Anabolic Steroids in Fattening Food-Producing Animals—A Review


{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Legislative Measures for the Use of AAS
2.1. History of the Use of Anabolic Steroids
2.2. Current Situation in Europe
2.3. Current Situation in the Rest of the World
3. The Most Commonly Used or Misused Anabolic Steroids
3.1. Boldenon
3.2. Chlortestosteron
3.3. Nandrolon
3.4. Stanozolol
3.5. Trenbolon
4. Effect of Anabolic Steroids on Tissues
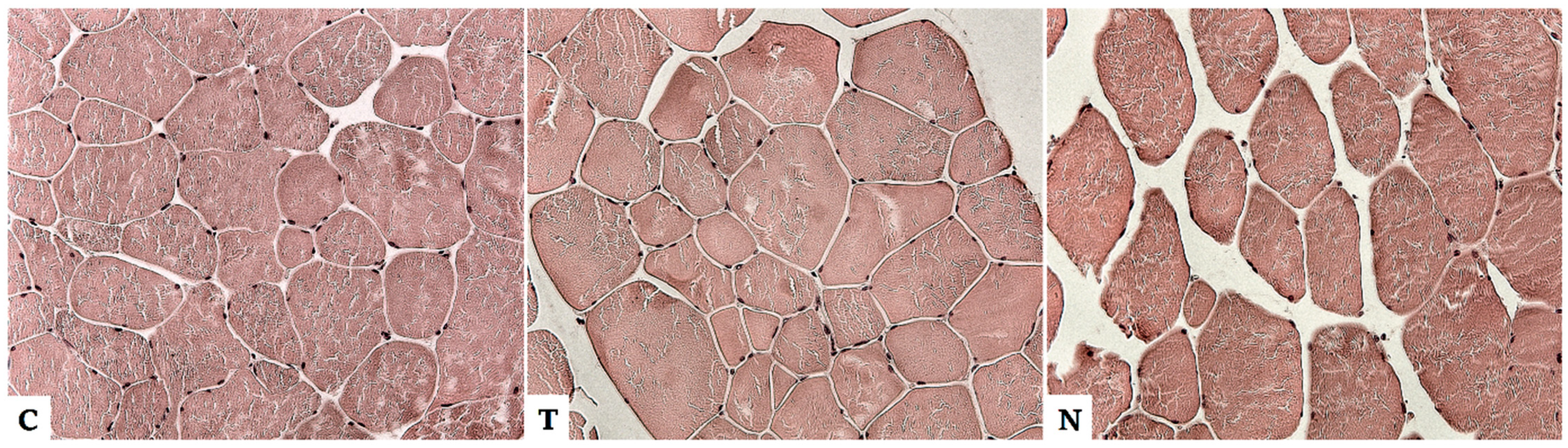
4.1. Effect of Anabolic Steroids on Skeletal Muscle
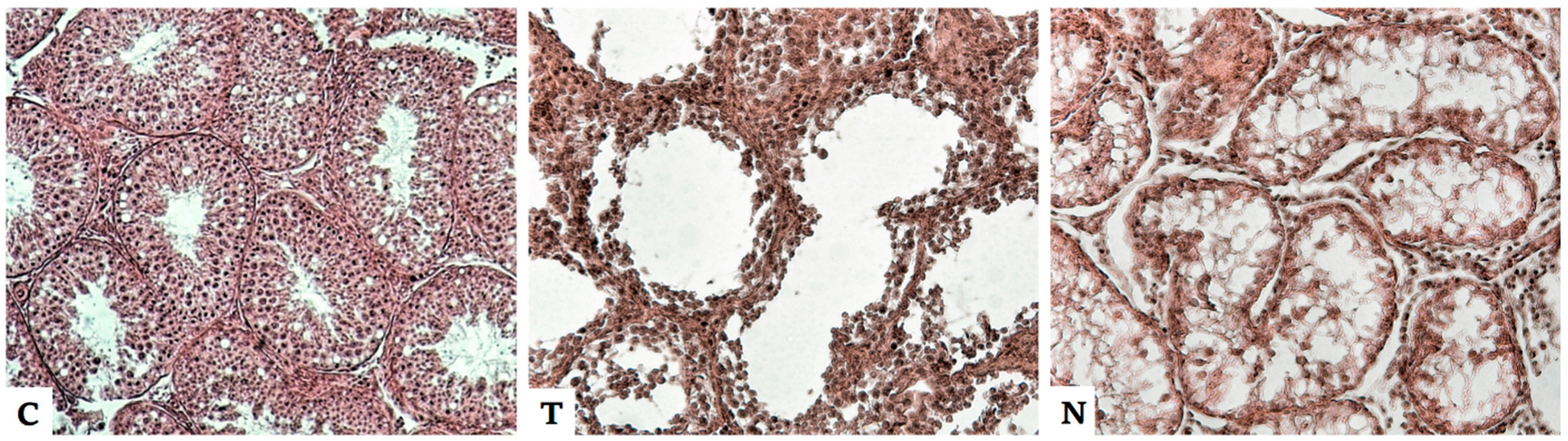
4.2. Effect of Anabolic Steroids on the Male Reproductive System
4.3. Effect of Anabolic Steroids on Other Tissues
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Medvei, V.C. A History of Endocrinology; MTP Press: Hingham, MA, USA, 1982. [Google Scholar]
- Cole, T.J.; Short, K.L.; Hooper, S.B. The science of steroids. Semin. Fetal Neonatal Med. 2019, 24, 170–175. [Google Scholar] [CrossRef] [PubMed]
- Echeverria, P.C.; Picard, D. Molecular chaperones, essential partners of steroid hormone receptors for activity and mobility. Biochim. Biophys. Acta Mol. Cell Res. 2010, 1803, 641–649. [Google Scholar] [CrossRef] [PubMed]
- Lösel, R.; Wehling, M. Nongenomic actions of steroid hormones. Nat. Rev. Mol. Cell Biol. 2003, 4, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, C.M. Anabolic Steroids. Recent Prog. Horm. Res. 2002, 57, 411–434. [Google Scholar] [CrossRef] [PubMed]
- Kreutzer, K.V.; Turk, J.R.; Casteel, S.W. Clinical Biochemistry in Toxicology. In Clinical Biochemistry of Domestic Animals, 6th ed.; Kaneko, J.J., Harvey, J.W., Bruss, M.L., Eds.; Academic Press: Cambridge, MA, USA, 2008; pp. 821–838. [Google Scholar]
- Johnson, B.J.; White, M.E.; Hathaway, M.R.; Christians, C.J.; Dayton, W.R. Effect of a combined trenbolone acetate and estradiol implant on steady-state IGF-I mRNA con-centrations in the liver of wethers and the longissimus muscle of steers. J. Anim. Sci. 1998, 76, 491–497. [Google Scholar] [CrossRef] [PubMed]
- Becker, C.h.; Riedmaier, I.; Reiter, M.; Tichopad, A.; Pflaffl, M.W.; Meyer, H.H.D. Effect of trenbolone acetate plus estradiol on transcriptional regulation of metabolism path-ways in bovine liver. Horm. Mol. Biol. Clin. Investig. 2010, 2, 257–265. [Google Scholar]
- Smith, Z.K.; Johnson, B.J. Mechanisms of steroidal implants to improve beef cattle growth: A review. J. Appl. Anim. Res. 2020, 48, 133–141. [Google Scholar] [CrossRef]
- Yoshida, E.M.; Erb, S.R.; Scudamore, C.H.; Owen, D.A. Severe Cholestasis and Jaundice Secondary to an Esterified Testosterone, a Non-C17 Alkylated Anabolic Steroid. J. Clin. Gastroenterol. 1994, 18, 268–269. [Google Scholar] [CrossRef]
- Webb, M.J.; Pendell, D.L.; Harty, A.A.; Salverson, R.R.; Rotz, C.A.; Underwood, K.R.; Olson, K.C.; Blair, A.D. Influence of Growth Promoting Technologies on Animal Per-formance, Production, Economics, Environmental Impacts and Carcass Characteristics of Beef. Meat Muscle Biol. 2017, 1, 23–24. [Google Scholar] [CrossRef]
- Capper, J.L.; De Carvalho, T.B.; Hancock, A.S.; Filho, O.G.S.; Odeyemi, I.; Bartram, D.J. Modeling the effects of steroid implant use on the environmental and economic sus-tainability of Brazilian beef production. Transl. Anim. Sci. 2021, 5, txab144. [Google Scholar] [CrossRef] [PubMed]
- Capper, J.L. The environmental and economic impact of steroid implant and be-ta-adrenergic agonist use within U.S. beef production. In Proceedings of the ADSA-ASAS Joint Annual Meeting, Indianapolis, IN, USA, 8–12 July January 2013. [Google Scholar]
- Dotson, J.L.; Brown, R.T. The History of the Development of Anabolic-Androgenic Steroids. Pediatric Clin. N. Am. 2007, 54, 761–769. [Google Scholar] [CrossRef] [PubMed]
- Fourcroy, J. History of androgens and anabolic steroids: Use, abuse, and identification. J. Urol. 2010, 183, e433. [Google Scholar] [CrossRef]
- Zhao, S.; Zhu, W.; Xue, S.; Han, D. Testicular defense systems: Immune privilege and innate immunity. Cell. Mol. Immunol. 2014, 11, 428–437. [Google Scholar] [CrossRef] [PubMed]
- Nieschlag, E.; Nieschlag, S. Testosterone deficiency: A historical perspective. Asian J. Androl. 2014, 16, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Soma, K.K. Testosterone and Aggression: Berthold, Birds and Beyond. J. Neuroendocrinol. 2006, 18, 543–551. [Google Scholar] [CrossRef] [PubMed]
- Lukas, S.E. Current perspectives on anabolic-androgenic steroid abuse. Trends Pharmacol. Sci. 1993, 14, 61–68. [Google Scholar] [CrossRef]
- Dinusson, W.E.; Andrews, F.N.; Beenson, W.M. The effects of stilbestrol, testosterone, thyroid alteration and spaying on the growth and fattening of beef heifers. J. Anim. Sci. 1950, 9, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Ronquillo, M.G.; Hernandez, J.C.A. Antibiotic and synthetic growth promoters in animal diets: Review of impact and analytical methods. Food Control. 2017, 72, 255–267. [Google Scholar] [CrossRef]
- Yeh, S.; Lovitt, S.; Schuster, M.W. Usage of megestrol acetate in the treatment of ano-rexia-cachexia syndrome in the elderly. J. Nutr. Health Aging 2009, 13, 448–454. [Google Scholar] [CrossRef]
- Verbeke, R. Senstitive multi-residue method for detection of anabolics in urine and in tissues of slaughtered animals. J. Chromatogr. A 1979, 177, 69–84. [Google Scholar] [CrossRef]
- Brunetti, A.; Manfioletti, G. Hormone receptorsand breast cancer. Front. Endocrinol. 2019, 10, 205. [Google Scholar] [CrossRef] [PubMed]
- Reig, M.; Toldrá, F. Veterinary drug residuesin meat: Concerns and rapid methods for detection. Meat Sci. 2008, 78, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Official Journal of the European Union, L12523/05/1996. Council Directive 96/22/EC of 29 April 1996 Concerning the Prohibition on the Use in Stockfarming of Certain Substances Having a Hormonal or Thyrostatic Action and of Beta-Agonists, and Repealing Directives 81/602/EEC, 88/146/EEC and 88/299/EEC; European Commission: Brussels, Belgium, 1996. [Google Scholar]
- Official Journal of the European Union, L125, 23/05/1996. Council Directive 96/23/EC of 29 April 1996 on Measures to Monitor Certain Substances and Residues Thereof in Live Animals and Animal Products and Repealing Directives 85/358/EEC and 86/469/EEC and Decision 89/187/EEC and 91/664/EEC; European Commission: Brussels, Belgium, 1996. [Google Scholar]
- Official Journal of the European Union, L 95/1. Council Regulation (EU) 2017/625 Of The European Parliament And Of The Council of 15 March 2017 on Official Controls and Other Official Activities Performed to Ensure the Application of Food and Feed, Law, Rules on Animal Health and Welfare, Plant Health and Plant Protection Products, Amending; European Commission: Brussels, Belgium, 2017. [Google Scholar]
- Nachman, K.E.; Smith, T.J. Hormone use in food animal production:assessing potential dietary exposures and breast cancer risk. Curr. Environ. Health Rep. 2015, 2, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Official Journal of the European Union, L22418 August 1990. Council Regulation 2377/90/EC of 26 June 1990 Laying Down a Community Procedure for the Establishment of Maximum Residue Limits of Veterinary Medicinal Products in Foodstuffs of Animal Origin; European Commission: Brussels, Belgium, 1990. [Google Scholar]
- Official Journal of the European Communities L221. Commission Decision (2002/657/EC) of 12 August 2002; European Commission: Brussels, Belgium, 2002. [Google Scholar]
- Official Journal of the European Union, L 180/84. Commission Implementing Regulation (EU) 2021/808 of 22 March 2021 on the Performance of Analytical Methods for Residues of Pharmacologically Active Substances Used in Food-Producing Animals and on the Interpretation of Results as well as on the Methods to be Used for Sampling and Repealing Decisions 2002/657/EC and 98/179/EC; European Commission: Brussels, Belgium, 2021. [Google Scholar]
- Official Journal of the European Union, L 262/17. Directive 2003/74/EC of The Europe-An Parliament and of the Council of 22 September 2003 Amending Council Directive 96/22/EC Concerning the Prohibition on the Use in Stockfarming of Certain Substances Having a Hormonal or Thyrostatic Action and of Beta-Agonists; European Commission: Brussels, Belgium, 2003. [Google Scholar]
- Karg, H.; Vogt, K. Control of Hormone Treatment in Animals and Residues in Meat—Regulatory Aspects and Approaches in Methodology. J. Assoc. Off. Anal. Chem. 2020, 61, 1201–1208. [Google Scholar] [CrossRef]
- National Research Council (US) Committee to Study the Human Health Effects of Subtherapeutic Antibiotic Use in Animal Feeds. The Effects on Human Health of Sub-Therapeutic Use of Antimicrobials in Animal Feeds; National Academies Press (US): Washington, DC, USA, 1980. [Google Scholar]
- FOOD AND DRUG ADMINISTRATION: Steroid Hormone Implants Used for Growth in Food-Producing Animals [Online]. Available online: https://www.fda.gov/animal-veterinary/product-safety-information/steroid-hormone-implants-used-growth-food-producing-animals (accessed on 13 March 2022).
- Stackhouse, K.R.; Rotz, C.A.; Oltjen, J.W.; Mitloehner, F.M. Growth promoting technologies reduce the carbon footprint, ammonia emissions, and costs of California beef production systems. J. Anim. Sci. 2012, 90, 4656–4665. [Google Scholar] [CrossRef] [PubMed]
- Webb, M.J.; Block, J.J.; Harty, A.A.; Salverson, R.R.; Daly, R.F.; Jaeger, J.R.; Underwood, K.R.; Funston, R.N.; Pendell, D.P.; Rotz, C.A.; et al. Cattle and carcass performance, and life cycle assessment of production systems utilizing additive combinations of growth promotant technologies. Transl. Anim. Sci. 2020, 4, txaa216. [Google Scholar] [CrossRef]
- CANADIAN ANIMAL HEALTH INSTITUTE: Hormones [Online]. Available online: https://www.cahi-icsa.ca/hormones (accessed on 14 March 2022).
- Sundlof, S.F. Drug and chemical residues in livestock. Vet. Clin. N. Am. Food Anim. Pract. 1989, 5, 411–449. [Google Scholar] [CrossRef]
- Yongmin, B. The Challenges for Food Safety in China. China Perspect. 2004, 53. [Google Scholar] [CrossRef]
- Zeng, Z.; Yang, F.; Wang, L. Veterinary Drug Residues in China: Science, Technology, Management and Regulation. In Food Safety in China; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2017; pp. 219–235. [Google Scholar]
- MOA. Regulation on Administration of Veterinary Drugs. 2004. Available online: http://www.gov.cn/gongbao/content/2004/content_62760.htm (accessed on 26 April 2022).
- MOA. Measures for Registration of Veterinary Drugs. 2012. Available online: http://www.moa.gov.cn/fwllm/zxbs/xzxk/bszl/syj/201204/t20120426_2612402.htm (accessed on 13 March 2022).
- Wu, H.; Weng, C. Certified Veterinarian and Certified Veterinarian System. Chin. J. Vet. Drug 2010, 55, 29–33. [Google Scholar]
- Wang, X.; Liu, Y.; Su, Y.; Yang, J.; Bian, K.; Wang, Z.; He, L. High-throughput screening and confirmation of 22 banned veterinary drugs in feedstuffs using LC-MS/MS and high-resolution Orbitrap mass spectrometry. J. Agric. Food Chem. 2014, 62, 516–527. [Google Scholar] [CrossRef]
- Passantino, A. Steroid Hormones in Food Producing Animals. In A Bird’s-Eye View of Veterinary Medicine; Perez-Marin, C.C., Ed.; InTech: Córdoba, Spain, 2012; ISBN 978-953-51-0031-7. [Google Scholar]
- Aslam, M.H.; Hashem, M.A.; Hossain, M.M.; Islam, M.S.; Rana, M.S.; Habibullah, M. Present status on the use of anabolic steroids and feed additives in small scale cattle fattening in Bangladesh. Progress. Agric. 2012, 23, 1–13. [Google Scholar] [CrossRef]
- Al-Amri, I.; Kadim, I.T.; Alkindi, A.; Hamaed, A.; Al-Magbali, R.; Khalaf, S.; Al-Hosni, K.; Mabood, F. Determination of residues of pesticides, anabolic steroids, antibiotics, and antibacterial compounds in meat products in Oman by liquid chromatography/mass spectrometry and enzyme-linked immunosorbent assay. Vet. World 2021, 14, 709–720. [Google Scholar] [CrossRef] [PubMed]
- APVMA. Australian Pesticides and Veterinary Medicines Authority. Substances Not Permitted for Use on Food-Producing Animals in Australia. Available online: https://apvma.gov.au/node/11626 (accessed on 30 April 2022).
- Aroeira, C.N.; Feddern, V.; Gressler, V.; Contreras-Castillo, C.F.; Hopkins, D.L. Growth Promoters in Cattle and Pigs: A Review of Legislation and Implications for Human Health. Food Rev. Int. 2021, 38, 1–23. [Google Scholar] [CrossRef]
- Patrick, S.M.; Aneck-Hahn, N.H.; Wyk, S.V.; Van Zilj, M.C.; Huma, M.; Jager, C.H. Veterinary growth promoters in cattle feedlot runoff: Estrogenic activity and potential effects on the rat male reproductive systém. Environ. Sci. Pollut. Res. 2020, 27, 13939–13948. [Google Scholar] [CrossRef] [PubMed]
- Mitema, E.S. Improved management of drugs, hormones and pesticides in Africa. Onderstepoort J. Vet. Res. 2009, 76, 155–159. [Google Scholar] [CrossRef]
- De Brabander, H.F.; Poelmans, S.; Schilt, R.; Stephany, R.W.; Le Bizec, B.; Draisci, R.; Sterk, S.S.; Ginkel, L.A.; Courtheyn, D.; Van Hoof, N.; et al. Presence and metabolism of the anabolic steroid boldenone in various animal species: A review. Food Addit. Contam. 2004, 21, 515–525. [Google Scholar] [CrossRef]
- Behairy, A.; Mohamed, W.A.M.; Ebraheim, L.L.M.; Soliman, M.M.; Abd-Elhakim, Y.M.; El-Sharkawy, N.I.; Saber, T.M.; El Deib, M.M. Boldenone Undecylenate-Mediated Hepatorenal Impairment by Oxidative Damage and Dysregulation of Heat Shock Protein 90 and Androgen Receptors Expressions: Vitamin C Preventive Role. Front. Pharmacol. 2021, 12, 651497. [Google Scholar] [CrossRef]
- Saber, T.M.; Omran, B.H.F.; El Deib, M.M.; El-Sharkawy, N.I.; Metwally, M.M.M.; Abd-Elhakim, Y.M. Early postmortem biochemical, histological, and immunohistochemical alterations in skeletal muscles of rats exposed to boldenone undecylenate: Forensic implication. J. Forensic Leg. Med. 2021, 83, 102248. [Google Scholar] [CrossRef]
- Elmajdoub, A.; Garbaj, A.; Abolghait, S.; El-Mahmoudy, A. Evaluation of boldenone as a growth promoter in broilers: Safety and meat quality aspects. J. Food Drug Anal. 2016, 24, 284–292. [Google Scholar] [CrossRef]
- Aly, M.A.S.; El-Shamarka, M.E.; Soliman, T.N.; Elgabry, M.A.E. Protective effect of nanoencapsulated curcumin against boldenone-induced testicular toxicity and oxidative stress in male albino rats. Egypt. Pharm. J. 2021, 20, 72–81. [Google Scholar]
- Rossi, C.A.S.; Arioli, F.; Bassini, A.; Chiesa, L.M.; Dell’orto, V.; Montana, M.; Pompa, G. Evidence for false-positive results for boldenone testing of veal urine due to faecal cross-contamination during sampling. Food Addit. Contam. 2004, 21, 756–762. [Google Scholar]
- Groot, M.J.; Lasaroms, J.J.P.; Bennekom, E.O.; Meijer, T.; Vinyeta, E.; Klis, J.D.; Nielen, M.W. Illegal treatment of barrows with nandrolone ester: Effect on growth, histology and residue levels in urine and hair. Food Addit. Contam. Part A 2012, 29, 727–735. [Google Scholar] [CrossRef]
- De Wasch, K.; Poelmans, S.; Verslycke, T.; Janssen, C.; Van Hoof, N.; De Brabander, H.F. Alternative to vertebrate animal experiments in the study of metabolism of illegal growth promotors and veterinary drugs. Anal. Chim. Acta 2002, 473, 59–69. [Google Scholar] [CrossRef]
- Gallina, G.; Ferretti, G.; Merlanti, R.; Civitareale, C.; Capolongo, F.; Draisci, R.; Montesissa, C. Boldenone, Boldione, and Milk Replacers in the Diet of Veal Calves: The Effects of Phytosterol Content on the Urinary Excretion of Boldenone Metabolites. J. Agric. Food Chem. 2007, 55, 8275–8283. [Google Scholar] [CrossRef] [PubMed]
- Verheyden, K.; Noppe, H.; Zorn, H.; Van Immerssel, F.; Bussche, J.V.; Wille, K.; Bekaert, K.; Janssen, C.R.; De Brabander, H.F.; Vanhaecke, L. Endogenous boldenone-formation in cattle: Alternative invertebrate organisms to elucidate the enzymatic pathway and the potential role of edible fungi on cattle’s feed. J. Steroid Biochem. Mol. Biol. 2010, 119, 161–170. [Google Scholar] [CrossRef]
- Nielen, M.W.F.; Rutgers, P.; Bennekom, E.O.; Lasaroms, J.J.P.; Rhinj, J.A. Confirmatory analysis of 17β-boldenone, 17α-boldenone and androsta-1,4-diene-3,17-dione in bovine urine, faeces, feed and skin swab samples by liquid chromatography–electrospray ionisation tandem mass spektrometry. J. Chromatogr. B 2004, 801, 273–283. [Google Scholar] [CrossRef]
- Viljanto, M.; Kaabia, Z.; Taylor, P.; Muir, T.; Habershon-butcher, J.; Bailly-Chouriberry, L.; Scarth, J. Differentiation of boldenone administration from ex vivo transformation in the urine of castrated male horses. Drug Test. Anal. 2022, 14, 887–901. [Google Scholar] [CrossRef] [PubMed]
- Decloedt, A.; Van Landschoot, A.; Vanhaecke, L. Mass Spectrometry for the Detection of Endogenous Steroids and Steroid Abuse in (Race) Horses and Human Athletes. In Mass Spectrometry; InTech: London, UK, 2016. [Google Scholar] [CrossRef]
- Leund, G.N.W.; Ho, E.N.M.; Leung, D.K.K.; Tang, F.P.W.; Wan, T.S.M.; Yeung, J.H.K.; Wong, H.N.C. Metabolic Studies of Clostebol Acetate in Horses. Chromatographia 2005, 61, 397. [Google Scholar]
- Rahnema, C.D.; Crosnoe, L.E.; Kim, E.D. Designer steroids–over-the-counter supplements and their androgenic component: Review of an increasing problem. Andrology 2015, 3, 150–155. [Google Scholar] [CrossRef]
- Leyssens, L.; Royackers, E.; Gielen, B.; Missotten, M.; Schoofs, J.; Czech, J.; Noben, J.P.; Hendriks, L.; Raus, J. Metabolites of 4-chlorotestosterone acetate in cattle urine as diagnostic markers for its illegal use. J. Chromatogr. B 1994, 654, 43–54. [Google Scholar] [CrossRef]
- Le Bizec, B.; Montrade, M.; Monteau, F.; Gaudin, I.; Andre, F. 4-Chlorotestosterone acetate metabolites in cattle after intramuscular and oral administrations. Clin. Chem. 1998, 44, 973–984. [Google Scholar] [CrossRef] [PubMed]
- Crabbe, P.; Meyer, U.J.; Zhi, Z.; Pieraccini, G.; O’keeffe, M.; Van Peteghem, C. Screening of Clostebol and its Metabolites in Bovine Urine with ELISA and Comparison with GC-MS Results in an Interlaboratory Study. J. Anal. Toxicol. 2003, 27, 213–220. [Google Scholar] [CrossRef]
- Jiafeng, Y.; Decheng, S.; Xiaoyong, L.; Guangyu, L.; Min, B.S. Multiresidue determina-tion of 19 anabolic steroids in animal oil using enhanced matrix removal lipid cleanup and ultrahigh performance liquid chromatography-tandem mass spectrometry. Anal. Ical Methods 2021, 13, 2374–2383. [Google Scholar] [CrossRef] [PubMed]
- Pan, M.M.; Kovac, J.R. Beyond testosterone cypionate: Evidence behind the use of nandrolone in male health and wellness. Transl. Androl. Urol. 2016, 5, 213–219. [Google Scholar] [CrossRef]
- Sauer, M.J.; Samuels, T.P.W.; Howells, L.G.; Seymour, M.A.; Nedderman, A.; Houghton, E.; Bellworthy, S.J.; Andersons, S.; Coldham, N.G. Residues and metabolism of 19-nortestosterone laureate in steers. Analyst 1998, 123, 2653–2660. [Google Scholar] [CrossRef]
- Rosegger, J.; Schmerold, I.; Ahmed, S.; Schuch, R.; Eppinger, G.; Steiner, S.; Baumgartner, W.; Armstrong, H.; Schauberger, G.; Mcevoy, J.D.G.; et al. Natural occurrence and elimination of 19-nortestosterone in sheep: Pregnant ewes, male and female lambs before and after treatment. Vet. Med. Austria 2009, 96, 171–183. [Google Scholar]
- Houghton, E.; Dumasia, M.C. Studies related to the metabolism of anabolic steroids in the horse: The identification of some 16-oxygenated metabolites of 19- nortestosterone. Xenobiotica 1980, 10, 381–390. [Google Scholar] [CrossRef] [PubMed]
- Dehennin, L.; Silberzahn, P.; Reiffsteck, A.; Zwain, I. 19-norandrostenedione and 19-nortestosterone in human and equine follicular fluid incidence on the accuracy of radioimmunoassay of some androgens. Pathol. Biol. 1984, 32, 828–829. [Google Scholar]
- Poelmans, S.; De Wasch, K.; Noppe, H.; Van Hoof, N.; Van Cruchten, S.; Le Bizec, B.; Deceuninck, Y.; Sterk, S.; Van Rossum, H.J.; Hoffman, M.K.; et al. Endogenous occurrence of some anabolic steroids in swine matrices. Food Addit. Contam. 2005, 22, 808–815. [Google Scholar] [CrossRef] [PubMed]
- Scarth, J.; Akre, C.; Ginkel, L.; Le Bizec, B.; De Brabander, H.; Korth, W.; Points, J.; Teale, P.; Kay, J. Presence and metabolism of endogenous androgenic-anabolic steroid hormones in meat-producing animals: A review. Food Addit. Contam. 2009, 26, 640–671. [Google Scholar] [CrossRef]
- Ouzia, S.; Royer, A.; Pezzolato, M.; Benedetto, A.; Biasibetti, E.; Guitton, Y.; Le Bizec, B.; Bozetta, E.; Dervilly, G. Nandrolone and estradiol biomarkers identification in bovine urine applying a liquid chromatography high-resolution mass spectrometry metabolomics approach. Drug Testind Anal. 2021, 14, 879–886. [Google Scholar] [CrossRef] [PubMed]
- National Center for Biotechnology Information. PubChem Compound Summary for CID 25249, Stanozolol. 2022. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Stanozolol (accessed on 22 March 2022).
- Adamama-Moraitou, K.K.; Pardali, D.; Athanasiou, L.V.; Prassinos, N.N.; Kritsepi, M.; Rallis, T.S. Conservative Management of Canine Tracheal Collapse with Stanozolol: A Double Blinded, Placebo Control Clinical Trial. Int. J. Immunopathol. Pharmacol. 2011, 24, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Martins, M.C.; Peffers, M.J.; Lee, K.; Rubios-Martines, L.M. Effects of stanozolol on normal and IL-1β-stimulated equine chondrocytes in vitro. BMC Vet. Res. 2018, 14, 103. [Google Scholar]
- Salmani, S.; Rastabi, H.I.; Tabatabaei, S.R.F.; Rezaee, A.; Gooraninejad, S.; Mosallanejad, B. The Effects of Stanozolol and Nandrolone Decanolate Hormones on Erytropoetin and Testosteron Serum Concentrations in Dogs. Iran. J. Vet. Med. 2021, 15, 325–334. [Google Scholar]
- Ferchaud, V.; Bizec, B.; Montrade, M.P.; Maume, D.; Monteau, F.; André, F. Gas chromatographic–mass spectrometric identification of main metabolites of stanozolol in cattle after oral and subcutaneous administration. J. Chromatogr. B 1997, 695, 269–277. [Google Scholar] [CrossRef]
- Courtheyn, D.; Bizec, B.; Brambilla, G.; De Brabander, H.F.; Cobbaert, E.; Van De Wiele, M.; Vercammen, J.; Wasch, K. Recent developments in the use and abuse of growth promoters. Anal. Chim. Acta 2002, 473, 71–82. [Google Scholar] [CrossRef]
- Tsitsimpikou, C.h.; Tsarouhas, K.; Spandidos, D.A.; Tsatsakis, A.M. Detection of stanozolol in the urine of athletes at a pg level: The possibility of passive exposure. Biomed. Rep. 2016, 5, 665–666. [Google Scholar] [CrossRef] [PubMed]
- Poelmans, S.; Wasch, K.; De Brabander, H.F.; Wiele, M.V.; Courtheyn, D.; Ginkel, L.A.; Sterk, S.S.; Delehaut, P.; Dubois, M.; Schilt, R.; et al. Analytical possibilities for the detection of stanozolol and its metabolites. Anal. Chim. Acta 2002, 473, 39–47. [Google Scholar] [CrossRef]
- Post, L.O.; Bataller, N.; Parkhie, M.; Keller, W.C. Regulatory Toxicology. In Clinical Veterinary Toxicology; Mosby: Maryland Heights, MO, USA, 2004; pp. 28–45. [Google Scholar]
- Saičic, S.; Spiric, A.; Jankovic, S.; Dordevic, M. A trenbolone acetate/estradiol combination in feedlot simmental bulls: Meat quality and withdrawal time of trenbolone. Acta Vet. 2000, 50, 137–146. [Google Scholar]
- Kolok, A.S.; Ali, J.M.; Rogan, E.G.; Bartelt-Hunt, S.L. The Fate of Synthetic and Endogenous Hormones Used in the US Beef and Dairy Industries and the Potential for Human Exposure. Curr. Environ. Health Rep. 2018, 5, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Qaid, M.M.; Abdoun, K.A. Safety and concerns of hormonal application in farm animal production: A review. J. Appl. Anim. Res. 2022, 50, 426–439. [Google Scholar] [CrossRef]
- Munawaroh, I.S.; Rahayu, P. Determination of trenbolone acetate hormone residue on imported beef meat and imported beef liver at slaughterhouse and cold storage. In Proceedings of the International Seminar on Livestock Production and Veterinary Technology, Bogor, Indonesia, 6–7 September 2021; p. 31. [Google Scholar]
- Widiastuti, R.; Murdiati, T.B.; Yuningsih. Residu of 17-beta-trenbolon on imported calf meat and liver distributed in Jakarta (Indonesia). In Proceedings of the Seminar Nasional Peternakan dan Veteriner, Bogor, Indonesia, 18–19 September 2000. [Google Scholar]
- El Shahid, E.Y.M.A.; El Shater, M.A.; Hasan, M.A.; Ibrahim, M.H. Chemical residues in burger and sausage meat products. Benha Vet. Med. J. 2021, 40, 161–164. [Google Scholar]
- Hirpessa, B.B.; Ulusoy, B.H.; Hecer, C. Hormones and Hormonal Anabolics: Residues in Animal Source Food, Potential Public Health Impacts, and Methods of Analysis. J. Food Qual. 2020, 2020, 5065386. [Google Scholar] [CrossRef]
- Nazli, B.; Olgun, E.O.; Çakir, B.; Demirci, M. An analytical study to determine prohibited anabolic residues in red meat tissue using LC-MS/MS system. Food Sci. Technol. 2022, 42. [Google Scholar] [CrossRef]
- Pleadin, J.; Samardžija, M. Hormonally active substances in the food chain from farm animals to consumers. Vet. Stanica 2019, 50, 501–512. [Google Scholar]
- Benedetto, A.; Pezzolato, M.; Biasibetti, E.; Bozzetta, E. Omics applications in the fight against abuse of anabolic substances in cattle: Challenges, perspectives and opportunities. Curr. Opin. Food Sci. 2021, 40, 112–120. [Google Scholar] [CrossRef]
- Perry, T.C.; Fox, D.G.; Beermann, D.H. Effect of an implant of trenbolone acetate and estradiol on growth, feed efficiency, and carcass composition of Holstein and beef steers. J. Anim. Sci. 1992, 69, 4696–4702. [Google Scholar] [CrossRef] [PubMed]
- Clancy, M.J.; Lester, J.M.; Roche, J.F. The Effects of Anabolic Agents and Breed on the Fibers of the Longissimus Muscle of Male Cattle. J. Anim. Sci. 1986, 63, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Guyomarda, H.; Bouamra-Mechemacheb, Z.; Chatellierc, V.; Delabyd, L.; Dé-tang-Dessendree, C.; Peyraudf, J.L.; Réquillartb, V. Review: Why and how to regulate animal production and consumption: The case of the European Union. Animal 2021, 15, 100283. [Google Scholar] [CrossRef] [PubMed]
- Kellermeier, J.D.; Tittor, A.W.; Brooks, J.C.; Galyean, M.L.; Yates, D.A.; Hutcheson, J.P.; Nichols, W.T.; Streeter, M.N.; Johnson, B.J.; Miller, M.F. Effects of zilpaterol hydrochloride with or without an estrogen-trenbolone acetate terminal implant on carcass traits, retail cutout, tenderness, and muscle fiber diameter in finishing steers. J. Anim. Sci. 2009, 87, 3702–3711. [Google Scholar] [CrossRef] [PubMed]
- Fontana, K.; Campos, G.E.R.; Staron, R.S.; Cruz-Höfling, M.A. Effects of Anabolic Steroids and High-Intensity Aerobic Exercise on Skeletal Muscle of Transgenic Mice. PLoS ONE. 2013, 8, e80909. [Google Scholar] [CrossRef] [PubMed]
- Elgendy, H.; Alhawary, A.; El-Shahat, M.; Ali, A. Effect of Anabolic Steroids on the Cardiac and Skeletal Muscles of Adult Male Rats. Int. J. Clin. Dev. Anat. 2018, 4, 1–14. [Google Scholar] [CrossRef]
- Reichhardt, C.C.; Ahmadpour, A.; Christensen, R.G.; Ineck, N.E.; Murdoch, G.K.; Thornton, K.J. Understanding the influence of trenbolone acetate and polyamines on proliferation of bovine satellite cells. Domest. Anim. Endocrinol. 2021, 74, 106479. [Google Scholar] [CrossRef] [PubMed]
- Velders, M.; Diel, P. How Sex Hormones Promote Skeletal Muscle Regeneration. Sports Med. 2013, 43, 1089–1100. [Google Scholar] [CrossRef] [PubMed]
- Carson, J.A.; Manolagas, S.C. Effects of sex steroids on bones and muscles: Similarities, parallels, and putative interactions in health and disease. Bone 2015, 80, 67–78. [Google Scholar] [CrossRef]
- Kim, J.T.; Roberts, K.; Dunlap, G.; Perry, R.; Washington, T.; Wolchok, J.C. Nandrolone supplementation does not improve functional recovery in an aged animal model of volumetric muscle loss injury. J. Tissue Eng. Regen. Med. 2022, 16, 367–379. [Google Scholar] [CrossRef]
- Snijders, T.; Aussieker, T.; Holwerda, A.; Parise, G.; Van Loon, L.J.C.; Verdijk, L.B. The concept of skeletal muscle memory: Evidence from animal and human studies. Acta Physiol. 2020, 229, e13465. [Google Scholar] [CrossRef] [PubMed]
- Egner, I.M.; Brussgaard, J.C.; Eftestøl, E.; Gundersen, K. A cellular memory mechanism aids overload hypertrophy in muscle long after an episodic exposure to anabolic steroids. J. Physiol. 2013, 591, 6221–6230. [Google Scholar] [CrossRef] [PubMed]
- Hijazi, M.M.; Azmi, M.A.; Hussain, A.; Naqvi, S.N.H.; Perveen, R.; Hijazi, S. Androgenic Anabolic Steroidal-Based Effects on the Morphology of Testicular Structures of Albino Rats. Pak. J. Zool. 2012, 44, 1529–1537. [Google Scholar]
- Tousson, E.; El-Moghazy, M.; Massoud, A.; Akel, A. Histopathological and Immunohistochemical Changes in the Testes of Rabbits After Injection With the Growth Promoter Boldenone. Reprod. Sci. 2012, 19, 253–259. [Google Scholar] [CrossRef]
- Kahal, A.; Allem, R. Reversible effects of anabolic steroid abuse on cyto-architectures of the heart, kidneys and testis in adult male mice. Biomed. Pharmacother. 2018, 106, 917–922. [Google Scholar] [CrossRef]
- Yarrow, J.F.; Conover, C.F.; McCoy, S.C.; Lipinska, J.A.; Santillana, C.A.; Hance, J.M.; Cannady, D.F.; VanPelt, T.D.; Sanchez, J.; Conrad, B.P.; et al. 17β-Hydroxyestra-4,9,11-trien-3-one (trenbolone) exhibits tissue selective anabolic activity: Effects on muscle, bone, adiposity, hemoglobin, and prostate. Am. J. Physiol. Endocrinol. Metab. 2011, 300, E650–E660. [Google Scholar] [CrossRef]
- Salerno, M.; Cascio, O.; Bertozzi, G.; Sessa, F.; Messina, A.; Monda, V.; Cipolloni, L.; Biondi, A.; Daniele, A.; Pomara, C. Anabolic androgenic steroids and carcinogenicity focusing on Leydig cell: A literature review. Oncotarget 2018, 9, 19415–19426. [Google Scholar] [CrossRef]
- Kumar, P.; Lindberg, L.; Thirkill, T.L.; Ji, J.W.; Martsching, L.; Douglas, G.C. The MUC1 Extracellular Domain Subunit Is Found in Nuclear Speckles and Associates with Spliceosomes. PLoS ONE. 2012, 7, e42712. [Google Scholar] [CrossRef]
- Seara, F.A.C.; Barbosa, R.A.Q.; Oliveira, D.F.; Silva, D.L.S.G.; Carvalho, A.B.; Ferreira, A.C.F.; Nascimento, J.H.M.; Olivares, E.L. Administration of anabolic steroid during adolescence induces long-term cardiac hypertrophy and increases susceptibility to ischemia/reperfusion injury in adult Wistar rats. Mol. Biol. 2017, 171, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Aljeboori, K.H.; Majhool, A.B. Pathological and Immunological changes induced in male rats treated with therapeutic doses of sustanon. Al-Anbar J. Vet. Sci. 2017, 10, 52–57. [Google Scholar]
- Mohammed, A.R.S.; Al-Galad, G.M.; Abd-Elgayd, A.A.; Mwaheb, M.A.; Al-Hamboly, H.M. Effect of Nandrolone Decanoate (Anabolic Steroid) on the Liver and Kidney of Male Albino Rats and the Role of Antioxidant (Antox-Silymarin) as Adjuvant Therapy. J. Drug Metab. Toxicol. 2017, 8, 224. [Google Scholar] [CrossRef]
- Cho, J.; Izumi, K.; Huang, C.h.K. Androgen and Androgen Receptor in Kidney Cancer. Nephrol. Open J. 2015, 1, e7–e8. [Google Scholar] [CrossRef]
- Hartung, R.; Gerth, J.; Fünfstück, R.; Gröne, H.J.; Stein, G. End-stage renal disease in a bodybuilder: A multifactorial process or simply doping? Nephrol. Dial. Transplant. 2001, 16, 163–165. [Google Scholar] [CrossRef]
- Navarro, V.J.; Khan, I.; Björnsson, E.; Seeff, L.B.; Serrano, J.; Hoofnagle, J.H. Liver injury from herbal and dietary supplements. Hepatology 2017, 65, 363–373. [Google Scholar] [CrossRef]
- Guimarães, A.P.F.G.M.; Butezloff, M.M.; Zamarioli, A.; Assa, J.P.M.; Volpon, J.B. Nandrolone decanoate appears to increase bone callus formation in young adult rats after a complete femoral fracture. Acta Cirúrgica Bras. 2017, 32, 924–934. [Google Scholar] [CrossRef] [PubMed]
- Souza, D.B.; Brasil, F.B.; Marchon, R.G.; Félix-Patrício, B. Effects of nandrolone decanoate on femur morphology. Experimental study. Acta Cirúrgica Bras. 2021, 36, e360507. [Google Scholar] [CrossRef]
- Marchi, P.N.; Sueur Vieira, A.N.L.; Ribeiro, J.F.A.; Geraldes, S.S.G.; Ramos, P.R.R.; Malchert, A.; Guimarães-Okamoto, P.T.C. Use of Nandrolone Decanoate in Treatment of Pure Red Cell Aplasia Secondary to Diclofenac Administration: A Case Report. Top. Companion Anim. Med. 2017, 32, 44–47. [Google Scholar] [CrossRef] [PubMed]
- Spadari, A.; Romagnoli, N.; Predieri, P.G.; Borghetti, P.; Cantoni, A.M.; Corradi, A. Effects of intraarticular treatment with stanozolol on synovial membrane and cartilage in an ovine model of osteoarthritis. Res. Vet. Sci. 2013, 3, 379–387. [Google Scholar] [CrossRef] [PubMed]
- Falanga, V.; Greenberg, A.S.; Zhou, L.; Ochoa, S.M.; Roberts, A.B.; Falabella, A.; Yamaguchi, Y. Stimulation of Collagen Synthesis by the Anabolic Steroid Stanozolol. J. Investig. Dermatol. 1998, 6, 1193–1197. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Skoupá, K.; Šťastný, K.; Sládek, Z. Anabolic Steroids in Fattening Food-Producing Animals—A Review. Animals 2022, 12, 2115. https://doi.org/10.3390/ani12162115
Skoupá K, Šťastný K, Sládek Z. Anabolic Steroids in Fattening Food-Producing Animals—A Review. Animals. 2022; 12(16):2115. https://doi.org/10.3390/ani12162115
Chicago/Turabian StyleSkoupá, Kristýna, Kamil Šťastný, and Zbyšek Sládek. 2022. "Anabolic Steroids in Fattening Food-Producing Animals—A Review" Animals 12, no. 16: 2115. https://doi.org/10.3390/ani12162115