
MHCII Expression on Peripheral Blood Monocytes in Canine Lymphoma: An Impact of Glucocorticoids

 , , and
, , and 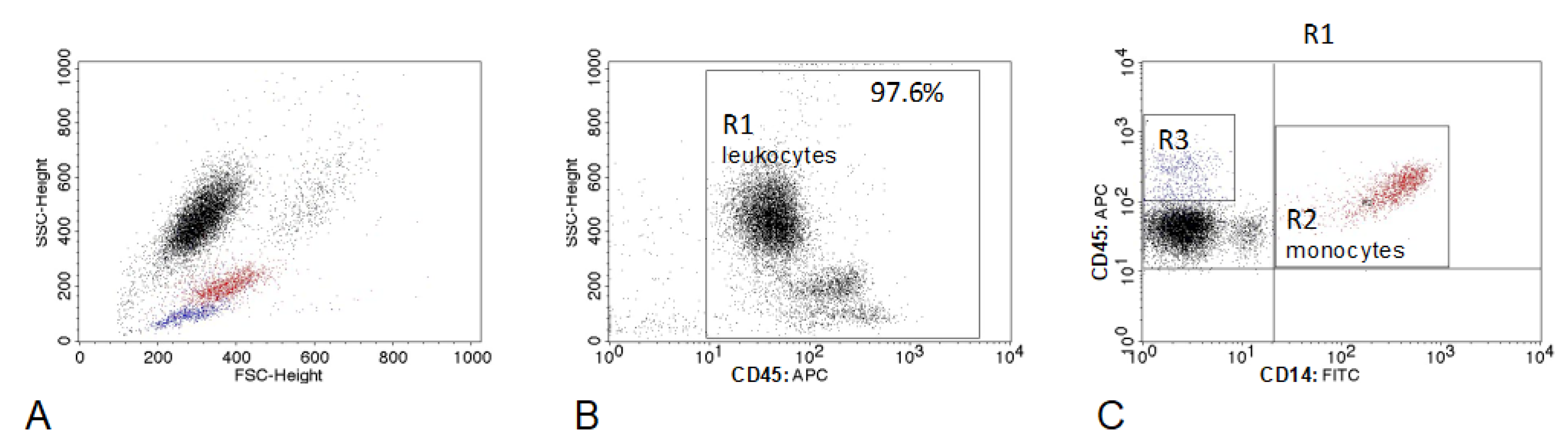{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Dogs and Sample Collection
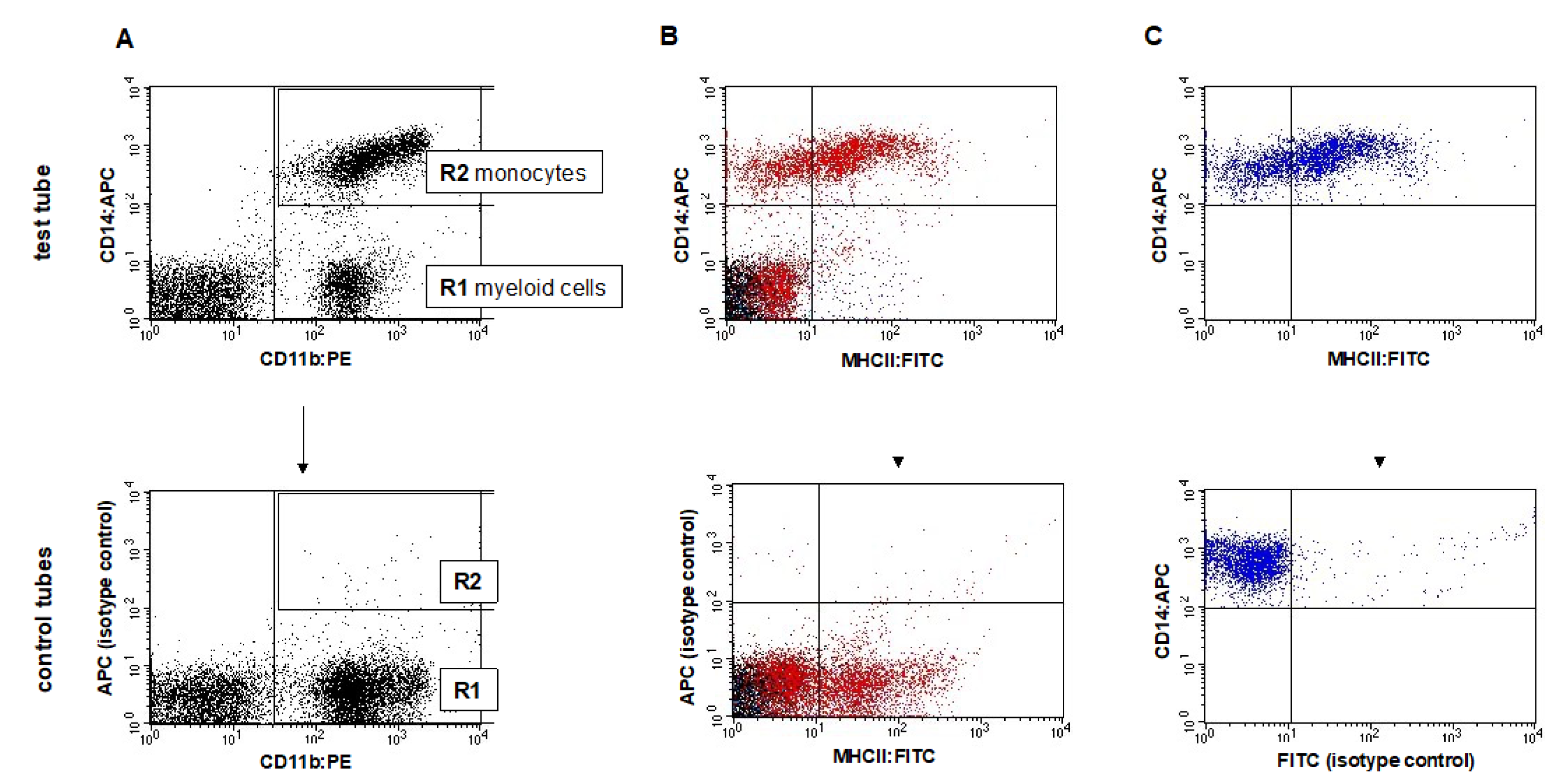
2.2. Flow Cytometry
2.3. Statistical Analysis
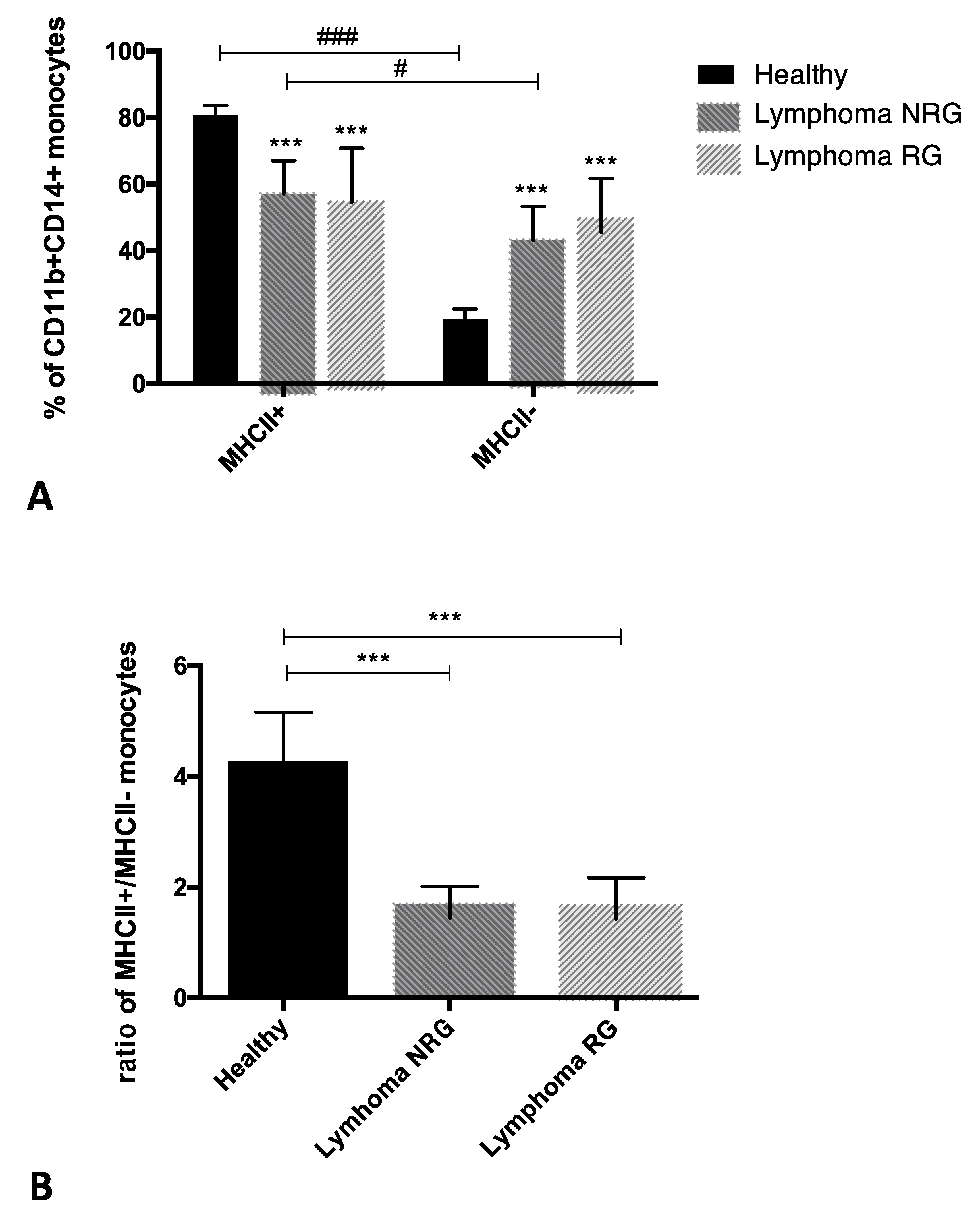
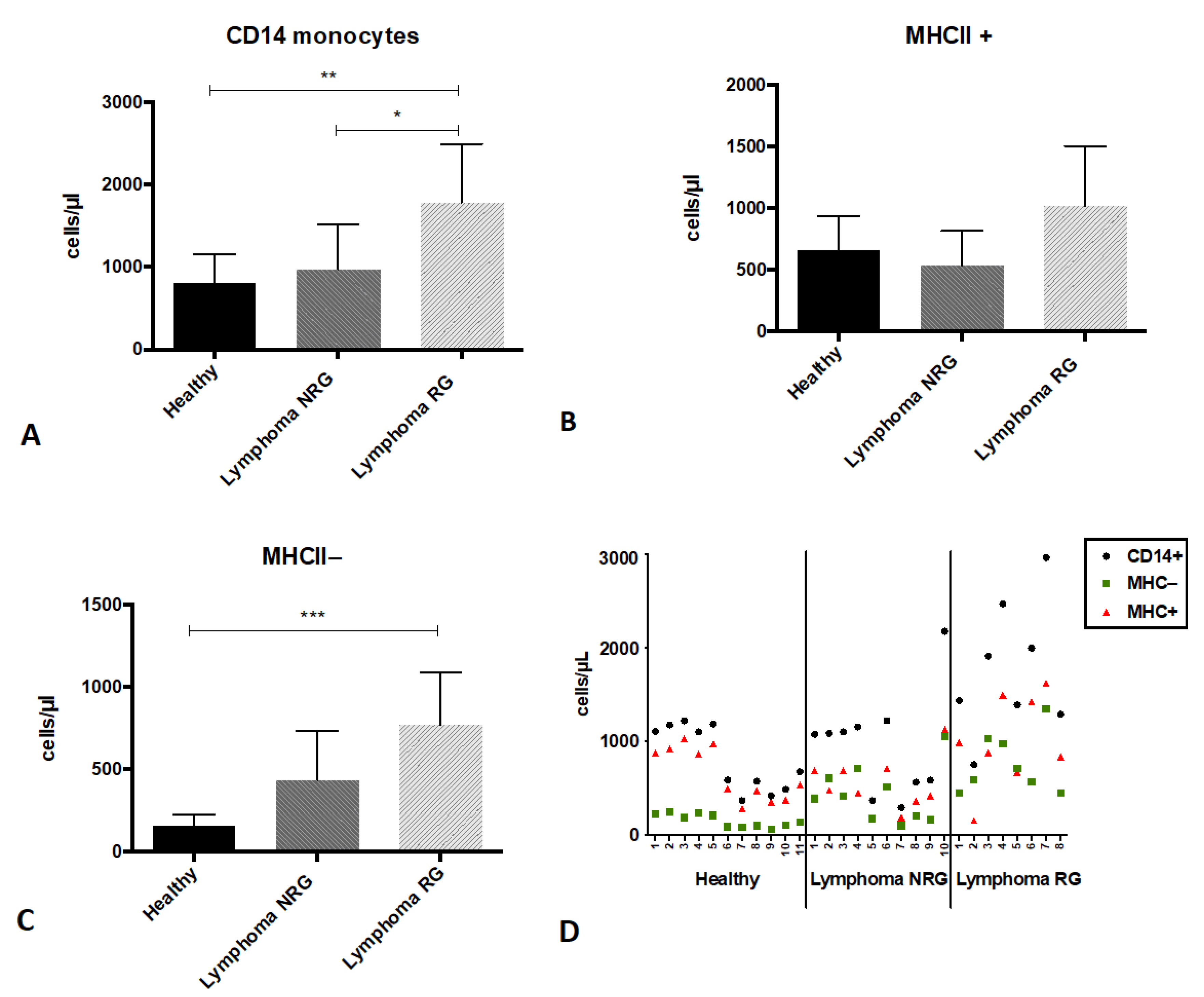
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Holling, T.M.; Schooten, E.; van Den Elsen, P.J. Function and regulation of MHC class II molecules in T-lymphocytes: Of mice and men. Hum. Immunol. 2004, 65, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Ono, S.; Tsujimoto, H.; Matsumoto, A.; Ikuta, S.; Kinoshita, M.; Mochizuki, H. Modulation of human leukocyte antigen-DR on monocytes and CD16 on granulocytes in patients with septic shock using hemoperfusion with polymyxin B-immobilized fiber. Am. J. Surg. 2004, 188, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.M.; Yu, Y.; Chai, J.K.; Hu, S.; Sheng, Z.Y.; Yao, Y.M. Low HLA-DR expression on CD14+ monocytes of burn victims with sepsis, and the effect of carbachol in vitro. Burns 2008, 34, 1158–1162. [Google Scholar] [CrossRef] [PubMed]
- Winkler, M.S.; Rissiek, A.; Priefler, M.; Schwedhelm, E.; Robbe, L.; Bauer, A.; Zahrte, C.; Zoellner, C.; Kluge, S.; Nierhaus, A. Human leucocyte antigen (HLA-DR) gene expression is reduced in sepsis and correlates with impaired TNFα response: A diagnostic tool for immunosuppression? PLoS ONE 2017, 12, e0182427. [Google Scholar] [CrossRef]
- Weiss, D.J.; Welle, M.; Mortiz, A.; Walcheck, B. Evaluation of leukocyte cell surface markers in dogs with septic and nonseptic inflammatory diseases. Am. J. Vet. Res. 2004, 65, 59–63. [Google Scholar] [CrossRef]
- Homma, S.; Komita, H.; Sagawa, Y.; Ohno, T.; Toda, G. Antitumour activity mediated by CD4+ cytotoxic T lymphocytes against MHC class II-negative mouse hepatocellular carcinoma induced by dendritic cell vaccine and interleukin-12. Immunology 2005, 115, 451–461. [Google Scholar] [CrossRef]
- Kitano, S.; Tsuji, T.; Liu, C.; Hirschhorn-Cymerman, D.; Kyi, C.; Mu, Z.; Allison, J.P.; Gnjatic, S.; Yuan, J.D.; Wolchok, J.D. Enhancement of tumor-reactive cytotoxic CD4+ T cell responses after ipilimumab treatment in four advanced melanoma patients. Cancer Immunol. Res. 2013, 1, 235–244. [Google Scholar] [CrossRef] [Green Version]
- Quezada, S.A.; Peggs, K.S. Tumor-reactive CD4+ T cells: Plasticity beyond helper and regulatory activities. Immunotherapy 2011, 3, 915–917. [Google Scholar] [CrossRef]
- Sun, Z.; Chen, F.; Meng, F.; Wei, J.; Liu, B. MHC class II restricted neoantigen: A promising target in tumor immunotherapy. Cancer Lett. 2017, 392, 17–25. [Google Scholar] [CrossRef]
- Javeed, N.; Gustafson, M.P.; Dutta, S.K.; Lin, Y.; Bamlet, W.R.; Oberg, A.L.; Petersen, G.M.; Chari, S.T.; Dietz, A.B.; Mukhopadhyay, D. Immunosuppressive CD14+HLA-DRlo/neg monocytes are elevated in pancreatic cancer and "primed" by tumor-derived exosomes. Oncoimmunology 2016, 6, e1252013. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.; Gustafson, M.P.; Bulur, P.A.; Gastineau, D.A.; Witzig, T.E.; Dietz, A.B. Immunosuppressive CD14+HLA-DR(low)/- monocytes in B-cell non-Hodgkin lymphoma. Blood 2011, 117, 872–881. [Google Scholar] [CrossRef] [Green Version]
- Vuk-Pavlović, S.; Bulur, P.A.; Lin, Y.; Qin, R.; Szumlanski, C.L.; Zhao, X.; Dietz, A.B. Immunosuppressive CD14+HLA-DRlow/- monocytes in prostate cancer. Prostate 2010, 70, 443–455. [Google Scholar] [CrossRef] [Green Version]
- Xiu, B.; Lin, Y.; Grote, D.M.; Ziesmer, S.C.; Gustafson, M.P.; Maas, M.L.; Zhang, Z.; Dietz, A.B.; Porrata, L.F.; Novak, A.J.; et al. IL-10 induces the development of immunosuppressive CD14(+)HLA-DR(low/-) monocytes in B-cell non-Hodgkin lymphoma. Blood Cancer J. 2015, 5, e328. [Google Scholar] [CrossRef] [Green Version]
- Ito, D.; Frantz, A.M.; Modiano, J.F. Canine lymphoma as a comparative model for human non-Hodgkin lymphoma: Recent progress and applications. Vet. Immunol. Immunopathol. 2014, 159, 192–201. [Google Scholar] [CrossRef] [Green Version]
- Marconato, L.; Gelain, M.E.; Comazzi, S. The dog as a possible animal model for human non-Hodgkin lymphoma: A review. Hematol. Oncol. 2013, 31, 1–9. [Google Scholar] [CrossRef]
- Richards, K.L.; Suter, S.E. Man's best friend: What can pet dogs teach us about non-Hodgkin's lymphoma? Immunol. Rev. 2015, 263, 173–191. [Google Scholar] [CrossRef]
- Seelig, D.M.; Avery, A.C.; Ehrhart, E.J.; Linden, M.A. The Comparative Diagnostic Features of Canine and Human Lymphoma. J. Vet. Sci. 2016, 3, 11. [Google Scholar] [CrossRef]
- Zandvliet, M. Canine lymphoma: A review. Vet. Q. 2016, 36, 76–104. [Google Scholar] [CrossRef]
- Chun, R. Lymphoma: Which chemotherapy protocol and why? Top. Companion Anim. Med. 2009, 24, 157–162. [Google Scholar] [CrossRef]
- Lautscham, E.M.; Kessler, M.; Ernst, T.; Willimzig, L.; Neiger, R. Comparison of a CHOP-LAsp-based protocol with and without maintenance for canine multicentric lymphoma. Vet. Rec. 2017, 180, 303. [Google Scholar] [CrossRef]
- Jacobsen, C.N.; Aasted, B.; Broe, M.K.; Petersen, J.L. Reactivities of 20 anti-human monoclonal antibodies with leucocytes from ten different animal species. Vet. Immunol. Immunopathol. 1993, 39, 461–466. [Google Scholar] [CrossRef]
- Lawrence, J.; Chang, Y.M.; Szladovits, B.; Davison, L.J.; Garden, O.A. Breed-specific hematological phenotypes in the dog: A natural resource for the genetic dissection of hematological parameters in a mammalian species. PLoS ONE 2013, 8, e81288. [Google Scholar] [CrossRef]
- Kim, O.Y.; Monsel, A.; Bertrand, M.; Coriat, P.; Cavaillon, J.M.; Adib-Conquy, M. Differential down-regulation of HLA-DR on monocyte subpopulations during systemic inflammation. Crit. Care. 2010, 14, R61. [Google Scholar] [CrossRef] [Green Version]
- Rocha, N.; Neefjes, J. MHC class II molecules on the move for successful antigen presentation. EMBO J. 2008, 27, 1–5. [Google Scholar] [CrossRef]
- Giroux, M.; Schmidt, M.; Descoteaux, A. IFN-gamma-induced MHC class II expression: Transactivation of class II transactivator promoter IV by IFN regulatory factor-1 is regulated by protein kinase C-alpha. J. Immunol. 2003, 171, 4187–4194. [Google Scholar] [CrossRef] [Green Version]
- Thelemann, C.; Eren, R.O.; Coutaz, M.; Brasseit, J.; Bouzourene, H.; Rosa, M.; Duval, A.; Lavanchy, C.; Mack, V.; Mueller, C.; et al. Interferon-γ induces expression of MHC class II on intestinal epithelial cells and protects mice from colitis. PLoS ONE 2014, 9, e86844. [Google Scholar] [CrossRef] [Green Version]
- Schroder, K.; Hertzog, P.J.; Ravasi, T.; Hume, D.A. Interferon-gamma: An overview of signals, mechanisms and functions. J. Leukoc. Biol. 2004, 75, 163–189. [Google Scholar] [CrossRef]
- Zaidi, M.R.; Merlino, G. The two faces of interferon-γ in cancer. Clin. Cancer Res. 2011, 17, 6118–6124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klotz, D.; Baumgärtner, W.; Gerhauser, I. Type I interferons in the pathogenesis and treatment of canine diseases. Vet. Immunol. Immunopathol. 2017, 191, 80–93. [Google Scholar] [CrossRef] [PubMed]
- Calvalido, J.; Wood, G.A.; Mutsaers, A.J.; Wood, D.; Sears, W.; Woods, J.P. Comparison of serum cytokine levels between dogs with multicentric lymphoma and healthy dogs. Vet. Immunol. Immunopathol. 2016, 182, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Cho, K.J.; Roche, P.A. Regulation of MHC Class II-Peptide Complex Expression by Ubiquitination. Front. Immunol. 2013, 4, 369. [Google Scholar] [CrossRef] [Green Version]
- Koppelman, B.; Neefjes, J.J.; de Vries, J.E.; de Waal Malefyt, R. Interleukin-10 down- regulates MHC class II alphabeta peptide complexes at the plasma membrane of monocytes by affecting arrival and recycling. Immunity 1997, 7, 861–871. [Google Scholar] [CrossRef] [Green Version]
- Thibodeau, J.; Bourgeois-Daigneault, M.C.; Huppé, G.; Tremblay, J.; Aumont, A.; Houde, M.; Bartee, E.; Brunet, A.; Gauvreau, M.E.; de Gassart, A.; et al. Interleukin-10-induced MARCH1 mediates intracellular sequestration of MHC class II in monocytes. Eur. J. Immunol. 2008, 38, 1225–1230. [Google Scholar] [CrossRef] [Green Version]
- de Waal Malefyt, R.; Haanen, J.; Spits, H.; Roncarolo, M.G.; te Velde, A.; Figdor, C.; Johnson, K.; Kastelein, R.; Yssel, H.; de Vries, J.E. Interleukin 10 (IL-10) and viral IL-10 strongly reduce antigen-specific human T cell proliferation by diminishing the antigen- presenting capacity of monocytes via downregulation of class II major histocompatibility complex expression. J. Exp. Med. 1991, 174, 915–924. [Google Scholar] [CrossRef]
- Bohlen, H.; Kessler, M.; Sextro, M.; Diehl, V.; Tesch, H. Poor clinical outcome of patients with Hodgkin's disease and elevated interleukin-10 serum levels. Clinical significance of interleukin-10 serum levels for Hodgkin's disease. Ann. Hematol. 2000, 79, 110–113. [Google Scholar] [CrossRef]
- Gupta, M.; Han, J.J.; Stenson, M.; Maurer, M.; Wellik, L.; Hu, G.; Ziesmer, S.; Dogan, A.; Witzig, T.E. Elevated serum IL-10 levels in diffuse large B-cell lymphoma: A mechanism of aberrant JAK2 activation. Blood 2012, 119, 2844–2853. [Google Scholar] [CrossRef]
- Blay, J.Y.; Burdin, N.; Rousset, F.; Lenoir, G.; Biron, P.; Philip, T.; Banchereau, J.; Favrot, M.C. Serum interleukin-10 in non-Hodgkin's lymphoma: A prognostic factor. Blood 1993, 82, 2169–2174. [Google Scholar] [CrossRef] [Green Version]
- Vassilakopoulos, T.P.; Nadali, G.; Angelopoulou, M.K.; Siakantaris, M.P.; Dimopoulou, M.N.; Kontopidou, F.N.; Rassidakis, G.Z.; Doussis-Anagnostopoulou, I.A.; Hatzioannou, M.; Vaiopoulos, G.; et al. Serum interleukin-10 levels are an independent prognostic factor for patients with Hodgkin's lymphoma. Haematologica 2001, 86, 274–281. [Google Scholar]
- Benjamin, D.; Knobloch, T.J.; Dayton, M.A. Human B-cell interleukin-10: B-cell lines derived from patients with acquired immunodeficiency syndrome and Burkitt's lymphoma constitutively secrete large quantities of interleukin-10. Blood 1992, 80, 1289–1298. [Google Scholar] [CrossRef] [Green Version]
- Masood, R.; Zhang, Y.; Bond, M.W.; Scadden, D.T.; Moudgil, T.; Law, R.E.; Kaplan, M.H.; Jung, B.; Espina, B.M.; Lunardi-Iskandar, Y. Interleukin-10 is an autocrine growth factor for acquired immunodeficiency syndrome-related B-cell lymphoma. Blood 1995, 85, 3423–3430. [Google Scholar] [CrossRef] [Green Version]
- O'Garra, A.; Stapleton, G.; Dhar, V.; Pearce, M.; Schumacher, J.; Rugo, H.; Barbis, D.; Stall, A.; Cupp, J.; Moore, K.; et al. Production of cytokines by mouse B cells: B lymphomas and normal B cells produce interleukin 10. Int. Immunol. 1990, 2, 821–832. [Google Scholar] [CrossRef]
- Sherger, M.; Kisseberth, W.; London, C.; Olivo-Marston, S.; Papenfuss, T.L. Identification of myeloid derived suppressor cells in the peripheral blood of tumor bearing dogs. BMC Vet. Res. 2012, 8, 209. [Google Scholar] [CrossRef] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rzepecka, A.; Jagielski, D.; Cywińska, A.; Sapierzyński, R.; Żmigrodzka, M.; Witkowska-Piłaszewicz, O.; Winnicka, A. MHCII Expression on Peripheral Blood Monocytes in Canine Lymphoma: An Impact of Glucocorticoids. Animals 2022, 12, 2135. https://doi.org/10.3390/ani12162135
Rzepecka A, Jagielski D, Cywińska A, Sapierzyński R, Żmigrodzka M, Witkowska-Piłaszewicz O, Winnicka A. MHCII Expression on Peripheral Blood Monocytes in Canine Lymphoma: An Impact of Glucocorticoids. Animals. 2022; 12(16):2135. https://doi.org/10.3390/ani12162135
Chicago/Turabian StyleRzepecka, Alicja, Dariusz Jagielski, Anna Cywińska, Rafał Sapierzyński, Magdalena Żmigrodzka, Olga Witkowska-Piłaszewicz, and Anna Winnicka. 2022. "MHCII Expression on Peripheral Blood Monocytes in Canine Lymphoma: An Impact of Glucocorticoids" Animals 12, no. 16: 2135. https://doi.org/10.3390/ani12162135