
BMP6 Promotes the Secretion of 17 Beta-Estradiol and Progesterone in Goat Ovarian Granulosa Cells

 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Cell Cultures and Treatment
2.3. Cellular Immunochemical Staining
2.4. Ovarian Immunohistochemistry
2.5. EdU Cell Proliferation Measurements
2.6. Apoptosis and Cell Cycle Measurements
2.7. Steroid Assay
2.8. Quantitative Real-Time PCR
2.9. Western Blotting
2.10. Statistical Analysis
3. Results
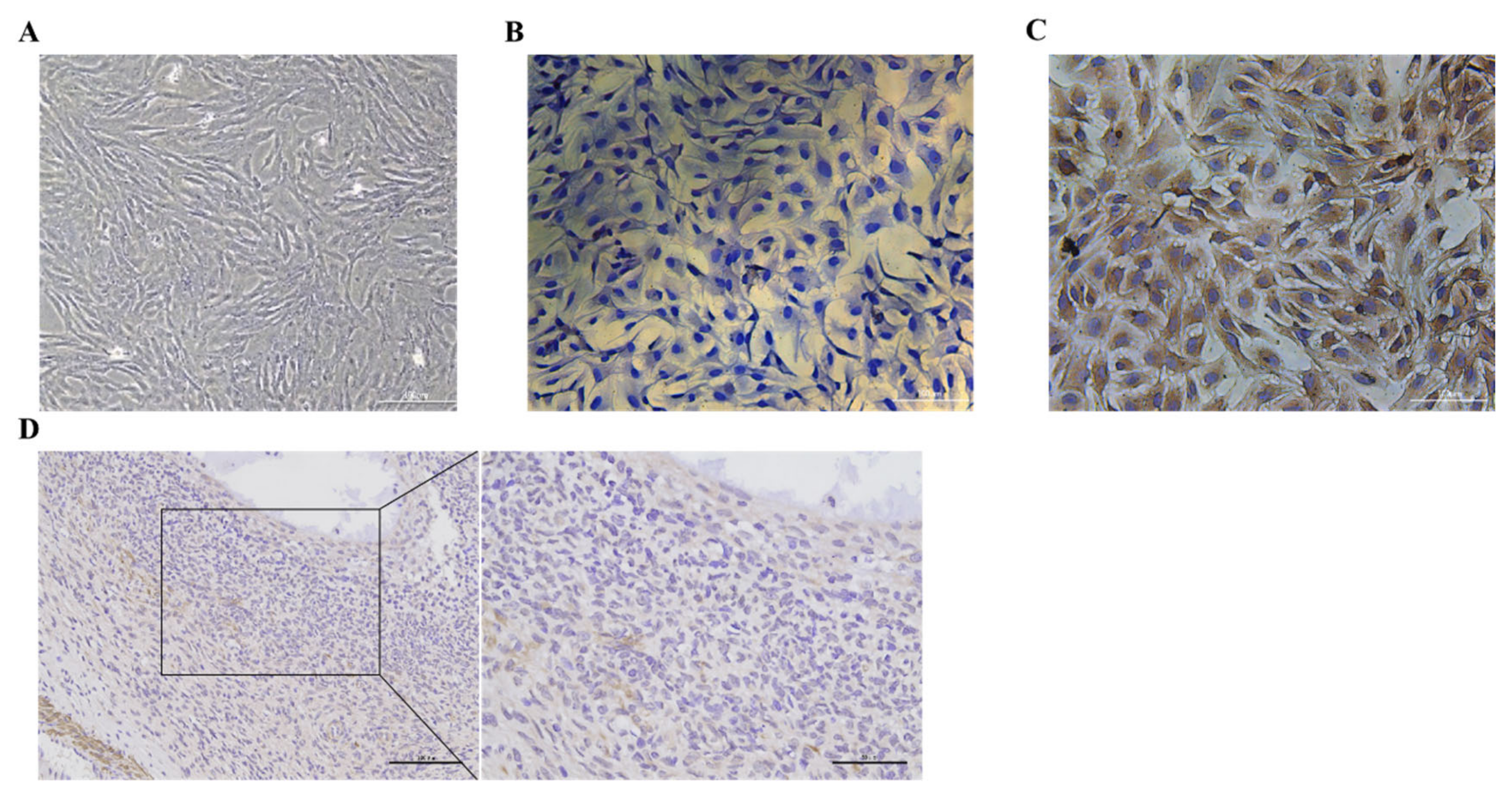
3.1. Identification of Goat Ovarian Granulosa Cells and Expression of BMP6 in Ovaries
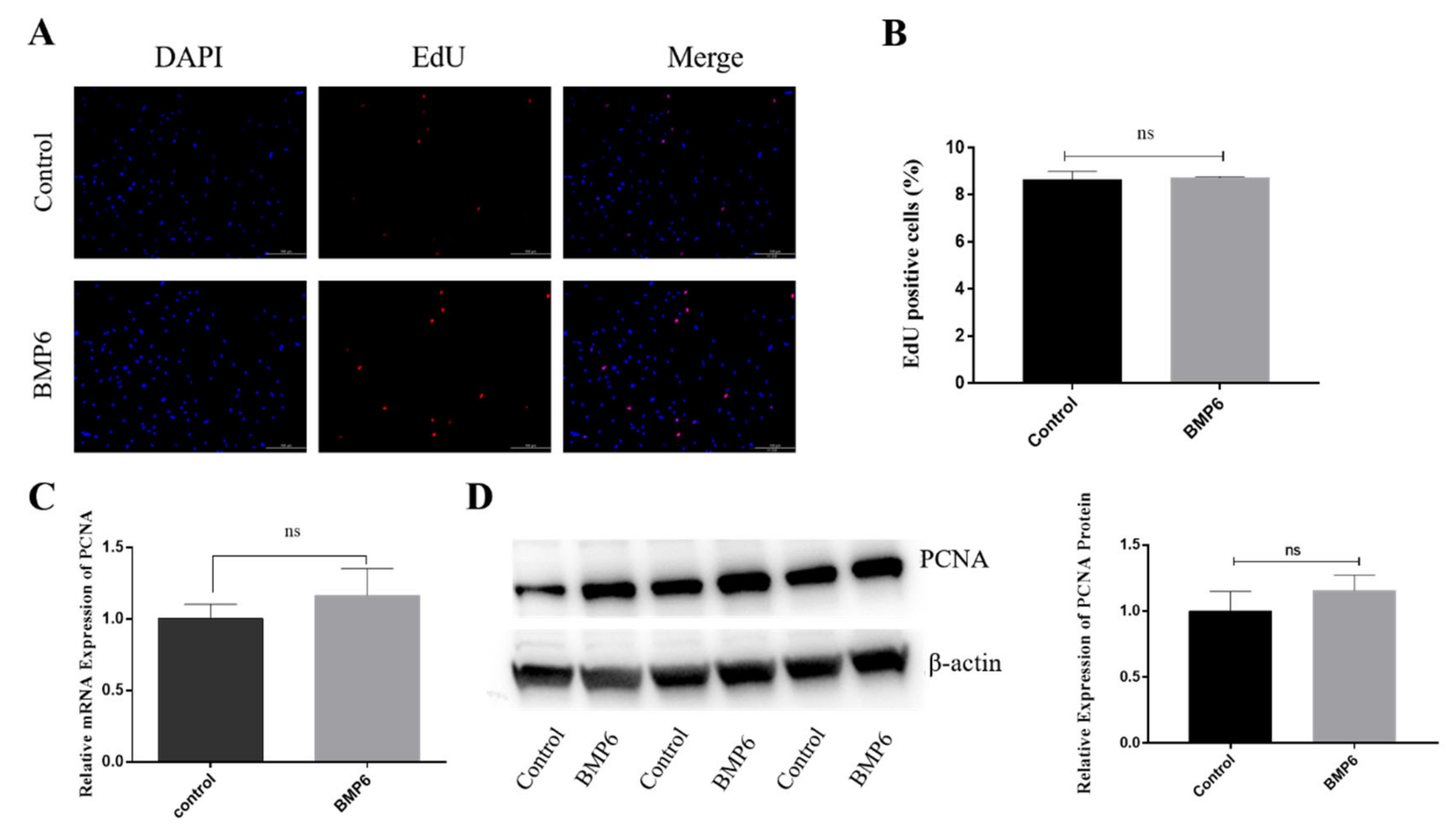
3.2. Effects of BMP6 on Proliferation in GCs
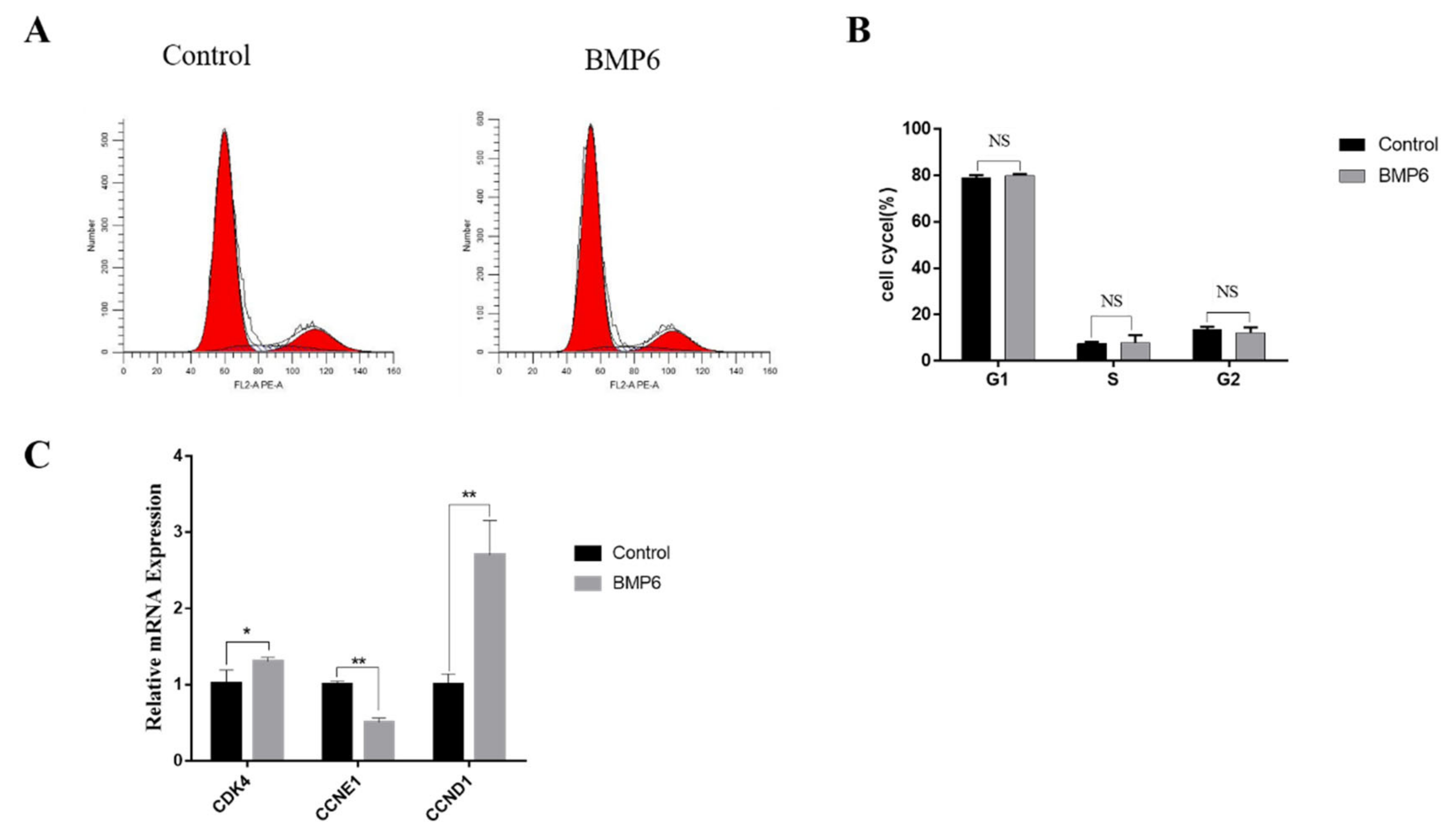
3.3. Effects of BMP6 on Cell Cycle in GCs
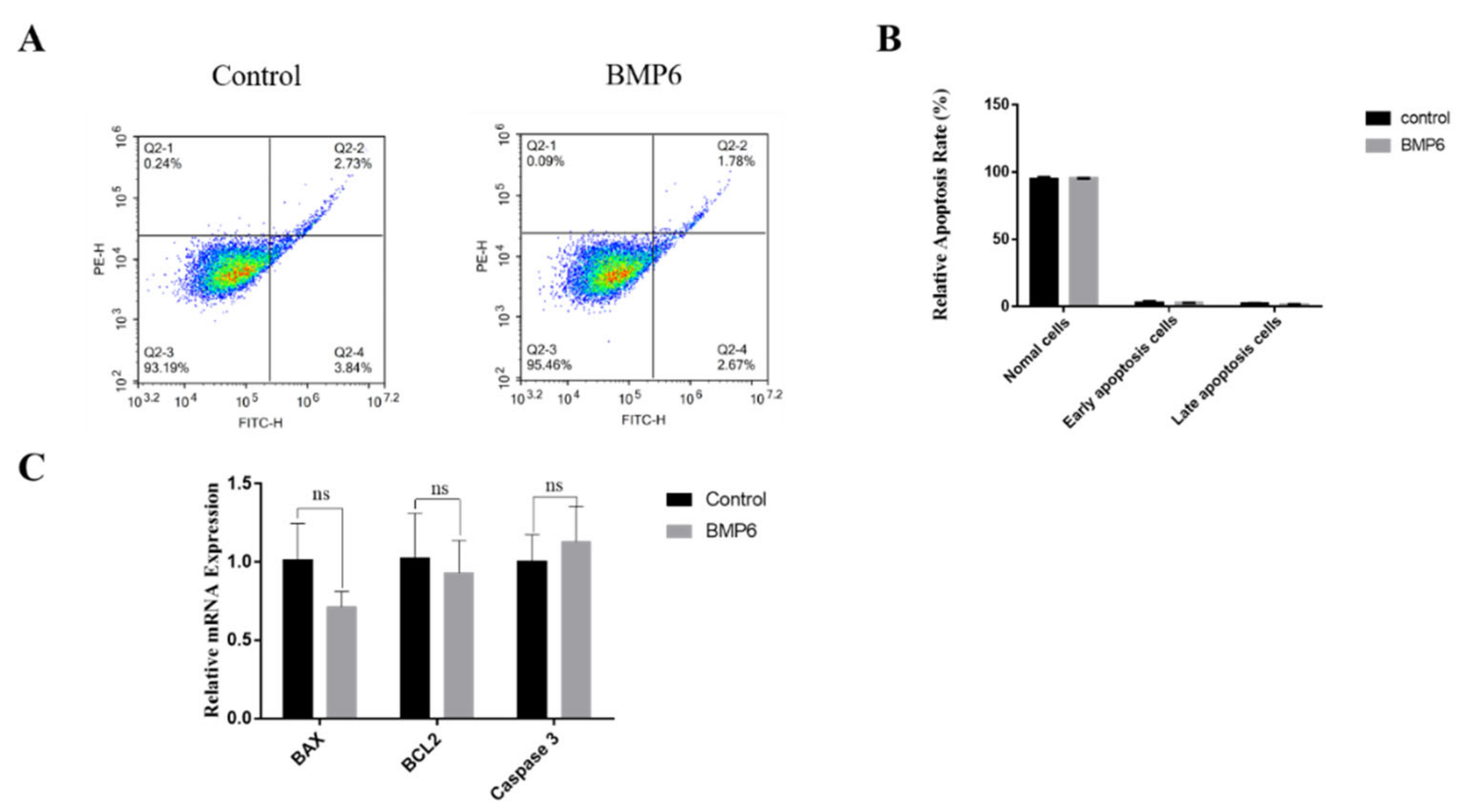
3.4. Effects of BMP6 on Apoptosis in GCs
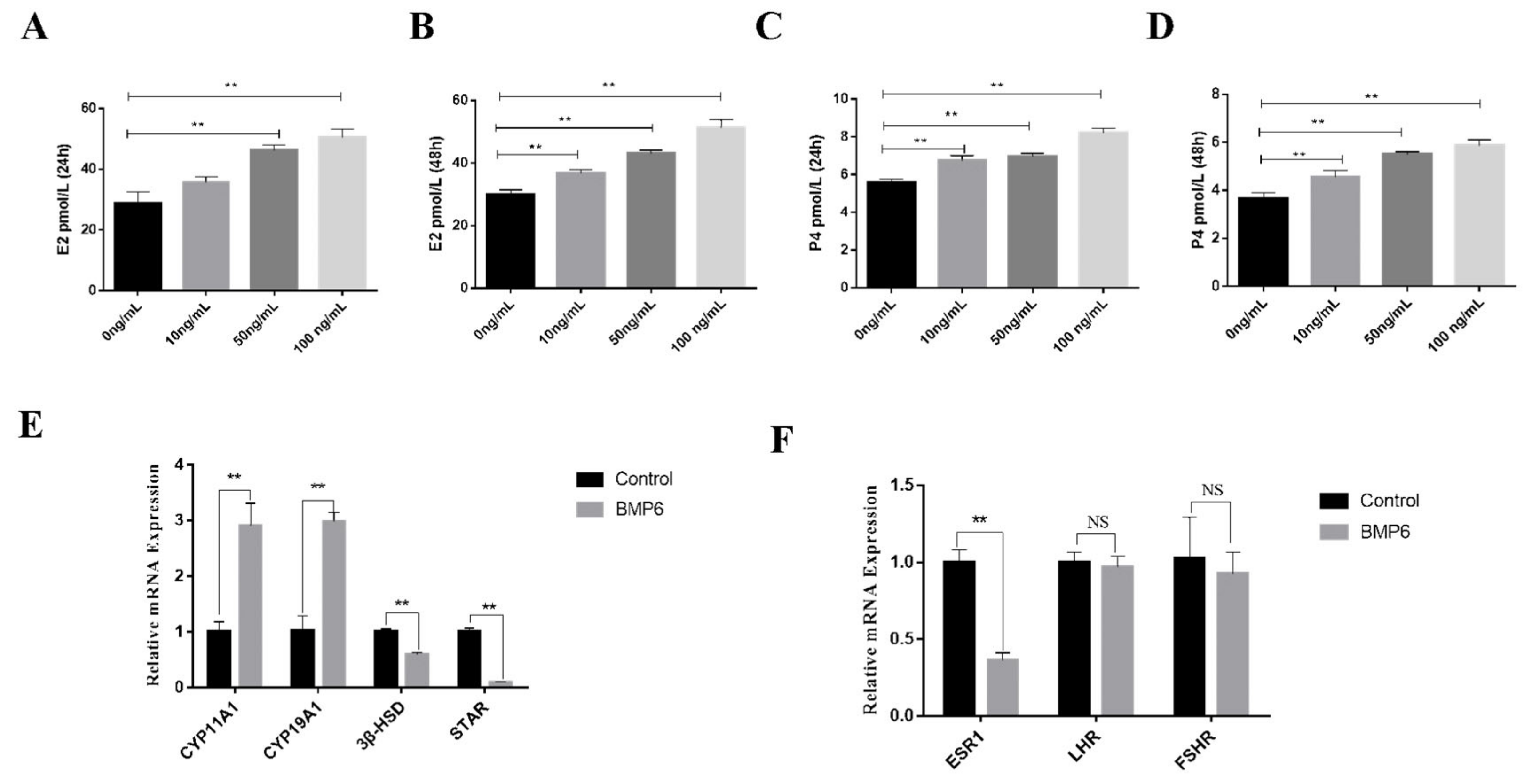
3.5. Effects of BMP6 on Steroid Hormones’ Secretion in GCs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- He, Y.M.; Deng, H.H.; Jiang, Z.L.; Li, Q.W.; Shi, M.H.; Chen, H.L.; Han, Z.S. Effects of melatonin on follicular atresia and granulosa cell apoptosis in the porcine. Mol. Reprod. Dev. 2016, 83, 692–700. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; He, H.; Jiang, X.; Hua, R.; Chen, H.; Yang, L.; Cheng, J.; Duan, J.; Li, Q. Sirt2 plays a novel role on progesterone, estradiol and testosterone synthesis via ppars/lxrα pathways in bovine ovarian granular cells. J. Steroid. Biochem. Mol. Biol. 2018, 185, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Austin, E.J.; Mihm, M.; Evans, A.C.O.; Knight, P.G.; Ireland, J.L.H.; Ireland, J.J.; Roche, J.F. Alterations in intrafollicular regulatory factors and apoptosis during selection of follicles in the first follicular wave of the bovine estrous cycle. Biol. Reprod. 2001, 64, 839–848. [Google Scholar] [CrossRef] [PubMed]
- Ginther, O.J.; Beg, M.A.; Kot, K.; Meira, C.; Bergfelt, D.R. Associated and independent comparisons between the two largest follicles preceding follicle deviation in cattle. Biol. Reprod. 2003, 68, 524–529. [Google Scholar] [CrossRef] [PubMed]
- Cai, L.P.; Sun, A.D.; Li, H.; Tsinkgou, A.; Yu, J.N.; Ying, S.J.; Chen, Z.; Shi, Z.D. Molecular mechanisms of enhancing porcine granulosa cell proliferation and function by treatment in vitro with anti-inhibin alpha subunit antibody. Reprod. Biol. Endocrinol. 2015, 13, 26. [Google Scholar] [CrossRef]
- Cerbito, W.A.; Miyamoto, A.; Balagapo, C.R.; Natural, N.G.; Miyazawa, K.; Sato, K. Prostaglandin e(2) levels in uterine tissues and its relationship with uterine and luteal progesterone during the estrous-cycle in dairy-cows. Theriogenology 1994, 42, 941–950. [Google Scholar] [CrossRef]
- Carvalho, J.B.P.; Carvalho, N.A.T.; Reis, E.L.; Nichi, M.; Souza, A.H.; Baruselli, P.S. Effect of early luteolysis in progesterone-based al protocols in bos indicus, bos indicus x bos taurus and bos taurus heifers. Theriogenology 2008, 69, 167–175. [Google Scholar] [CrossRef]
- Abreu, F.M.; Geary, T.W.; da Siva, M.A.C.; Cruppe, L.H.; Mussard, M.L.; Madsen, C.A.; Martins, T.; Bridges, G.A.; Harstine, B.R.; Day, M.L. Role of progesterone concentrations during early follicular development in beef cattle: Ii. Ovulatory follicle growth and chock for pregnancy rates. Anim. Reprod. Sci. 2018, 196, 69–76. [Google Scholar] [CrossRef]
- King, S.R.; LaVoie, H.A. Gonadal transactivation of stard1, cyp11a1 and hsd3b. Front. Biosci. 2012, 17, 824–846. [Google Scholar] [CrossRef]
- Ernst, E.H.; Franks, S.; Hardy, K.; Villesen, P.; Lykke-Hartmann, K. Granulosa cells from human primordial and primary follicles show differential global gene expression profiles. Hum. Reprod. 2018, 33, 666–679. [Google Scholar] [CrossRef]
- Lazaros, L.A.; Hatzi, E.G.; Xita, N.V.; Makrydimas, G.V.; Kaponis, A.I.; Takenaka, A.; Kosmas, I.P.; Sofikitis, N.V.; Stefos, T.I.; Zikopoulos, K.A.; et al. Aromatase (cyp19) gene variants influence ovarian response to standard gonadotrophin stimulation. J. Assist. Reprod. Genet. 2012, 29, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wang, S.B.; Xu, Y.X. Signal transduction of bmp/smad and its relationship with mammalian folliculogenesis. Yi Chuan 2009, 31, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Shimasaki, S.; Moore, R.K.; Otsuka, F.; Erickson, G.F. The bone morphogenetic protein system in mammalian reproduction. Endocr. Rev. 2004, 25, 72–101. [Google Scholar] [CrossRef]
- Khan, D.R.; Fournier, É.; Dufort, I.; Richard, F.J.; Singh, J.; Sirard, M.A. Meta-analysis of gene expression profiles in granulosa cells during folliculogenesis. Reproduction 2016, 151, R103–R110. [Google Scholar] [CrossRef]
- Khalaf, M.; Morera, J.; Bourret, A.; Reznik, Y.; Denoual, C.; Herlicoviez, M.; Mittre, H.; Benhaim, A. Bmp system expression in gcs from polycystic ovary syndrome women and the in vitro effects of bmp4, bmp6, and bmp7 on gc steroidogenesis. Eur. J. Endocrinol. 2013, 168, 437–444. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, S.G.; Andersen, K.; Clement, C.A.; Franks, S.; Hardy, K.; Andersen, C.Y. Expression of tgf-beta superfamily growth factors, their receptors, the associated smads and antagonists in five isolated size-matched populations of pre-antral follicles from normal human ovaries. Mol. Hum. Reprod. 2014, 20, 293–308. [Google Scholar] [CrossRef] [PubMed]
- Erickson, G.F.; Shimasaki, S. The spatiotemporal expression pattern of the bone morphogenetic protein family in rat ovary cell types during the estrous cycle. Reprod. Biol. Endocrinol. 2003, 1, 9. [Google Scholar] [CrossRef]
- Frota, I.M.A.; Leitao, C.C.F.; Costa, J.J.N.; van den Hurk, R.; Saraiva, M.V.A.; Figueiredo, J.R.; Silva, J.R.V. Levels of bmp-6 mrna in goat ovarian follicles and in vitro effects of bmp-6 on secondary follicle development. Zygote 2013, 21, 270–278. [Google Scholar] [CrossRef]
- Lochab, A.K.; Extavour, C.G. Bone morphogenetic protein (bmp) signaling in animal reproductive system development and function. Dev. Biol. 2017, 427, 258–269. [Google Scholar] [CrossRef]
- Sugiura, K.; Su, Y.Q.; Eppig, J.J. Does bone morphogenetic protein 6 (bmp6) affect female fertility in the mouse? Biol. Reprod. 2010, 83, 997–1004. [Google Scholar] [CrossRef]
- Nio-Kobayashi, J.; Trendell, J.; Giakoumelou, S.; Boswell, L.; Nicol, L.; Kudo, M.; Sakuragi, N.; Iwanaga, T.; Duncan, W.C. Bone morphogenetic proteins are mediators of luteolysis in the human corpus luteum. Endocrinology 2015, 156, 1494–1503. [Google Scholar] [CrossRef] [PubMed]
- Otsuka, F.; Moore, R.K.; Shimasaki, S. Biological function and cellular mechanism of bone morphogenetic protein-6 in the ovary. J. Biol. Chem. 2001, 276, 32889–32895. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.Q.; Guo, F.Y.; Sun, X.W.; He, Q.J.; Dai, Z.N.; Chen, X.C.; Zhao, Y.J.; Wang, J. Bmp15 regulates amh expression via the p38 mapk pathway in granulosa cells from goat. Theriogenology 2018, 118, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Wu, G. The signaling and functions of heterodimeric bone morphogenetic proteins. Cytokine Growth Factor Rev. 2012, 23, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.Y.; Su, L.; Pan, X.Y.; Yao, J.; Li, Z.X.; Wang, X.N.; Xu, B.S. Effect of bmp-6 on development and maturation of mouse preantral follicles in vitro. Biotechnol. Biotechnol. Equip. 2015, 29, 336–344. [Google Scholar] [CrossRef]
- Yang, F.; He, L.; Luo, B.; Ye, F.; Cui, C.; Yu, X.; Xu, H.; Zhao, X.; Yin, H.; Li, D.; et al. Effect of bone morphogenetic protein 6 (bmp6) on chicken granulose cells proliferation and progesterone synthesis. Braz. J. Poultry Sci. 2019, 21, 1–8. [Google Scholar] [CrossRef]
- Jarrin, M.; Pandit, T.; Gunhaga, L. A balance of fgf and bmp signals regulates cell cycle exit and equarin expression in lens cells. Mol. Biol. Cell. 2012, 23, 3266–3274. [Google Scholar] [CrossRef]
- Hu, F.; Meng, X.; Tong, Q.; Liang, L.; Xiang, R.; Zhu, T.; Yang, S. Bmp-6 inhibits cell proliferation by targeting microrna-192 in breast cancer. Biochim. Biophys. Acta 2013, 1832, 2379–2390. [Google Scholar] [CrossRef]
- Jiang, B.S.; Wang, E.S.; Donovan, K.A.; Liang, Y.K.; Fischer, E.S.; Zhang, T.H.; Gray, N.S. Development of dual and selective degraders of cyclin-dependent kinases 4 and 6. Angew. Chem. Int. Ed. Engl. 2019, 58, 6321–6326. [Google Scholar] [CrossRef]
- Romero-Pozuelo, J.; Figlia, G.; Kaya, O.; Martin-Villalba, A.; Teleman, A.A. Cdk4 and cdk6 couple the cell-cycle machinery to cell growth via mtorc1. Cell. Rep. 2020, 31, 107504. [Google Scholar] [CrossRef]
- Ma, L.; Li, J. Microrna-519d-3p inhibits cell proliferation and cell cycle g1/s transition in glioma by targeting ccnd1. Biosci. Biotechnol. Biochem. 2020, 84, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Möröy, T.; Geisen, C. Cyclin e. Int. J. Biochem. Cell. Biol. 2004, 36, 1424–1439. [Google Scholar] [CrossRef] [PubMed]
- Worku, T.; Rehman, Z.U.; Talpur, H.S.; Bhattarai, D.; Ullah, F.; Malobi, N.; Kebede, T.; Yang, L. Micrornas: New insight in modulating follicular atresia: A review. Int. J. Mol. Sci. 2017, 18, 333. [Google Scholar] [CrossRef] [PubMed]
- Hussein, T.S.; Froiland, D.A.; Amato, F.; Thompson, J.G.; Gilchrist, R.B. Oocytes prevent cumulus cell apoptosis by maintaining a morphogenic paracrine gradient of bone morphogenetic proteins. J. Cell. Sci. 2005, 118, 5257–5268. [Google Scholar] [CrossRef]
- Chen, H.Y.; Liu, C.; Jiang, H.; Gao, Y.; Xu, M.Q.; Wang, J.W.; Liu, S.Y.; Fu, Y.; Sun, X.L.; Xu, J.J.; et al. Regulatory role of mirna-375 in expression of bmp15/gdf9 receptors and its effect on proliferation and apoptosis of bovine cumulus cells. Cell. Physiol. Biochem. 2017, 41, 439–450. [Google Scholar] [CrossRef]
- Zhang, J.; Xu, Y.; Liu, H.; Pan, Z. Micrornas in ovarian follicular atresia and granulosa cell apoptosis. Reprod. Biol. Endocrinol. 2019, 17, 9. [Google Scholar] [CrossRef] [PubMed]
- Tarumi, W.; Itoh, M.T.; Suzuki, N. Effects of 5α-dihydrotestosterone and 17β-estradiol on the mouse ovarian follicle development and oocyte maturation. PLoS ONE 2014, 9, e99423. [Google Scholar] [CrossRef]
- Li, Q.; He, H.; Zhang, Y.L.; Li, X.M.; Guo, X.; Huo, R.; Bi, Y.; Li, J.; Fan, H.Y.; Sha, J. Phosphoinositide 3-kinase p110δ mediates estrogen- and fsh-stimulated ovarian follicle growth. Mol. Endocrinol. 2013, 27, 1468–1482. [Google Scholar] [CrossRef]
- Guo, M.; Zhang, C.; Wang, Y.; Feng, L.; Wang, Z.; Niu, W.; Du, X.; Tang, W.; Li, Y.; Wang, C.; et al. Progesterone receptor membrane component 1 mediates progesterone-induced suppression of oocyte meiotic prophase i and primordial folliculogenesis. Sci. Rep. 2016, 6, 36869. [Google Scholar] [CrossRef]
- Yuan, X.H.; Yang, C.R.; Wang, X.N.; Zhang, L.L.; Gao, X.R.; Shi, Z.Y. Progesterone maintains the status of granulosa cells and slows follicle development partly through pgrmc1. J. Cell. Physiol. 2018, 234, 709–720. [Google Scholar] [CrossRef]
- Tokarz, J.; Möller, G.; de Angelis, M.H.; Adamski, J. Steroids in teleost fishes: A functional point of view. Steroids 2015, 103, 123–144. [Google Scholar] [CrossRef] [PubMed]
- Stocco, D.M. The role of the star protein in steroidogenesis: Challenges for the future. J. Endocrinol. 2000, 164, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Rhee, H.S.; Oh, S.H.; Ko, B.J.; Han, D.M.; Jeon, B.H.; Park, H.; Moon, H.B.; Kim, W.S. Expression of 3b-hydroxysteroid dehydrogenase and p450 side chain cleavage enzyme in the human uterine endometrium. Exp. Mol. Med. 2003, 35, 160–166. [Google Scholar] [CrossRef] [PubMed]
- Senthilkumaran, B.; Yoshikuni, M.; Nagahama, Y. A shift in steroidogenesis occurring in ovarian follicles prior to oocyte maturation. Mol. Cell. Endocrinol. 2004, 215, 11–18. [Google Scholar] [CrossRef]
- Yamashita, H.; Murayama, C.; Takasugi, R.; Miyamoto, A.; Shimizu, T. Bmp-4 suppresses progesterone production by inhibiting histone h3 acetylation of star in bovine granulosa cells in vitro. Mol. Cell. Biochem. 2011, 348, 183–190. [Google Scholar] [CrossRef]
- Chang, H.M.; Cheng, J.C.; Klausen, C.; Leung, P.C.K. Bmp15 suppresses progesterone production by down-regulating star via alk3 in human granulosa cells. Mol. Endocrinol. 2013, 27, 2093–2104. [Google Scholar] [CrossRef]
- Shimizu, K.; Nakamura, T.; Bayasula; Nakanishi, N.; Kasahara, Y.; Nagai, T.; Murase, T.; Osuka, S.; Goto, M.; Iwase, A.; et al. Molecular mechanism of fshr expression induced by bmp15 in human granulosa cells. J. Assist. Reprod. Genet. 2019, 36, 1185–1194. [Google Scholar] [CrossRef]
- Zhou, S.; Wen, S.; Sheng, Y.; Yang, M.; Shen, X.; Chen, Y.; Kang, D.; Xu, L. Association of Estrogen Receptor Genes Polymorphisms with Polycystic Ovary Syndrome: A Systematic Review and Meta-Analysis Based on Observational Studies. Front. Endocrinol. 2021, 12, 726184. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Forward Primer (5′→3′) | Reverse Primer (5′→3′) |
|---|---|---|
| BAX | CCAAGAAGCTGAGCGAGTGTCTG | GTGTCCACGGCTGCGATCATC |
| BCL2 | TGTGGATGACCGAGTACCTGAACC | GCCAGACTGAGCAGTGCCTTC |
| Caspase3 | ATACCAGTTGAGGCAGAC | TTAACCCGAGTAAGAATGT |
| PCNA | GTAGCCGTGTCATTGCGACTCC | GCTCTGTAGGTTCACGCCACTTG |
| StAR | GCGACCAAGAGCTTGCCTATATCC | TTGGCCTGCCGACTCTCCTTC |
| CYP19A1 | AGGTCATCCTGGTCACCCTTCTG | CGGTCGCTGGTCTCGTCTGG |
| 3β-HSD | CTCAGACGACACACCACACCAAAG | CAGCAGGAAGGCAAGCCAGTAC |
| CYP11A1 | GCTGCGGAAGGAGGTTCTGAATG | GCACCAGTGTCTTGGCAGGAATC |
| CDK4 | GCTGCTGCTGGAGATGCTGAC | CTCTGCGTCACCTTCTGCCTTG |
| CCND1 | TTCCTCTCCTATCACCGCCTGAC | TCCTCTCTTCCTCCTCCTCCTC |
| CCNE1 | AAGTGCTCCTGCCTCAGTATCCTC | ATACAAGGCGGAAGCAGCAAGTAC |
| ESR1 | CTGCTGCTGGAGATGCTGGATG | GCTGGCTCTGATTCACGTCTTCC |
| LHR | ATTCCGCCATCTTTGCTGAGAGTG | AGCATCTGGTTCAGGAGCACATTG |
| FSHR | TTTGTGGTCATCTGTGGCTGCTAC | CGCTTGGCTATCTTGGTGTCACTAG |
| β-actin | TGATATTGCTGCGCTCGTGGT | GTCAGGATGCCTCTCTTGCTC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, S.; Ding, W.; Yao, H.; Wang, L.; Li, B.; Wang, Y.; Tang, X.; Zhang, Y.; Huang, D.; Xu, D.; et al. BMP6 Promotes the Secretion of 17 Beta-Estradiol and Progesterone in Goat Ovarian Granulosa Cells. Animals 2022, 12, 2132. https://doi.org/10.3390/ani12162132
Song S, Ding W, Yao H, Wang L, Li B, Wang Y, Tang X, Zhang Y, Huang D, Xu D, et al. BMP6 Promotes the Secretion of 17 Beta-Estradiol and Progesterone in Goat Ovarian Granulosa Cells. Animals. 2022; 12(16):2132. https://doi.org/10.3390/ani12162132
Chicago/Turabian StyleSong, Shuaifei, Wenfei Ding, Hui Yao, Lei Wang, Bijun Li, Yukun Wang, Xue Tang, Yiyu Zhang, Deli Huang, Dejun Xu, and et al. 2022. "BMP6 Promotes the Secretion of 17 Beta-Estradiol and Progesterone in Goat Ovarian Granulosa Cells" Animals 12, no. 16: 2132. https://doi.org/10.3390/ani12162132