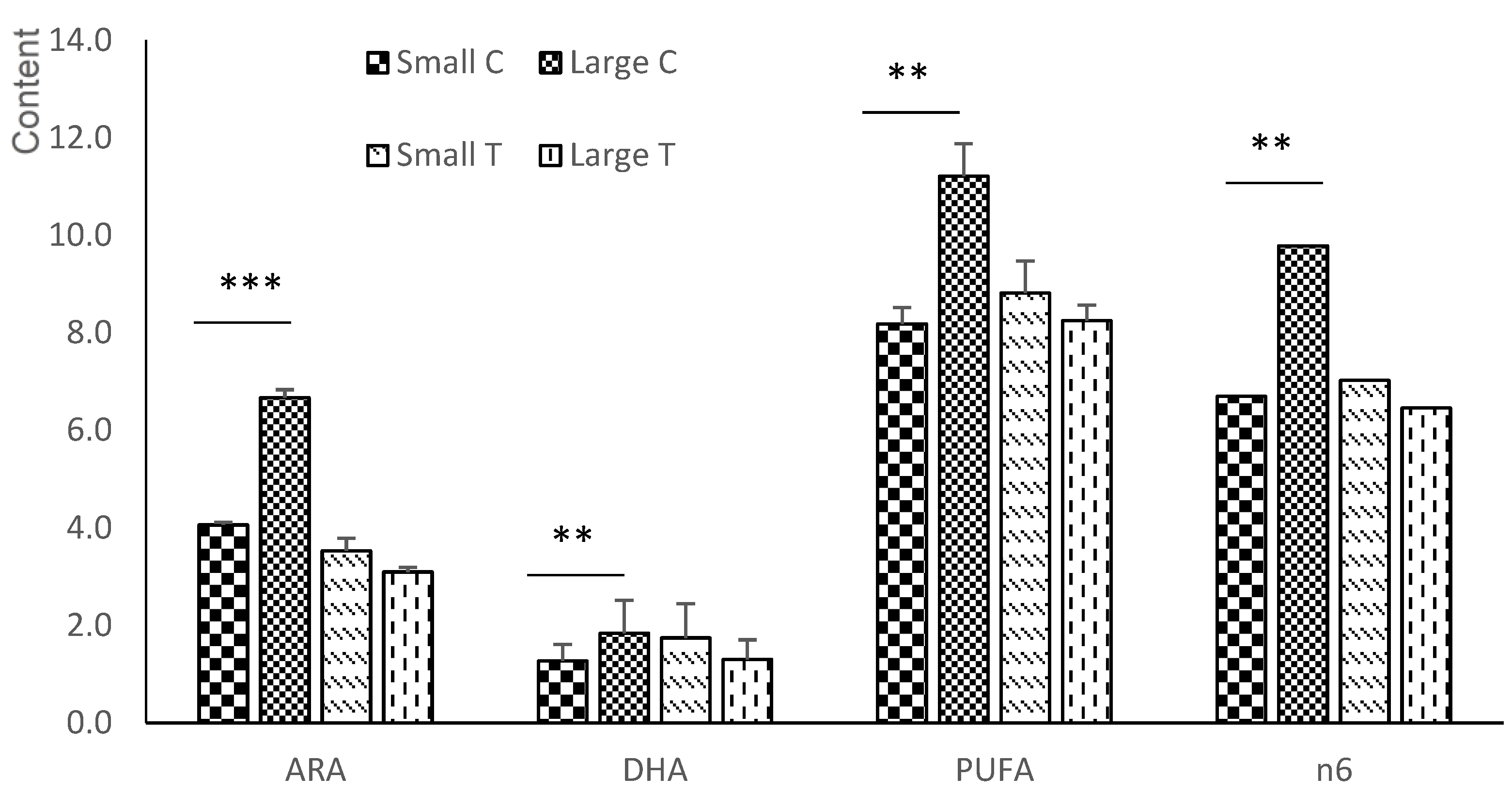
3.5. Fatty Acid Composition of the Brain
3.5.1. Neutral Fraction
3.5.2. Polar Fraction
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wu, G.; Bazer, F.W.; Cudd, T.A.; Meininger, C.J.; Spencer, T.E. Maternal nutrition and fetal development. J. Nutr. 2004, 134, 2169–2172. [Google Scholar] [CrossRef] [PubMed]
- Huting, A.M.S.; Middelkoop, A.; Guan, X.; Molist, F. Using nutritional strategies to shape the gastro-intestinal tracts of suckling and weaned piglets. Animals 2021, 11, 402. [Google Scholar] [CrossRef]
- Leskanich, C.O.; Noble, R.C. The comparative roles of polyunsaturated fatty acids in pig neonatal development. Br. J. Nutr. 1999, 81, 87–106. [Google Scholar] [CrossRef] [PubMed]
- Lavery, A.; Lawlor, P.G.; Miller, H.M.; Magowan, E. The effect of dietary oil type and energy intake in lactating sows on the fatty acid profile of colostrum and milk, and piglet growth to weaning. Animals 2019, 9, 1092. [Google Scholar] [CrossRef] [PubMed]
- Egert, S.; Baxheinrich, A.; Lee-Barkey, Y.H.; Tschoepe, D.; Wahrburg, U.; Stratmann, B. Effects of an energy-restricted diet rich in plant-derived α-linolenic acid on systemic inflammation and endothelial function in overweight-to-obese patients with metabolic syndrome traits. Br. J. Nutr. 2014, 112, 1315–1322. [Google Scholar] [CrossRef]
- Swiatkiewicz, S.; Arczewska-Wlosek, A.; Jozefiak, D. The relationship between dietary fat sources and immune response in poultry and pigs: An updated review. Livestock Sci. 2015, 180, 237–246. [Google Scholar] [CrossRef]
- Milligan, B.N.; Fraser, D.; Kramer, D.L. Within-litter birth weight variation in the domestic pig and its relation to pre-weaning survival, weight gain, and variation in weaning weights. Livest. Prod. Sci. 2002, 76, 181–191. [Google Scholar] [CrossRef]
- World Health Organization (W.H.O.). Interim summary of conclusions and dietary recommendations on total fat & fatty acids. In Proceedings of the Joint FAO/WHO Expert Consultation on Fats and Fatty Acids in Human Nutrition 2008, Geneva, Switzerland, 10–14 November 2008. [Google Scholar]
- Nuernberg, K.; Fischer, K.; Nuernberg, G.; Kuechenmeister, U.; Klosowska, D.; Eliminowska-Wenda, G.; Fiedler, I.; Ender, K. Effects of dietary olive and linseed oil on lipid composition, meat quality, sensory characteristics and muscle structure in pigs. Meat Sci. 2005, 70, 63–74. [Google Scholar] [CrossRef]
- Wood, J.D.; Enser, M.; Fisher, A.V.; Nute, G.R.; Richardson, R.I.; Sheard, P.R. Manipulating meat quality and composition. Proc. Nutr. Soc. 1999, 58, 363–370. [Google Scholar] [CrossRef]
- Amusquivar, E.; Rupérez, F.J.; Barbas, C.; Herrera, E. Low arachidonic acid rather than α-tocopherol is responsible for the delayed postnatal development in offspring of rats fed fish oil instead of olive oil during pregnancy and lactation. J. Nutr. 2000, 130, 2855–2865. [Google Scholar] [CrossRef]
- Thorsdottir, I.; Birgisdottir, B.E.; Halldorsdottir, S.; Geirsson, R.T. Association of fish and fish liver oil intake in pregnancy with infant size at birth among women of normal weight before pregnancy in a fishing community. Am. J. Epidemiol. 2004, 160, 460–465. [Google Scholar] [CrossRef]
- Cortinas, L.; Barroeta, A.; Villaverde, C.; Galobart, J.; Guardiola, F.; Baucells, M.D. Influence of the dietary polyunsaturation level on chicken meat quality: Lipid oxidation. Poult. Sci. 2005, 84, 48–55. [Google Scholar] [CrossRef]
- Valk, E.E.; Hornstra, G. Relationship between vitamin E requirement and polyunsaturated fatty acid intake in man: A review. Int. J. Vitam. Nutr. Res. 2000, 70, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Wani, T.A.; Masoodi, F.A.; Gani, A.; Baba, W.N.; Rahmanian, N.; Akhter, R.; Wani, I.A.; Ahmad, M. Olive oil and its principal bioactive compound: Hydroxytyrosol–A review of the recent literature. Trends Food Sci. Technol. 2018, 77, 77–90. [Google Scholar] [CrossRef]
- Garcia-Contreras, C.; Vazquez-Gomez, M.; Barbero, A.; Pesantez, J.; Zinellu, A.; Berlinguer, F.; Gonzalez-Añover, P.; Gonzalez, J.; Encinas, T.; Torres-Rovira, L.; et al. Polyphenols and IUGR Pregnancies: Effects of Maternal Hydroxytyrosol Supplementation on Placental Gene Expression and Fetal Antioxidant Status, DNA-Methylation and Phenotype. Int. J. Mol. Sci. 2019, 20, 1187. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Contreras, C.; Vazquez-Gomez, M.; Pardo, Z.; Heras-Molina, A.; Pesantez, J.L.; Encinas, T.; Torres-Rovira, L.; Astiz, S.; Nieto, R.; Ovilo, C.; et al. Polyphenols and IUGR pregnancies: Effects of maternal hydroxytyrosol supplementation on hepatic fat accretion and energy and fatty acids profile of fetal tissues. Nutrients 2019, 11, 1534. [Google Scholar] [CrossRef]
- Vazquez-Gomez, M.; Garcia-Contreras, C.; Torres-Rovira, L.; Pesantez, J.L.; Gonzalez-Añover, P.; Gomez-Fidalgo, E.; Sanchez-Sanchez, R.; Ovilo, C.; Isabel, B.; Astiz, S.; et al. Polyphenols and IUGR pregnancies: Maternal hydroxytyrosol supplementation improves prenatal and early-postnatal growth and metabolism of the offspring. PLoS ONE 2017, 12, e0177593. [Google Scholar] [CrossRef]
- Vazquez-Gomez, M.; Heras-Molina, A.; Garcia-Contreras, C.; Pesantez-Pacheco, J.L.; Torres-Rovira, L.; Martinez-Fernandez, B.; Gonzalez, J.; Encinas, T.; Astiz, S.; Ovilo, C.; et al. Polyphenols and IUGR Pregnancies: Effects of maternal hydroxytyrosol supplementation on postnatal growth, metabolism and body composition of the offspring. Antioxidants 2019, 8, 535. [Google Scholar] [CrossRef] [PubMed]
- Heras-Molina, A.; Pesántez-Pacheco, J.L.; Garcia-Contreras, C.; Vázquez-Gómez, M.; López, A.; Benítez, R.; Núñez, Y.; Astiz, S.; Óvilo, C.; Isabel, B.; et al. Maternal supplementation with polyphenols and omega-3 fatty acids during pregnancy: Prenatal effects on growth and metabolism. Animals 2021, 11, 1699. [Google Scholar] [CrossRef]
- Heras-Molina, A.; Pesantez-Pacheco, J.L.; Astiz, S.; Garcia-Contreras, C.; Vazquez-Gomez, M.; Encinas, T.; Óvilo, C.; Isabel, B.; Gonzalez-Bulnes, A. Maternal supplementation with polyphenols and omega-3 fatty acids during pregnancy: Effects on growth, metabolism, and body composition of the offspring. Animals 2020, 10, 1946. [Google Scholar] [CrossRef]
- National Research Council. Nutrient Requirements of Swine, 11th ed.; The National Academies Press: Washington, DC, USA, 2012. [Google Scholar] [CrossRef]
- Lopez-Bote, C.; Rey, A.; Ruiz, J.; Isabel, B.; Sanz Arias, R. Effect of feeding diets high in monounsaturated fatty acids and α-tocopheryl acetate to rabbits on resulting carcass fatty acid profile and lipid oxidation. Anim. Sci. 1997, 64, 177–186. [Google Scholar] [CrossRef]
- Sukhija, P.S.; Palmquist, D.L. Rapid method for determination of total fatty acid content and composition of feedstuffs and feces. J. Agric. Food Chem. 1988, 36, 1202–1206. [Google Scholar] [CrossRef]
- Segura, J.; Lopez-Bote, C.J. A laboratory efficient method for intramuscular fat analysis. Food Chem. 2014, 145, 821–825. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, J.; Antequera, T.; Andres, A.I.; Petron, M.J.; Muriel, E. Improvement of a solid phase extraction method for analysis of lipid fractions in muscle foods. Anal. Chim. Acta 2004, 520, 201–205. [Google Scholar] [CrossRef]
- Segura, J.; Escudero, R.; de Ávila, M.R.; Cambero, M.; López-Bote, C. Effect of fatty acid composition and positional distribution within the triglyceride on selected physical properties of dry-cured ham subcutaneous fat. Meat Sci. 2015, 103, 90–95. [Google Scholar] [CrossRef] [PubMed]
- Hulbert, A.J.; Pamplona, R.; Buffenstein, R.; Buttemer, W.A. Life and death: Metabolic rate, membrane composition, and life span of animals. Physiol. Rev. 2007, 87, 1175–1213. [Google Scholar] [CrossRef]
- Hulver, M.W.; Berggren, J.R.; Carper, M.J.; Miyazaki, M.; Ntambi, J.M.; Hoffman, E.P.; Thyfault, J.P.; Stevens, R.; Dohm, G.L.; Houmard, J.A. Elevated stearoyl-CoA desaturase-1 expression in skeletal muscle contributes to abnormal fatty acid partitioning in obese humans. Cell Metabol. 2005, 2, 251–261. [Google Scholar] [CrossRef] [PubMed]
- Tanghe, S.; Missotten, J.; Raes, K.; De Smet, S. The effect of different concentrations of linseed oil or fish oil in the maternal diet on the fatty acid composition and oxidative status of sows and piglets. J. Anim. Physiol. Anim. Nutr. 2015, 99, 938–949. [Google Scholar] [CrossRef]
- Gonzalez-Añover, P.; Encinas, T.; Torres-Rovira, L.; Pallares, P.; Muñoz-Frutos, J.; Gomez-Izquierdo, E.; Sanchez-Sanchez, R.; Gonzalez-Bulnes, A. Ovulation rate, embryo mortality and intrauterine growth retardation in obese swine with gene polymorphisms for leptin and melanocortin receptors. Theriogenology 2011, 75, 34–41. [Google Scholar] [CrossRef]
- Thomas, C.R.; Lowy, C. The interrelationships between circulating maternal esterified and non-esterified fatty acids in pregnant guinea pigs and their relative contributions to the fetal circulation. J. Dev. Physiol. 1987, 9, 203–214. [Google Scholar]
- Herrera, E.; Amusquivar, E.; López-Soldado, I.; Ortega, H. Maternal lipid metabolism and placental lipid transfer. Horm. Res. 2006, 65 (Suppl. S3), 59–64. [Google Scholar] [CrossRef]
- Kasser, T.R.; Martin, R.J.; Allen, C.E. Effect of gestational alloxan diabetes and fasting on fetal lipogenesis and lipid deposition in pigs. Neonatology 1981, 40, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Mersmann, H.J. Glycolytic and gluconeogenic enzyme levels in pre- and postnatal pigs. Am. J. Physiol. 1971, 220, 1297–1302. [Google Scholar] [CrossRef] [PubMed]
- Clandinin, M.T.; Chappell, J.E.; Heim, T.; Swyer, P.R.; Chance, G.W. Fatty acid utilization in perinatal de novo synthesis of tissues. Early Hum. Dev. 1981, 5, 355–366. [Google Scholar] [CrossRef]
- Schmitz, G.; Ecker, J. The opposing effects of n-3 and n-6 fatty acids. Prog. Lipid Res. 2008, 47, 147–155. [Google Scholar] [CrossRef]
- de Quelen, F.; Boudry, G.; Mourot, J. Linseed oil in the maternal diet increases long chain-PUFA status of the foetus and the newborn during the suckling period in pigs. Br. J. Nutr. 2010, 104, 533–543. [Google Scholar] [CrossRef]
- Tanghe, S.; Millet, S.; De Smet, S. Echium oil and linseed oil as alternatives for fish oil in the maternal diet: Blood fatty acid profiles and oxidative status of sows and piglets. J. Anim. Sci. 2013, 91, 3253–3264. [Google Scholar] [CrossRef]
- Bazinet, R.P.; McMillan, E.G.; Cunnane, S.C. Dietary alpha-linolenic acid increases the n-3 PUFA content of sow’s milk and the tissues of the suckling piglet. Lipids 2003, 38, 1045–1049. [Google Scholar] [CrossRef]
- Lugarà, R.; Realini, L.; Kreuzer, M.; Giller, K. Effects of maternal high-energy diet and spirulina supplementation in pregnant and lactating sows on performance, quality of carcass and meat, and its fatty acid profile in male and female offspring. Meat Sci. 2022, 187, 108769. [Google Scholar] [CrossRef]
- Barbero, A.; Astiz, S.; Lopez-Bote, C.J.; Perez-Solana, M.L.; Ayuso, M.; Garcia-Real, I.; Gonzalez-Bulnes, A. Maternal malnutrition and offspring sex determine juvenile obesity and metabolic disorders in a swine model of leptin resistance. PLoS ONE 2013, 8, e78424. [Google Scholar] [CrossRef]
- Decsi, T.; Kennedy, K. Sex-specific differences in essential fatty acid metabolism. Am. J. Clin. Nutr. 2011, 94, 1914s–1919s. [Google Scholar] [CrossRef] [PubMed]
- Childs, C.E. Sex hormones and n-3 fatty acid metabolism. Proc. Nutr. Soc. 2020, 79, 219–224. [Google Scholar] [CrossRef] [PubMed]
- Rey, A.I.; de-Cara, A.; Calvo, L.; Puig, P.; Hechavarría, T. Changes in plasma fatty acids, free amino acids, antioxidant defense, and physiological stress by oleuropein supplementation in pigs prior to slaughter. Antioxidants 2020, 9, 56. [Google Scholar] [CrossRef] [PubMed]
- Haugen, G.; Hanson, M.; Kiserud, T.; Crozier, S.; Inskip, H.; Godfrey, K.M. Fetal liver-sparing cardiovascular adaptations linked to mother’s slimness and diet. Circul. Res. 2005, 96, 12–14. [Google Scholar] [CrossRef]
- Pardo, C.; Bérard, J.; Kreuzer, M.; Bee, G. Intrauterine crowding impairs formation and growth of secondary myofibers in pigs. Animal 2013, 7, 430–438. [Google Scholar] [CrossRef]
- Aberle, E. Myofiber differentiation in skeletal muscles of newborn runt and normal weight pigs. J. Anim. Sci. 1984, 59, 1651–1656. [Google Scholar] [CrossRef]
- Alvarenga, A.; Chiarini-Garcia, H.; Cardeal, P.; Moreira, L.; Foxcroft, G.; Fontes, D.; Almeida, F. Intrauterine growth retardation affects birth-weight and postnatal development in pigs, impairing muscle accretion, duodenal mucosa morphology and carcass traits. Reprod. Fertil. Dev. 2013, 25, 387–395. [Google Scholar] [CrossRef]
- Fain, J.N.; Scow, R.O. Fatty acid synthesis in vivo in maternal and fetal tissues in the rat. Am. J. Physiol. Leg. Cont. 1966, 210, 19–25. [Google Scholar] [CrossRef]
- Vicente, J.G.; Isabel, B.; Cordero, G.; Lopez-Bote, C.J. Fatty acid profile of the sow diet alters fat metabolism and fatty acid composition in weanling pigs. Anim. Feed Sci. Technol. 2013, 181, 45–53. [Google Scholar] [CrossRef]
- Pan, D.A.; Hulbert, A.J.; Storlien, L.H. Dietary fats, membrane phospholipids and obesity. J. Nutr. 1994, 124, 1555–1565. [Google Scholar] [CrossRef]
- Liu, S.; Baracos, V.E.; Quinney, H.A.; Clandinin, M.T. Dietary omega-3 and polyunsaturated fatty acids modify fatty acyl composition and insulin binding in skeletal-muscle sarcolemma. Biochem. J. 1994, 299 Pt 3, 831–837. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fatty Acid (g/100 g Total FA) | Group C | Group T | Pooled S.E.M. | p-Value |
|---|---|---|---|---|
| C14:0 | 2.73 | 3.20 | 0.54 | 0.682 |
| C16:0 | 18.98 | 20.45 | 1.83 | 0.705 |
| C16:1n9 | 2.20 | 3.50 | 0.43 | 0.137 |
| C16:1n7 | 2.99 | 4.34 | 0.86 | 0.999 |
| C17:0 | 1.99 | 2.06 | 0.68 | 0.710 |
| C17:1 | 2.43 | 1.59 | 0.71 | 0.805 |
| C18:0 | 14.50 | 16.19 | 1.88 | 0.670 |
| C18:1n9 | 20.14 | 14.37 | 2.23 | 0.207 |
| C18:1n7 | 2.77 | 2.44 | 0.23 | 0.490 |
| C18:2n6 (LA) | 12.99 | 11.49 | 2.48 | 0.777 |
| C18:3n6 | 1.32 | 1.28 | 0.35 | 0.620 |
| C18:3n3 (ALA) | 1.67 | 2.25 | 0.37 | 0.805 |
| C18:4n3 | 1.12 | 1.50 | 0.25 | 0.209 |
| C20:1n9 | 2.45 | 2.82 | 0.55 | 0.747 |
| C20:3n6 | 1.00 | 1.15 | 0.20 | 0.730 |
| C20:4n6 (ARA) | 3.25 | 3.10 | 0.42 | 0.865 |
| C20:5n3 (EPA) | 1.22 | 2.07 | 0.34 | 0.227 |
| C22:4n6 | 2.76 | 2.54 | 0.43 | 0.815 |
| C22:5n3 (DPA) | 2.20 | 2.26 | 0.42 | 0.945 |
| C22:6n3 (DHA) | 1.30 | 1.38 | 0.16 | 0.806 |
| SFA | 38.20 | 41.91 | 3.02 | 0.561 |
| MUFA | 32.98 | 29.07 | 1.60 | 0.236 |
| PUFA | 28.83 | 29.03 | 2.54 | 0.969 |
| UI | 1.24 | 1.25 | 0.08 | 0.987 |
| MUFA/SFA | 0.86 | 0.69 | 0.08 | 0.302 |
| n3 | 7.51 | 9.47 | 1.20 | 0.437 |
| n6 | 21.32 | 19.56 | 2.27 | 0.716 |
| n6/n3 | 2.84 | 2.07 | 0.75 | 0.165 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heras-Molina, A.; Escudero, R.; Pesántez-Pacheco, J.L.; García-Contreras, C.; Vázquez-Gómez, M.; Astiz, S.; Óvilo, C.; González-Bulnes, A.; Isabel, B. Maternal Supplementation with Polyphenols and Omega-3 Fatty Acids during Pregnancy: Prenatal Effects on Fetal Fatty Acid Composition in the Iberian Pig. Animals 2022, 12, 2140. https://doi.org/10.3390/ani12162140
Heras-Molina A, Escudero R, Pesántez-Pacheco JL, García-Contreras C, Vázquez-Gómez M, Astiz S, Óvilo C, González-Bulnes A, Isabel B. Maternal Supplementation with Polyphenols and Omega-3 Fatty Acids during Pregnancy: Prenatal Effects on Fetal Fatty Acid Composition in the Iberian Pig. Animals. 2022; 12(16):2140. https://doi.org/10.3390/ani12162140
Chicago/Turabian StyleHeras-Molina, Ana, Rosa Escudero, José L. Pesántez-Pacheco, Consolación García-Contreras, Marta Vázquez-Gómez, Susana Astiz, Cristina Óvilo, Antonio González-Bulnes, and Beatriz Isabel. 2022. "Maternal Supplementation with Polyphenols and Omega-3 Fatty Acids during Pregnancy: Prenatal Effects on Fetal Fatty Acid Composition in the Iberian Pig" Animals 12, no. 16: 2140. https://doi.org/10.3390/ani12162140