
Evidence-Based Approaches for Determining Effective Target Antigens to Develop Vaccines against Post-Weaning Diarrhea Caused by Enterotoxigenic Escherichia coli in Pigs: A Systematic Review and Network Meta-Analysis


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Research Question and Search Strategy
2.2. Eligibility and Exclusion Criteria
2.3. Data Extraction
2.4. Risk of Bias Assessment
2.5. Statistical Analyses of the Data
2.5.1. Pairwise MA
2.5.2. Bayesian NMA
3. Results
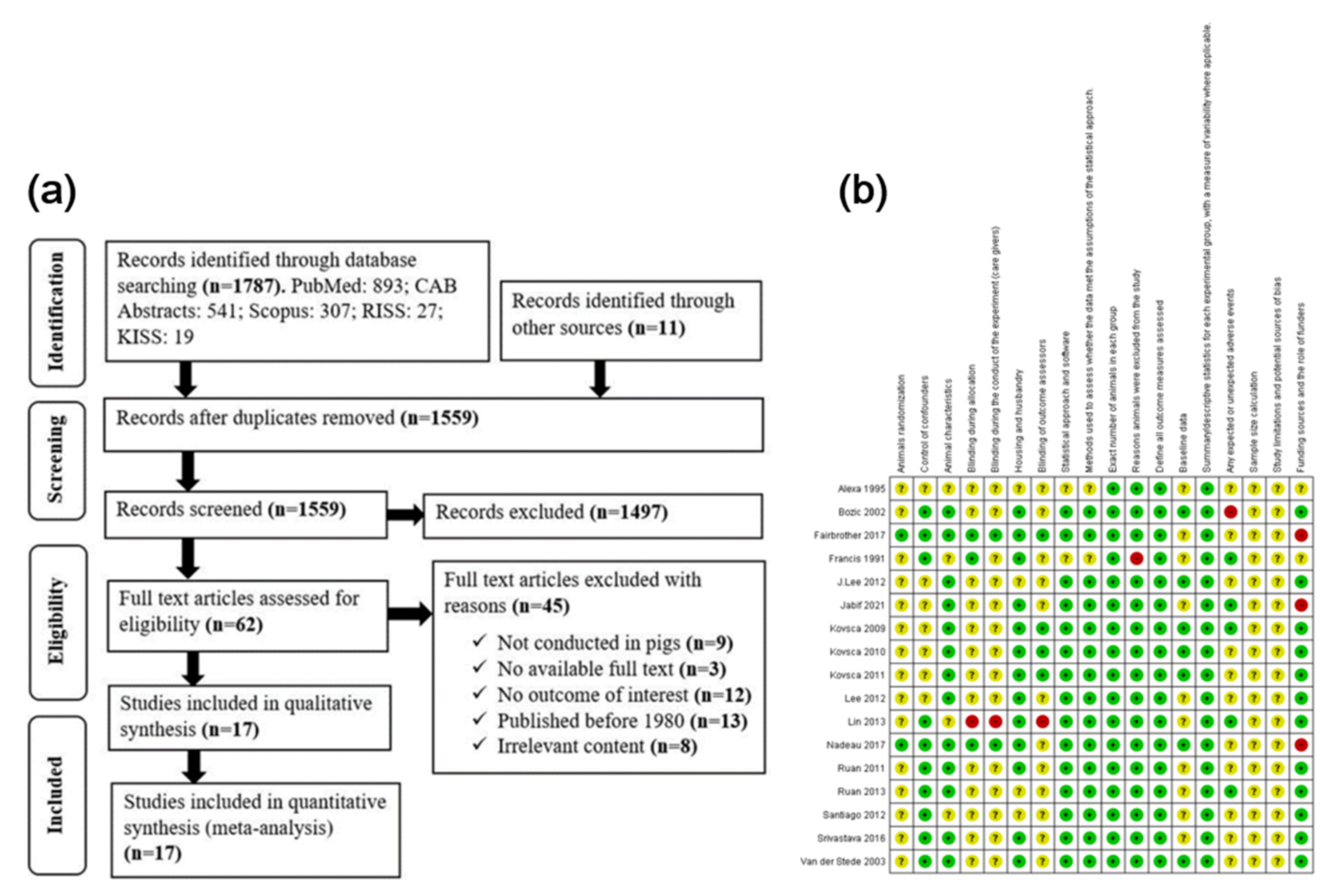
3.1. Search Results
3.2. Study Characteristics
3.3. Quality Assessment
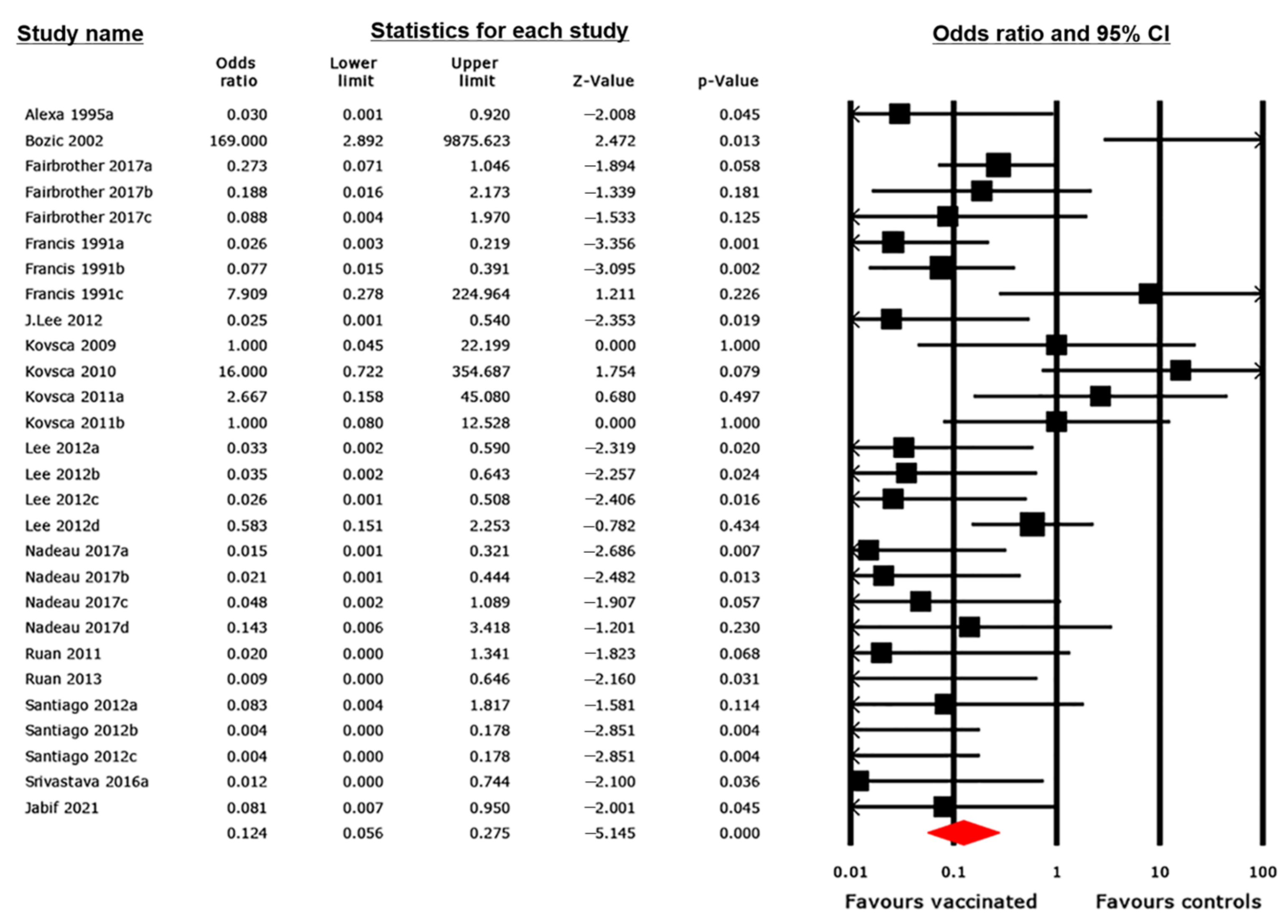
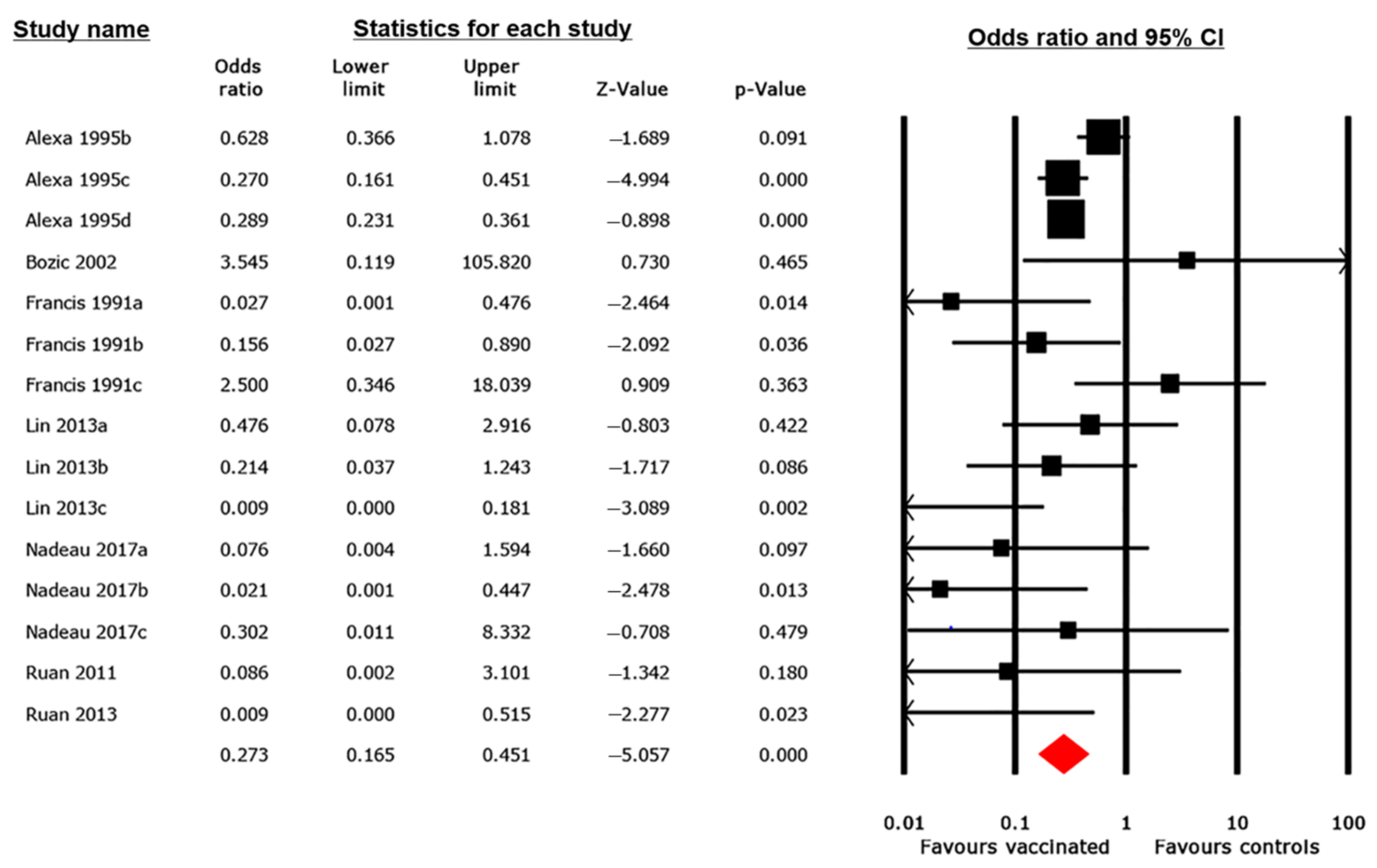
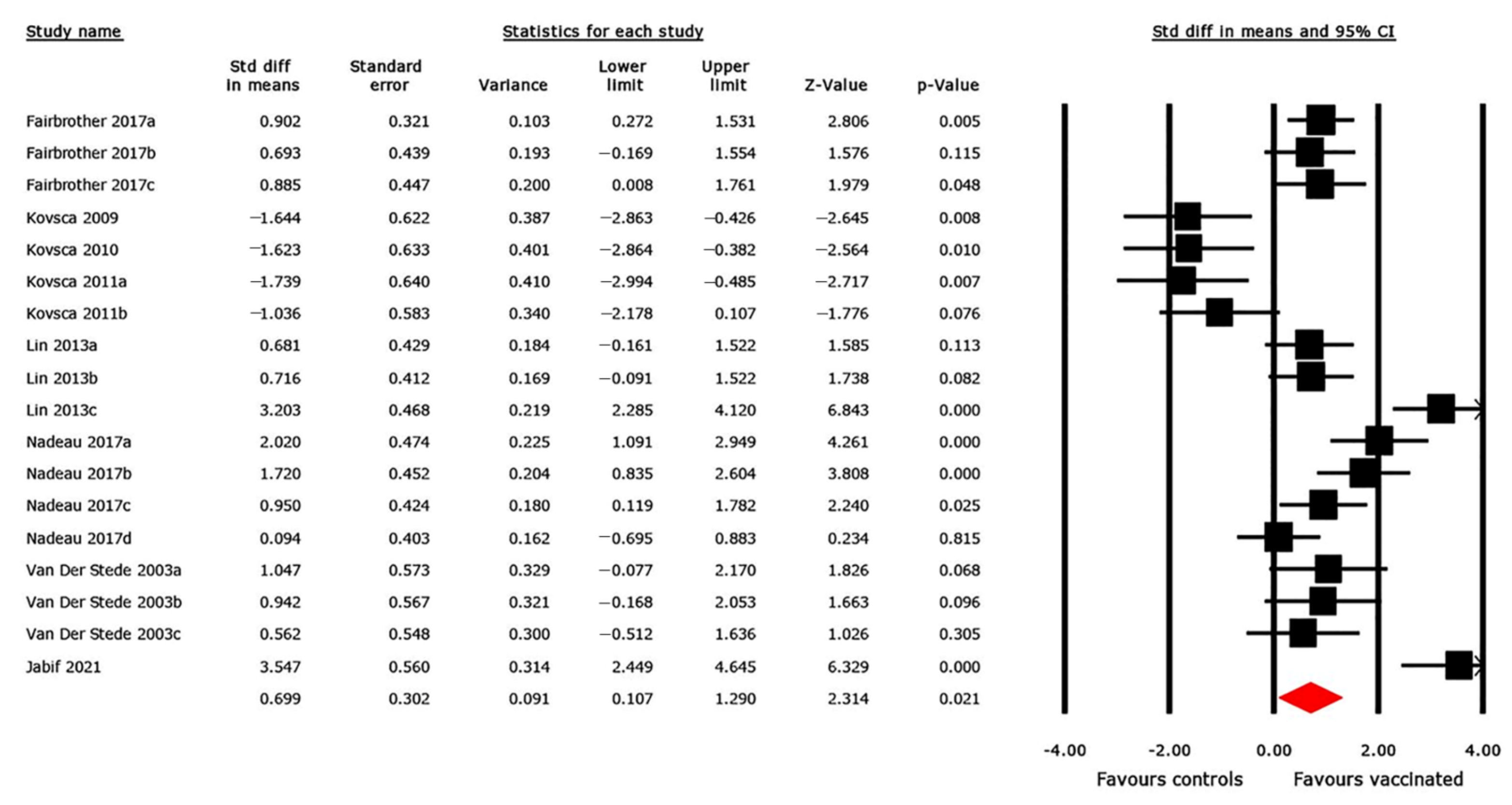
3.4. Pairwise MA Results
3.4.1. Diarrhea Outcome
3.4.2. Mortality Outcome
3.4.3. ADWG Outcome
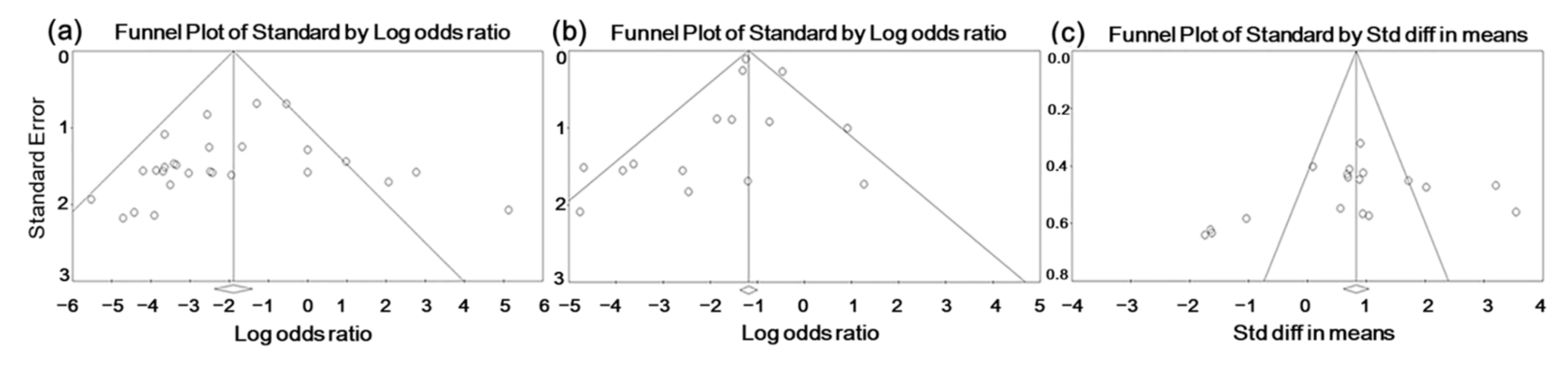
3.4.4. Publication Bias
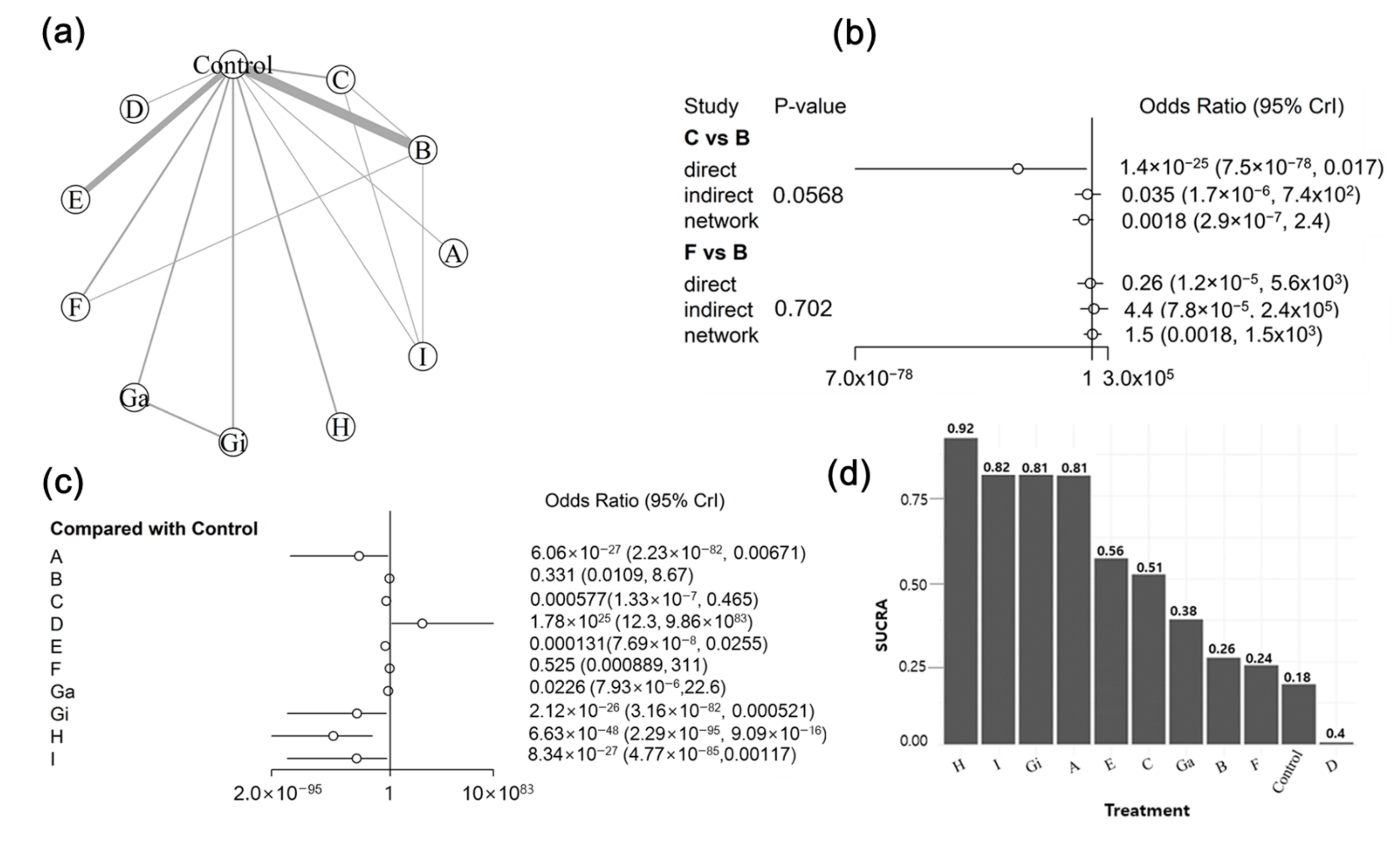
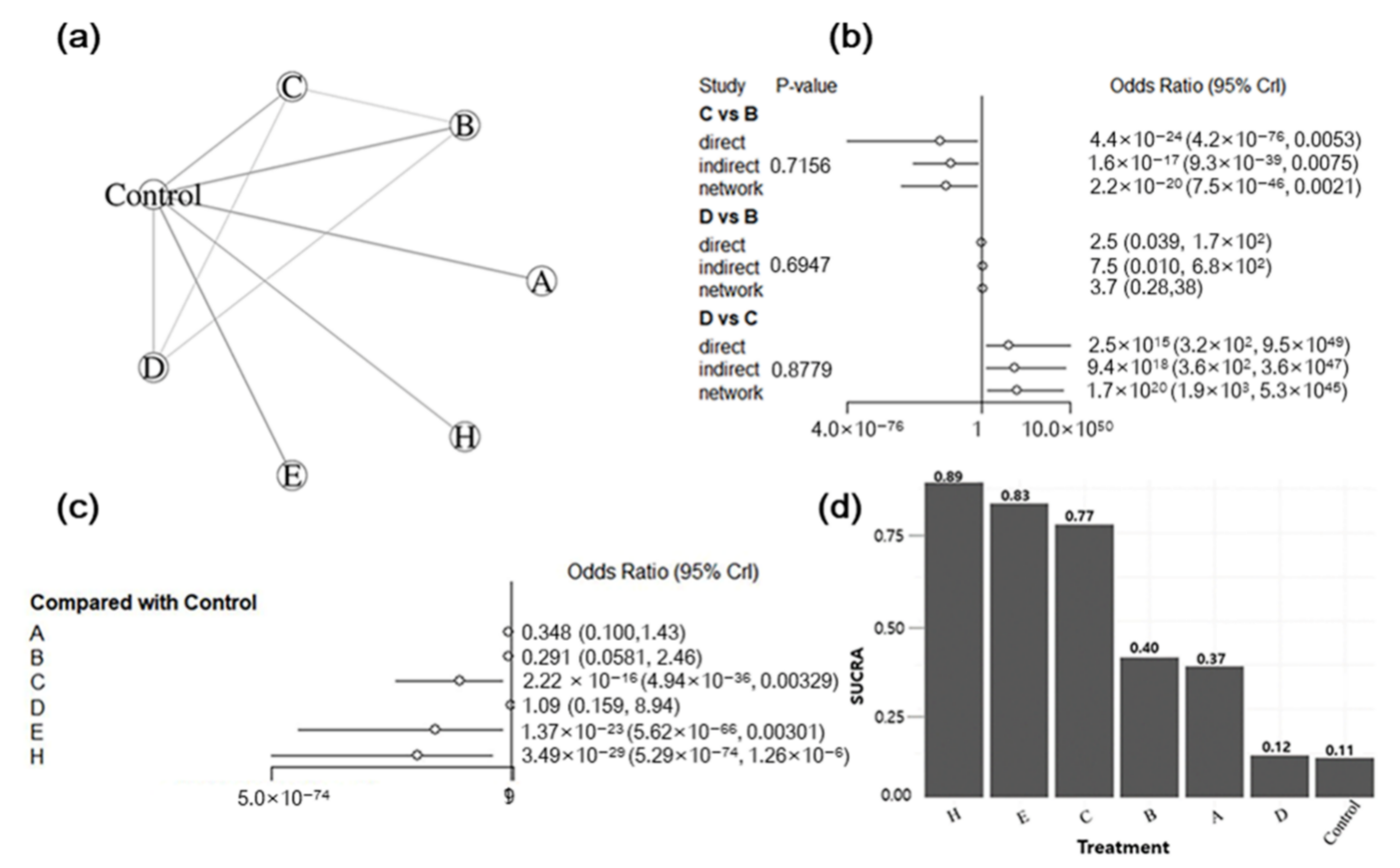
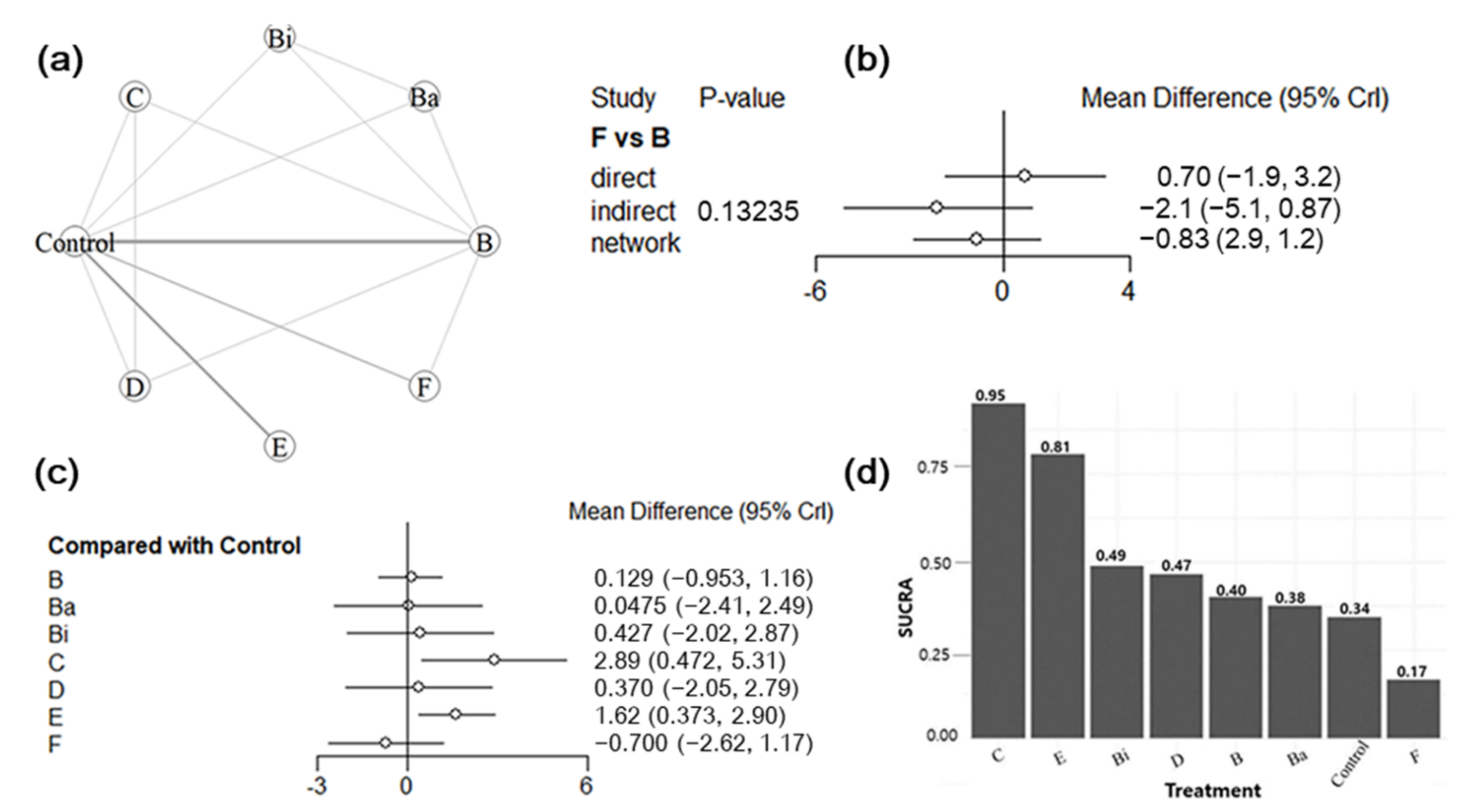
3.5. Bayesian NMA
3.5.1. Summary of the Network Geometry
3.5.2. Assessment of Inconsistency
3.5.3. Synthesis of NMA Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hartadi, E.B.; Effendi, M.H.; Plumeriastuti, H.; Sofiana, E.D.; Wibisono, F.M.; Hidayatullah, A.R. A Review of Enterotoxigenic Escherichia coli Infection in Piglets: Public Health Importance. Syst. Rev. Pharm. 2020, 11, 687–698. [Google Scholar]
- Alexa, P.; Hamrik, J.; Stouracova, K.; Salajka, E. Oral immunization against enterotoxigenic colibacillosis in weaned piglets by non-pathogenic Escherichia coli strain with K88 (F4) colonizing factors. Veterinární Med. 2005, 50, 315–320. [Google Scholar] [CrossRef]
- Cox, E.; Melkebeek, V.; Devriendt, B.; Goddeeris, B.; Vanrompay, D. Vaccines against enteric E. coli infections in animals. In Pathogenic Escherichia coli: Molecular and Cellular Microbiology; Caister Academic Press: Norfolk, UK, 2014; pp. 255–270. [Google Scholar]
- Nadeau, É.; Fairbrother, J.M.; Zentek, J.; Bélanger, L.; Tremblay, D.; Tremblay, C.L.; Röhe, I.; Vahjen, W.; Brunelle, M.; Hellmann, K.; et al. Efficacy of a single oral dose of a live bivalent E. coli vaccine against post-weaning diarrhea due to F4 and F18-positive enterotoxigenic E. coli. Vet. J. 2017, 226, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Fairbrother, J.M.; Nadeau, É.; Gyles, C.L. Escherichia coli in postweaning diarrhea in pigs: An update on bacterial types, pathogenesis, and prevention strategies. Anim. Health Res. Rev. 2005, 6, 17. [Google Scholar] [CrossRef]
- Luppi, A. Swine enteric colibacillosis: Diagnosis, therapy and antimicrobial resistance. Porcine Health Manag. 2017, 3, 16. [Google Scholar] [CrossRef]
- Janjatović, A.K.; Lacković, G.; Božić, F.; Špoljarić, D.; Popović, M.; Valpotić, H.; Vijtiuk, N.; Pavičić, Ž.; Valpotić, I. Histomorphometric characteristics of immune cells in small intestine of pigs perorally immunized with vaccine candidate F18ac+ nonenterotoxigenic E. coli strain. Eur. J. Histochem. : EJH 2009, 53, e23. [Google Scholar] [CrossRef]
- Nagy, B.; Fekete, P.Z. Enterotoxigenic Escherichia coli in veterinary medicine. Int. J. Med. Microbiol. 2005, 295, 443–454. [Google Scholar] [CrossRef]
- Fairbrother, J.M.; Nadeau, É.; Bélanger, L.; Tremblay, C.L.; Tremblay, D.; Brunelle, M.; Wolf, R.; Hellmann, K.; Hidalgo, Á. Immunogenicity and protective efficacy of a single-dose live non-pathogenic Escherichia coli oral vaccine against F4-positive enterotoxigenic Escherichia coli challenge in pigs. Vaccine 2017, 35, 353–360. [Google Scholar] [CrossRef]
- Meijerink, E.; Neuenschwander, S.; Fries, R.; Dinter, A.; Bertschinger, H.U.; Stranzinger, G.; Vögeli, P. A DNA polymorphism influencing α (1, 2) fucosyltransferase activity of the pig FUT1 enzyme determines susceptibility of small intestinal epithelium to Escherichia coli F18 adhesion. Immunogenetics 2000, 52, 129–136. [Google Scholar] [CrossRef]
- Liu, L.; Wang, J.; Zhao, Q.; Zi, C.; Wu, Z.; Su, X.; Huo, Y.; Zhu, G.; Wu, S.; Bao, W. Genetic variation in exon 10 of the BPI gene is associated with Escherichia coli F18 susceptibility in Sutai piglets. Gene 2013, 523, 70–75. [Google Scholar] [CrossRef]
- Vögeli, P.; Bertschinger, H.U.; Stamm, M.; Stricker, C.; Hagger, C.; Fries, R.; Rapacz, J.; Stranzinger, G. Genes specifying receptors for F18 fimbriated Escherichia coli, causing oedema disease and postweaning diarrhoea in pigs, map to chromosome 6. Anim. Genet. 1996, 27, 321–328. [Google Scholar] [PubMed]
- Ren, J.; Yan, X.; Ai, H.; Zhang, Z.; Huang, X.; Ouyang, J.; Yang, M.; Yang, H.; Han, P.; Zeng, W. Susceptibility towards enterotoxigenic Escherichia coli F4ac diarrhea is governed by the MUC13 gene in pigs. PLoS ONE 2012, e44573. [Google Scholar] [CrossRef] [PubMed]
- Ji, H.Y.; Yang, B.; Zhang, Z.Y.; Ouyang, J.; Yang, M.; Zhang, X.F.; Zhang, W.C.; Su, Y.; Zhao, K.W.; Xiao, S.J.; et al. A genome-wide association analysis for susceptibility of pigs to enterotoxigenic Escherichia coli F41. Animal 2016, 10, 1602–1608. [Google Scholar] [CrossRef] [PubMed]
- Luise, D.; Lauridsen, C.; Bosi, P.; Trevisi, P. Methodology and application of Escherichia coli F4 and F18 encoding infection models in post-weaning pigs. J. Anim. Sci. Biotechnol. 2019, 10, 53. [Google Scholar] [CrossRef]
- Zhao, Q.; Liu, Y.; Dong, W.; Zhu, S.; Huo, Y.; Wu, S.; Bao, W. Genetic variations of TAP1 gene exon 3 affects gene expression and Escherichia coli F18 resistance in piglets. Int. J. Mol. Sci. 2014, 15, 11161–11171. [Google Scholar] [CrossRef]
- Bogere, P.; Choi, Y.; Heo, J. Probiotics as alternatives to antibiotics in treating post-weaning diarrhoea in pigs. South Afr. J. Anim. Sci. 2019, 49, 403–416. [Google Scholar] [CrossRef]
- Van den Broeck, W.; Cox, E.; Goddeeris, B.M. Receptor-dependent immune responses in pigs after oral immunization with F4 fimbriae. Infect. Immun. 1999, 67, 520–526. [Google Scholar] [CrossRef]
- Melkebeek, V.; Goddeeris, B.M.; Cox, E. ETEC vaccination in pigs. Vet. Immunol. Immunopathol. 2013, 152, 37–42. [Google Scholar] [CrossRef]
- O’brien, A.; Holmes, R. Protein toxins of Escherichia coli and Salmonella. Escherichia Coli Salmonella: Cell. Mol. Biol. 1996, 2, 2788–2802. [Google Scholar]
- Duan, Q.; Xia, P.; Nandre, R.; Zhang, W.; Zhu, G. Review of Newly Identified Functions Associated With the Heat-Labile Toxin of Enterotoxigenic Escherichia coli. Front. Cell. Infect. Microbiol. 2019, 9, 292. [Google Scholar] [CrossRef]
- Rhouma, M.; Fairbrother, J.M.; Beaudry, F.; Letellier, A. Post weaning diarrhea in pigs: Risk factors and non-colistin-based control strategies. Acta Vet. Scand. 2017, 59, 31. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W. Progress and Challenges in Vaccine development Against Enterotoxigenic Escherichia Coli (ETEC)-Associated porcine PostWeaning Diarrhoea (PWD). J. Vet. Med. Res. 2014, 1. [Google Scholar]
- Haidich, A.B. Meta-analysis in medical research. Hippokratia 2010, 14, 29–37. [Google Scholar] [PubMed]
- Le Boedec, K. A systematic review and meta-analysis of the association between Mycoplasma spp and upper and lower respiratory tract disease in cats. J. Am. Vet. Med. Assoc. 2017, 250, 397–407. [Google Scholar] [CrossRef]
- Lean, I.J.; Rabiee, A.R.; Duffield, T.F.; Dohoo, I.R. Invited review: Use of meta-analysis in animal health and reproduction: Methods and applications. J. Dairy Sci. 2009, 92, 3545–3565. [Google Scholar] [CrossRef]
- Andy, P. Field, R.G. How to do a meta-analysis. Br. J. Math. Stat. Psychol. 2010, 63, 665–694. [Google Scholar] [CrossRef]
- O’Connor, A.M.; Hu, D.; Totton, S.C.; Scott, N.; Winder, C.B.; Wang, B.; Wang, C.; Glanville, J.; Wood, H.; White, B.; et al. A systematic review and network meta-analysis of bacterial and viral vaccines, administered at or near arrival at the feedlot, for control of bovine respiratory disease in beef cattle. Anim. Health Res. Rev. 2019, 20, 143–162. [Google Scholar] [CrossRef]
- Mills, E.J.; Ioannidis, J.P.; Thorlund, K.; Schünemann, H.J.; Puhan, M.A.; Guyatt, G.H. How to use an article reporting a multiple treatment comparison meta-analysis. JAMA 2012, 308, 1246–1253. [Google Scholar] [CrossRef]
- Rouse, B.; Chaimani, A.; Li, T. Network meta-analysis: An introduction for clinicians. Intern. Emerg. Med. 2017, 12, 103–111. [Google Scholar] [CrossRef]
- Higgins, J.P.; Thomas, J.; Chandler, J.; Cumpston, M.; Li, T.; Page, M.J.; Welch, V.A. Cochrane Handbook for Systematic Reviews of Interventions; John Wiley & Sons: Hoboken, NJ, USA, 2019. [Google Scholar]
- Yogiswara, N.; Azmi, Y.; Azmi, Y.A. A network meta-analysis on comparative efficacy and tolerability of on-demand therapy for premature ejaculation. Bali Med. J. 2020, 9, 591–598. [Google Scholar] [CrossRef]
- Watt, J.A.; Tricco, A.C.; Straus, S.E.; Veroniki, A.A.; Naglie, G.; Drucker, A.M. Research Techniques Made Simple: Network Meta-Analysis. J. Investig. Dermatol. 2019, 139, 4–12.e11. [Google Scholar] [CrossRef] [PubMed]
- Shim, S.R.; Kim, S.-J.; Lee, J.; Rücker, G. Network meta-analysis: Application and practice using R software. Epidemiol. Health 2019, 41, e2019013. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-I.; Ntakiyisumba, E.; Won, G. Systematic review and network meta-analysis to compare vaccine effectiveness against porcine edema disease caused by Shiga toxin-producing Escherichia coli. Sci. Rep. 2022, 12, 6460. [Google Scholar] [CrossRef] [PubMed]
- Calzetta, L.; Pistocchini, E.; Leo, A.; Roncada, P.; Ritondo, B.L.; Palma, E.; di Cave, D.; Britti, D. Anthelminthic medicinal plants in veterinary ethnopharmacology: A network meta-analysis following the PRISMA-P and PROSPERO recommendations. Heliyon 2020, 6, e03256. [Google Scholar] [CrossRef] [PubMed]
- Page, M.J.; Moher, D.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. PRISMA 2020 explanation and elaboration: Updated guidance and exemplars for reporting systematic reviews. BMJ 2021, 372, n160. [Google Scholar] [CrossRef]
- Hutton, B.; Salanti, G.; Caldwell, D.M.; Chaimani, A.; Schmid, C.H.; Cameron, C.; Ioannidis, J.P.; Straus, S.; Thorlund, K.; Jansen, J.P.; et al. The PRISMA extension statement for reporting of systematic reviews incorporating network meta-analyses of health care interventions: Checklist and explanations. Ann. Intern. Med. 2015, 162, 777–784. [Google Scholar] [CrossRef]
- Drevon, D.; Fursa, S.R.; Malcolm, A.L. Intercoder Reliability and Validity of WebPlotDigitizer in Extracting Graphed Data. Behav. Modif. 2017, 41, 323–339. [Google Scholar] [CrossRef]
- Percie du Sert, N.; Hurst, V.; Ahluwalia, A.; Alam, S.; Avey, M.T.; Baker, M.; Browne, W.J.; Clark, A.; Cuthill, I.C.; Dirnagl, U.; et al. The ARRIVE guidelines 2.0: Updated guidelines for reporting animal research. J. Cereb. Blood Flow Metab. 2020, 40, 1769–1777. [Google Scholar] [CrossRef]
- Higgins, J.P.; Thompson, S.G.; Deeks, J.J.; Altman, D.G. Measuring inconsistency in meta-analyses. BMJ 2003, 327, 557–560. [Google Scholar] [CrossRef]
- Sabarimurugan, S.; Kumarasamy, C.; Baxi, S.; Devi, A.; Jayaraj, R. Systematic review and meta-analysis of prognostic microRNA biomarkers for survival outcome in nasopharyngeal carcinoma. PLoS ONE 2019, 14, e0209760. [Google Scholar] [CrossRef]
- Duval, S.; Tweedie, R. Trim and fill: A simple funnel-plot–based method of testing and adjusting for publication bias in meta-analysis. Biometrics 2000, 56, 455–463. [Google Scholar] [CrossRef] [PubMed]
- Salanti, G.; Ades, A.E.; Ioannidis, J.P.A. Graphical methods and numerical summaries for presenting results from multiple-treatment meta-analysis: An overview and tutorial. J. Clin. Epidemiol. 2011, 64, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Plummer, M.; Stukalov, A.; Denwood, M. rjags: Bayesian graphical models using MCMC. Available online: https://CRAN.R-project.org/package=rjags (accessed on 12 August 2022).
- Brooks, S.P.; Gelman, A. General Methods for Monitoring Convergence of Iterative Simulations. J. Comput. Graph. Stat. 1998, 7, 434–455. [Google Scholar] [CrossRef]
- Faltinsen, E.G.; Storebø, O.J.; Jakobsen, J.C.; Boesen, K.; Lange, T.; Gluud, C. Network meta-analysis: The highest level of medical evidence? BMJ Evid. Based Med. 2018, 23, 56–59. [Google Scholar] [CrossRef]
- Higgins, J.; Jackson, D.; Barrett, J.; Lu, G.; Ades, A.; White, I. Consistency and inconsistency in network meta-analysis: Concepts and models for multi-arm studies. Res. Synth. Methods 2012, 3, 98–110. [Google Scholar] [CrossRef] [PubMed]
- Van Valkenhoef, G.; Dias, S.; Ades, A.E.; Welton, N.J. Automated generation of node-splitting models for assessment of inconsistency in network meta-analysis. Res. Synth. Methods 2016, 7, 80–93. [Google Scholar] [CrossRef]
- Schwarzer, G.; Carpenter, J.R.; Rücker, G. Meta-Analysis with R.; Springer: Berlin/Heidelberg, Germany, 2015; Volume 4784. [Google Scholar]
- Hur, J.; Lee, J.H. Comparative evaluation of a vaccine candidate expressing enterotoxigenic Escherichia coli (ETEC) adhesins for colibacillosis with a commercial vaccine using a pig model. Vaccine 2012, 30, 3829–3833. [Google Scholar] [CrossRef]
- Francis, D.H.; Willgohs, J.A. Evaluation of a live avirulent Escherichia coli vaccine for K88+, LT+ enterotoxigenic colibacillosis in weaned pigs. Am. J. Vet. Res. 1991, 52, 1051–1055. [Google Scholar]
- Alexa, P.; Salajka, E.; Salajková, Z.; Máchová, A. [Combined parenteral and oral immunization against enterotoxigenic Escherichia coli diarrhea in weaned piglets]. Vet. Med. 1995, 40, 365–370. [Google Scholar]
- Bozić, F.; Lacković, G.; Stokes, C.R.; Valpotić, I. Recruitment of intestinal CD45RA+ and CD45RC+ cells induced by a candidate oral vaccine against porcine post-weaning colibacillosis. Vet. Immunol. Immunopathol. 2002, 86, 137–146. [Google Scholar] [CrossRef]
- Van der Stede, Y.; Cox, E.; Verdonck, F.; Vancaeneghem, S.; Goddeeris, B.M. Reduced faecal excretion of F4+-E coli by the intramuscular immunisation of suckling piglets by the addition of 1alpha,25-dihydroxyvitamin D3 or CpG-oligodeoxynucleotides. Vaccine 2003, 21, 1023–1032. [Google Scholar] [CrossRef]
- Janjatović, A.K.; Lacković, G.; Božić, F.; Valpotić, H.; Tominac-Trcin, M.; Sladoljev, S.; Šeol, B.; Valpotić, I.; Popović, M. De novo differentiation of intestinal villous M cells in weaned pigs immunized with levamisole-adjuvanted vaccine candidate F4ac+ or F18ac+ non-enterotoxigenic Escherichia coli strains. Vet. Arhiv. 2011, 81, 77–90. [Google Scholar]
- Ruan, X.; Liu, M.; Casey, T.A.; Zhang, W. A tripartite fusion, FaeG-FedF-LT(192)A2:B, of enterotoxigenic Escherichia coli (ETEC) elicits antibodies that neutralize cholera toxin, inhibit adherence of K88 (F4) and F18 fimbriae, and protect pigs against K88ac/heat-labile toxin infection. Clin. Vaccine Immunol. 2011, 18, 1593–1599. [Google Scholar] [CrossRef] [PubMed]
- Ruan, X.; Zhang, W. Oral immunization of a live attenuated Escherichia coli strain expressing a holotoxin-structured adhesin-toxoid fusion (1FaeG-FedF-LTA₂:5LTB) protected young pigs against enterotoxigenic E. coli (ETEC) infection. Vaccine 2013, 31, 1458–1463. [Google Scholar] [CrossRef]
- Hur, J.; Lee, J.H. Development of a novel live vaccine delivering enterotoxigenic Escherichia coli fimbrial antigens to prevent post-weaning diarrhea in piglets. Vet. Immunol. Immunopathol. 2012, 146, 283–288. [Google Scholar] [CrossRef]
- Santiago-Mateo, K.; Zhao, M.; Lin, J.; Zhang, W.; Francis, D.H. Avirulent K88 (F4)+ Escherichia coli strains constructed to express modified enterotoxins protect young piglets from challenge with a virulent enterotoxigenic Escherichia coli strain that expresses the same adhesion and enterotoxins. Vet. Microbiol. 2012, 159, 337–342. [Google Scholar] [CrossRef]
- Lin, J.; Mateo, K.S.; Zhao, M.; Erickson, A.K.; Garcia, N.; He, D.; Moxley, R.A.; Francis, D.H. Protection of piglets against enteric colibacillosis by intranasal immunization with K88ac (F4ac) fimbriae and heat labile enterotoxin of Escherichia coli. Vet. Microbiol. 2013, 162, 731–739. [Google Scholar] [CrossRef]
- Srivastava, A.; Gowda, D.V.; Madhunapantula, S.V.; Siddaramaiah. Development and Efficacy Assessment of an Enteric Coated Porous Tablet Loaded With F4 Fimbriae for Oral Vaccination of Piglets against F4+ Escherichia coli Infections. Curr. Drug Deliv. 2016, 13, 121–130. [Google Scholar] [CrossRef]
- Janjatović, A.K.; Lacković, G.; Božić, F.; Kezić, D.; Popović, M.; Valpotić, H.; Harapin, I.; Pavižić, Ž.; Njari, B.; Valpotić, I. Histomorphometric evaluation of intestinal cellular immune responses in pigs immunized with live oral F4ac+ non-enterotoxigenic E. coli vaccine against postweaning colibacillosis. Eur. J. Histochem. EJH 2010, 54, e4. [Google Scholar] [CrossRef]
- Jabif, M.F.; Gumina, E.; Hall, J.W.; Hernandez-Velasco, X.; Layton, S. Evaluation of a Novel Mucosal Administered Subunit Vaccine on Colostrum IgA and Serum IgG in Sows and Control of Enterotoxigenic Escherichia coli in Neonatal and Weanling Piglets: Proof of Concept. Front. Vet. Sci. 2021, 8, 640228. [Google Scholar] [CrossRef]
- Ou, B.; Jiang, B.; Jin, D.; Yang, Y.; Zhang, M.; Zhang, D.; Zhao, H.; Xu, M.; Song, H.; Wu, W.; et al. Engineered Recombinant Escherichia coli Probiotic Strains Integrated with F4 and F18 Fimbriae Cluster Genes in the Chromosome and Their Assessment of Immunogenic Efficacy in Vivo. ACS Synth. Biol. 2020, 9, 412–426. [Google Scholar] [CrossRef] [PubMed]
- Sack, R.; Kline, R.; Spira, W. Oral immunization of rabbits with enterotoxigenic Escherichia coli protects against intraintestinal challenge. Infect. Immun. 1988, 56, 387–394. [Google Scholar] [CrossRef] [PubMed]
- Vesikari, T.; Isolauri, E.; Delem, A.; D’Hondt, E.; André, F.; Zissis, G. Immunogenicity and safety of live oral attenuated bovine rotavirus vaccine strain RIT 4237 in adults and young children. Lancet 1983, 322, 807–811. [Google Scholar] [CrossRef]
- Duan, Q.; Pang, S.; Wu, W.; Jiang, B.; Zhang, W.; Liu, S.; Wang, X.; Pan, Z.; Zhu, G. A multivalent vaccine candidate targeting enterotoxigenic Escherichia coli fimbriae for broadly protecting against porcine post-weaning diarrhea. Vet. Res. 2020, 51, 93. [Google Scholar] [CrossRef] [PubMed]
- Hur, J.; Stein, B.D.; Lee, J.H. A vaccine candidate for post-weaning diarrhea in swine constructed with a live attenuated Salmonella delivering Escherichia coli K88ab, K88ac, FedA, and FedF fimbrial antigens and its immune responses in a murine model. Can. J. Vet. Res. 2012, 76, 186–194. [Google Scholar]
- Gyles, C.L.; Maas, W.K. Recombinant DNA technology and enterotoxigenic Escherichia coli vaccines. Prog. Vet. Microbiol. Immunol. 1987, 3, 139–158. [Google Scholar]
- Johnson, T.J.; Nolan, L.K. Pathogenomics of the virulence plasmids of Escherichia coli. Microbiol. Mol. Biol. Rev. 2009, 73, 750. [Google Scholar] [CrossRef]
- Zhang, W.; Berberov, E.M.; Freeling, J.; He, D.; Moxley, R.A.; Francis, D.H. Significance of heat-stable and heat-labile enterotoxins in porcine colibacillosis in an additive model for pathogenicity studies. Infect. Immun. 2006, 74, 3107–3114. [Google Scholar] [CrossRef]
- Schlager, T.A.; Wanke, C.A.; Guerrant, R.L. Net fluid secretion and impaired villous function induced by colonization of the small intestine by nontoxigenic colonizing Escherichia coli. Infect. Immun. 1990, 58, 1337–1343. [Google Scholar] [CrossRef]
- Smith, H.W.; Linggood, M.A. Observations on the pathogenic properties of the K88, Hly and Ent plasmids of Escherichia coli with particular reference to porcine diarrhoea. J. Med. Microbiol. 1971, 4, 467–485. [Google Scholar] [CrossRef]
- Bakker, D.; Willemsen, P.; Willems, R.; Huisman, T.T.; Mooi, F.; Oudega, B.; Stegehuis, F.; De Graaf, F. Identification of minor fimbrial subunits involved in biosynthesis of K88 fimbriae. J. Bacteriol. 1992, 174, 6350–6358. [Google Scholar] [CrossRef] [PubMed]
- Dubreuil, J.D. Pig vaccination strategies based on enterotoxigenic Escherichia coli toxins. Braz. J. Microbiol. 2021, 52, 2499–2509. [Google Scholar] [CrossRef] [PubMed]
- Smeds, A.; Hemmann, K.; Jakava-Viljanen, M.; Pelkonen, S.; Imberechts, H.; Palva, A. Characterization of the adhesin of Escherichia coli F18 fimbriae. Infect. Immun. 2001, 69, 7941–7945. [Google Scholar] [CrossRef] [PubMed]
- Clements, J.D.; Norton, E.B. The Mucosal Vaccine Adjuvant LT(R192G/L211A) or dmLT. mSphere 2018, 3, e00215–e00218. [Google Scholar] [CrossRef] [PubMed]
- Verdonck, F.; Tiels, P.; van Gog, K.; Goddeeris, B.M.; Lycke, N.; Clements, J.; Cox, E. Mucosal immunization of piglets with purified F18 fimbriae does not protect against F18+ Escherichia coli infection. Vet. Immunol. Immunopathol. 2007, 120, 69–79. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Number of Studies | Effect Size and 95% Confidence Interval | Test of Null (Two-Tail) | Heterogeneity | |||||
|---|---|---|---|---|---|---|---|---|---|
| Point Estimate | Lower Limit | Upper Limit | Z-Value | p-Value | Q-Value | df (Q) | p-Value | ||
| F18 + LT + ST | 1 | 0.030 | 0.000 | 1.896 | −1.658 | 0.097 | 19.162 | 8 | 0.014 |
| F18 | 2 | 1.000 | 0.076 | 13.160 | 0.000 | 1.000 | |||
| F4 | 9 | 0.388 | 0.118 | 1.270 | −1.565 | 0.117 | |||
| F4 + F18 | 6 | 0.041 | 0.009 | 0.194 | −4.040 | 0.000 | |||
| F4 + F18 + LT | 2 | 0.013 | 0.000 | 0.414 | −2.465 | 0.014 | |||
| F4 + F5 + F6 + F41 | 4 | 0.098 | 0.018 | 0.528 | −2.701 | 0.007 | |||
| F4 + LT | 2 | 0.014 | 0.001 | 0.183 | −3.248 | 0.001 | |||
| F4 + LT + ST | 1 | 0.004 | 0.000 | 0.346 | −2.427 | 0.015 | |||
| LT | 1 | 7.909 | 0.133 | 469.971 | 0.992 | 0.321 | |||
| Overall | 28 | 0.128 | 0.063 | 0.257 | −5.758 | 0.000 | |||
| Groups | Number of Studies | Effect Size and 95% Confidence Interval | Test of Null (Two-Tail) | Heterogeneity | |||||
|---|---|---|---|---|---|---|---|---|---|
| Point Estimate | Lower Limit | Upper Limit | Z-Value | p-Value | Q-Value | df (Q) | p-Value | ||
| F18 + LT + ST | 3 | 0.351 | 0.229 | 0.536 | −4.835 | 0.000 | |||
| F4 | 3 | 0.265 | 0.078 | 0.901 | −2.127 | 0.033 | |||
| F4 + F18 | 3 | 0.073 | 0.012 | 0.460 | −2.786 | 0.005 | |||
| F4 + F18 + LT | 2 | 0.031 | 0.002 | 0.483 | −2.481 | 0.013 | |||
| F4 + LT | 2 | 0.016 | 0.002 | 0.132 | −3.840 | 0.000 | |||
| LT | 2 | 1.022 | 0.252 | 4.155 | 0.031 | 0.975 | |||
| Overall | 15 | 0.301 | 0.208 | 0.435 | −6.385 | 0.000 | 15.789 | 5 | 0.007 |
| Groups | Number of Studies | Effect Size and 95% CI | Test of Null (Two-Tail) | Heterogeneity | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Point Estimate | Standard Error | Variance | Lower Limit | Upper Limit | Z-Value | p-Value | Q-Value | df(Q) | p-Value | ||
| F18 | 2 | −1.334 | 0.754 | 0.568 | −2.811 | 0.144 | −1.769 | 0.077 | |||
| F4 | 9 | 0.332 | 0.339 | 0.115 | −0.332 | 0.996 | 0.980 | 0.327 | |||
| F4 + F18 | 5 | 1.608 | 0.445 | 0.198 | 0.737 | 2.480 | 3.618 | 0.000 | |||
| F4 + LT | 1 | 3.203 | 0.997 | 0.993 | 1.250 | 5.156 | 3.214 | 0.001 | |||
| LT | 1 | 0.681 | 0.979 | 0.958 | −1.238 | 2.599 | 0.695 | 0.487 | |||
| Overall | 18 | 0.718 | 0.239 | 0.057 | 0.250 | 1.185 | 3.010 | 0.003 | 18.931 | 4 | 0.001 |
| Outcome | Vaccine Antigen | Antigen ID |
|---|---|---|
| Diarrhea | F18 + LT + ST | A |
| F4 | B | |
| F4 + LT | C | |
| LT | D | |
| F4 + F18 | E | |
| F18ac | F | |
| F4ab + F4ac + F5 + F6 + F41 F4ab + F4ac + F4ad + F5 + F6 + F41 | Gi Ga | |
| F4 + F18 + LT | H | |
| F4 + LT + ST | I | |
| Mortality | F18 + LT + ST | A |
| F4 | B | |
| F4 + LT | C | |
| LT | D | |
| F4 + F18 | E | |
| F4 + F18 + LT | H | |
| Average daily weight gain (ADWG) | F4 | B |
| F4 + LT | C | |
| LT | D | |
| F4 + F18 | E | |
| F18ac | F | |
| F4 + D3 | Bi | |
| F4 + Cpg | Ba |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ntakiyisumba, E.; Lee, S.; Won, G. Evidence-Based Approaches for Determining Effective Target Antigens to Develop Vaccines against Post-Weaning Diarrhea Caused by Enterotoxigenic Escherichia coli in Pigs: A Systematic Review and Network Meta-Analysis. Animals 2022, 12, 2136. https://doi.org/10.3390/ani12162136
Ntakiyisumba E, Lee S, Won G. Evidence-Based Approaches for Determining Effective Target Antigens to Develop Vaccines against Post-Weaning Diarrhea Caused by Enterotoxigenic Escherichia coli in Pigs: A Systematic Review and Network Meta-Analysis. Animals. 2022; 12(16):2136. https://doi.org/10.3390/ani12162136
Chicago/Turabian StyleNtakiyisumba, Eurade, Simin Lee, and Gayeon Won. 2022. "Evidence-Based Approaches for Determining Effective Target Antigens to Develop Vaccines against Post-Weaning Diarrhea Caused by Enterotoxigenic Escherichia coli in Pigs: A Systematic Review and Network Meta-Analysis" Animals 12, no. 16: 2136. https://doi.org/10.3390/ani12162136