1. Introduction
Besides production efficiency, the physiological functions and health of livestock are attracting more and more attention [
1]. Nutritional strategies have emerged and it has been proposed as a key factor to improve the health status and welfare of animals as well as to enhance productivity [
2,
3]. Modulation of fermentation parameters by supplementing additives to ruminant diets was considered an effective strategy for manipulating rumen function [
4,
5].
The shortage of feed ingredients and the high price of imported protein sources are the main factors restricting the further development of animal husbandry in China [
6]. Especially in the last few years, the price of soybean meal, the major source of protein for livestock feed, has increased substantially, as well as the cost of feedstuff [
7]. Cottonseed meal, the main by-product of the cottonseed oil extraction, contains more than 40% of crude protein (CP) and has been considered as a cheap substitute for soybean meal. However, due to the existence of toxic free gossypol, the proportion of cottonseed meal in animal feed is limited [
8]. For adult ruminants, although the mature microbial system in the rumen is able to convert free gossypol into protein-gossypol complexes, it is not absorbed in the digestive tract thereby hindering its entry into the blood [
9]. Excessive free gossypol intake could escape this protective mechanism and increase gossypol content in the plasma and milk of cows [
10], induce a negative nitrogen balance in lactating goats [
11], and disrupt the integrity of sperm cell membrane in male ruminants [
12]. Moreover, the high content of free gossypol had obvious negative effects on both microbial diversity and activity [
13,
14]. In addition to containing toxic free gossypol, the high fiber, low lysine and methionine contents, and lower digestibility of cottonseed meal limit its use in growing livestock [
15,
16]. Based on these, cottonseed meal has not been fully utilized at present.
Several methods have been proven to reduce free gossypol content in cottonseed meal. Solvent extraction could reduce free gossypol content to less than 0.045%, making cottonseed meal an edible high-quality protein [
17]. The free gossypol in cottonseed meal could be degraded to less than 0.005% by microbial (
C. tropicalis or
S. cerevisae) fermentation [
18]. Enzymolysis is a method of applying protease to reduce free gossypol, while hydrolyzing macromolecular proteins to peptides to improve the nutritional quality of cottonseed meal [
19,
20]. Earlier studies revealed that rumen microbes utilize both ammonia nitrogen (NH
3-N) and non-ammonia nitrogen (e.g., free amino acids and peptides) to synthesize microbial protein (MCP), and some of them preferentially use non-ammonia nitrogen over NH
3-N [
21,
22]. Supplementation of peptides enhanced the efficiency of rumen microbe growth, ruminal fermentation, and fiber digestion in vitro [
23,
24], and nitrogen utilization efficiency in vivo [
25]. Except for utilization by rumen microbes, small peptides obtained from proteolysis of plant proteins can be directly absorbed by the rumen epithelium, thereby more conducive to retaining their original biological activities including improving immunological and antioxidant status [
26,
27]. Enzymolysis would greatly improve the availability of cottonseed meal to alleviate the current shortage of protein resources. The addition of 2% enzymatic hydrolysate of cottonseed protein (ECP) in a starter diet enhanced the antioxidant status of newborn calves, whereas 6% ECP supplementation impaired the starter intake and growth of newborn calves and exposed them to a stressful status [
15]. However, the evaluation of the nutritional value of ECP as a feed material for adult ruminants has not yet been reported.
Yeast culture is a unique yeast product due to the composition of yeasts and their metabolites, and it maintains the fermentation activity of yeast [
28]. The composition and characteristics of these metabolites are determined by multiple factors during yeast cultivation, e.g., culture medium, yeast species, and culture conditions [
29]. Functional metabolites in yeast cultures may contribute to rumen function by providing beneficial nutrients otherwise scarce in the ruminal environment [
30]. Previous studies have shown that yeast culture had a positive effect on rumen fermentation and microbial metabolism [
31,
32]. We hypothesized that supplementation with different levels of ECP had no negative effect on rumen fermentation. In this study, yeast culture was used as a positive control and the primary objective was to investigate the in vitro dynamic changes of ruminal fermentation characteristics, kinetic of gas production, and microbial diversity with different levels of ECP supplementation.
4. Discussion
Gas produced by in vitro fermentation is mainly derived from soluble carbohydrates [
51]. The cumulative gas production gradually increased over time and was affected by different supplements at different times. More specifically, compared with the control group, supplementation of ECP with 12 g/kg of substrate increased the cumulative gas production at 2, 4, and 12 h; supplementation of XP yeast culture with 10 g/kg of substrate increased the cumulative gas production at 12, 24, and 36 h; supplementation of ECP with 18 g/kg of substrate increased the cumulative gas production at 48 h (
Table 2). The data of cumulative gas production implied that supplemented ECP with 12 g/kg of substrate increased the gas production rate in the initial period, supplemented XP yeast culture with 10 g/kg of substrate increased the gas production rate in the medium-term, and supplemented ECP with 18 g/kg of substrate increased the late-stage gas production rate at the late stage during in vitro fermentation. Although there were no statistical differences in the kinetic parameters of gas production among the five treatments. The value of RahG (the average gas production rate at the time when half of the asymptotic gas production occurred) in the CON treatment was lower than in the other four treatments (2.9 vs. 3.2, 3.0, 3.2, 3.1). The yeast cultures used in this study contained yeast cell walls that were rich in glucans and mannans. These carbohydrates can be used as prebiotics to increase the activity of rumen microorganisms and to improve rumen fermentation [
52]. Meanwhile, the glucans and mannans can be highly degraded in the rumen, producing more in vitro gas [
53]. As a fermentation substrate, the rumen degradable amino acid content is another key factor affecting the growth rate of rumen microbes [
54]. On the other hand, rumen available nitrogen in the form of peptides and amino acids can contribute to the digestion of structural carbohydrates and non-structural carbohydrates by rumen microbes [
25]. Thus, it seems that ECP supplementation accelerated the fermentation and gas production processes, implicating that ECP or XP yeast culture supplementation enhanced the availability of soluble carbohydrates from substrates and that such beneficial effect was pronounced in the XP and ECP2 treatments.
Nitrogen metabolism in the rumen begins with the degradation of dietary protein or non-protein nitrogen by microorganisms into NH
3-N, which is subsequently converted into MCP [
55]. Microbial protein synthesized in the rumen is an important source of amino acids available for ruminants [
56,
57]. Compared with the control treatment, supplementation of yeast culture increased the NH
3-N concentration at 24 h and the MCP concentration at 24 and 48 h in the incubation fluids; supplementation of ECP with 18 g/kg of substrate increased the NH
3-N concentration at 12 and 36 h, increased the MCP concentration at 12, 36, and 48 h in the incubation fluids. Meanwhile, the NH
3-N and MCP concentrations increased linearly and quadratically with the ECP supplementation at 36 h. The synthesis efficiency of MCP in the rumen is determined by the abundance of microbes that synthesize it and its ability to utilize NH
3-N [
51]. Both XP yeast culture and ECP could be used as a fermentation substrate so as to stimulate the growth of some ruminal microorganisms and improve the utilization of NH
3-N.
Volatile fatty acids, the major end-products of rumen fermentation, are the main energy sources for ruminants. The composition and production of VFAs are important indicators of the rumen fermentation function [
49,
51]. For high lactation cows, acetate and butyrate produced in the rumen are substances for the synthesis of milk lipids, and 66.7% of the glucose used to produce lactose comes from gluconeogenesis from propionate [
58,
59]. The energy efficiency improved with the increased production of propionate and decreased production of acetate [
60]. Because most carbohydrate fermentation by rumen bacteria results in a higher propionate concentration, an increase in propionate concentration often causes a drop in the ratio of acetate to butyrate in the rumen [
61]. In this study, supplementation of XP yeast culture with 10 g/kg of substrate decreased the molar acetate proportion and increased the molar butyrate and valerate proportion at 48 h and decreased the ratio of acetate to proportion, suggesting that supplementation of XP yeast culture changed the fermentation pattern in vitro.
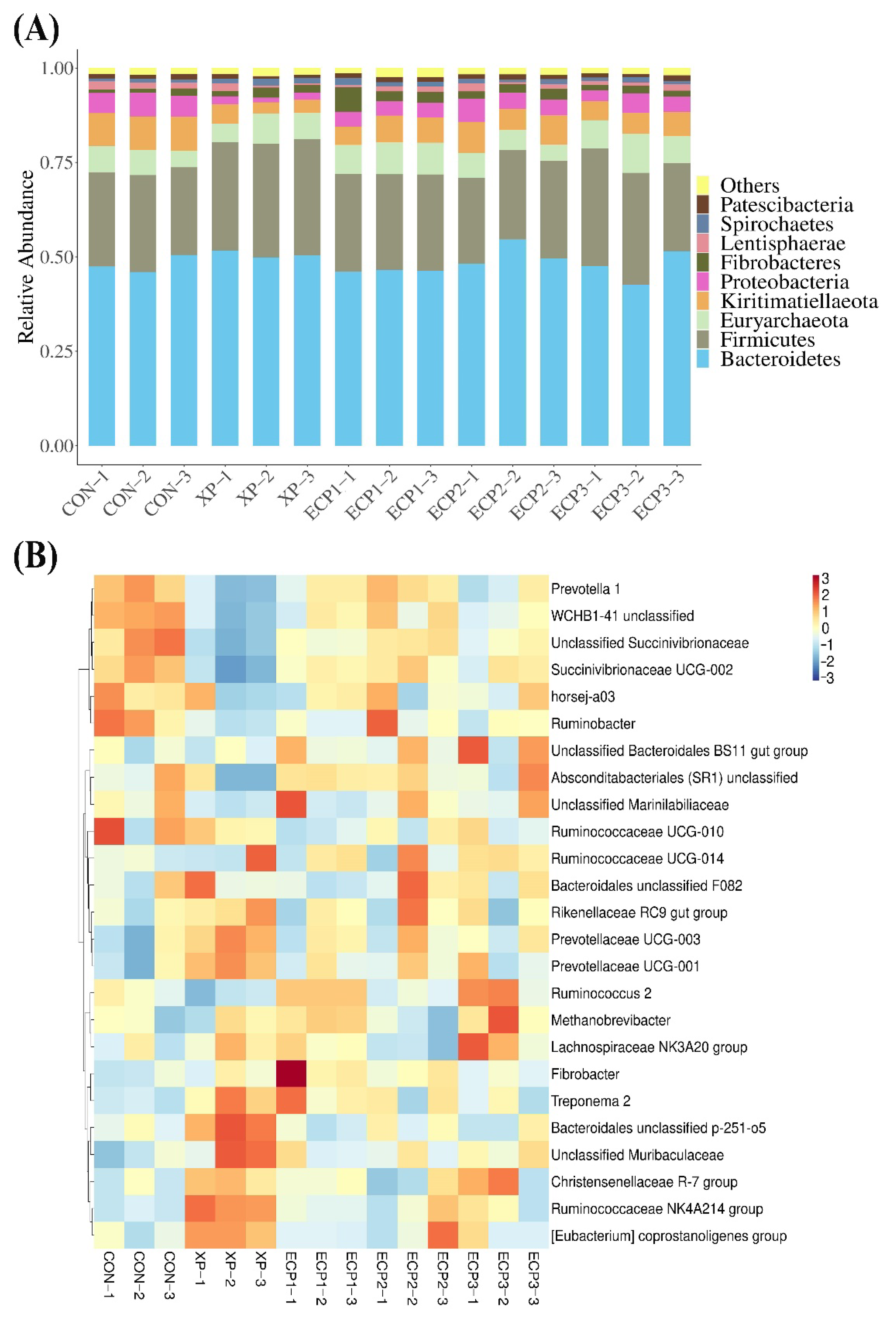
Previous studies have confirmed that the dominant phyla in rumen of ruminants are Firmicutes and Bacteroidetes [
62], which is also supported by data from this study (
Figure 3A and
Table 6). This illustrated that the bacterial composition and proportion of the in vitro fermentation system in this study was the same as that of the cow rumen. The Bacteroidetes in rumen are active in the degradation of non-cellulosic plant constituent and the production of acetate and propionate [
63]. The main members of the phylum Firmicutes are diverse fibrolytic and cellulolytic bacterial genera that are involved in the degradation of cellulose, hemicellulose, oligosaccharides, and starch, improving the digestion and utilization of fibers in ruminants [
64]. The phyla Kiritimatiellaeota and Proteobacteria have been suggested to be involved in the degradation of fiber and organic matter, respectively [
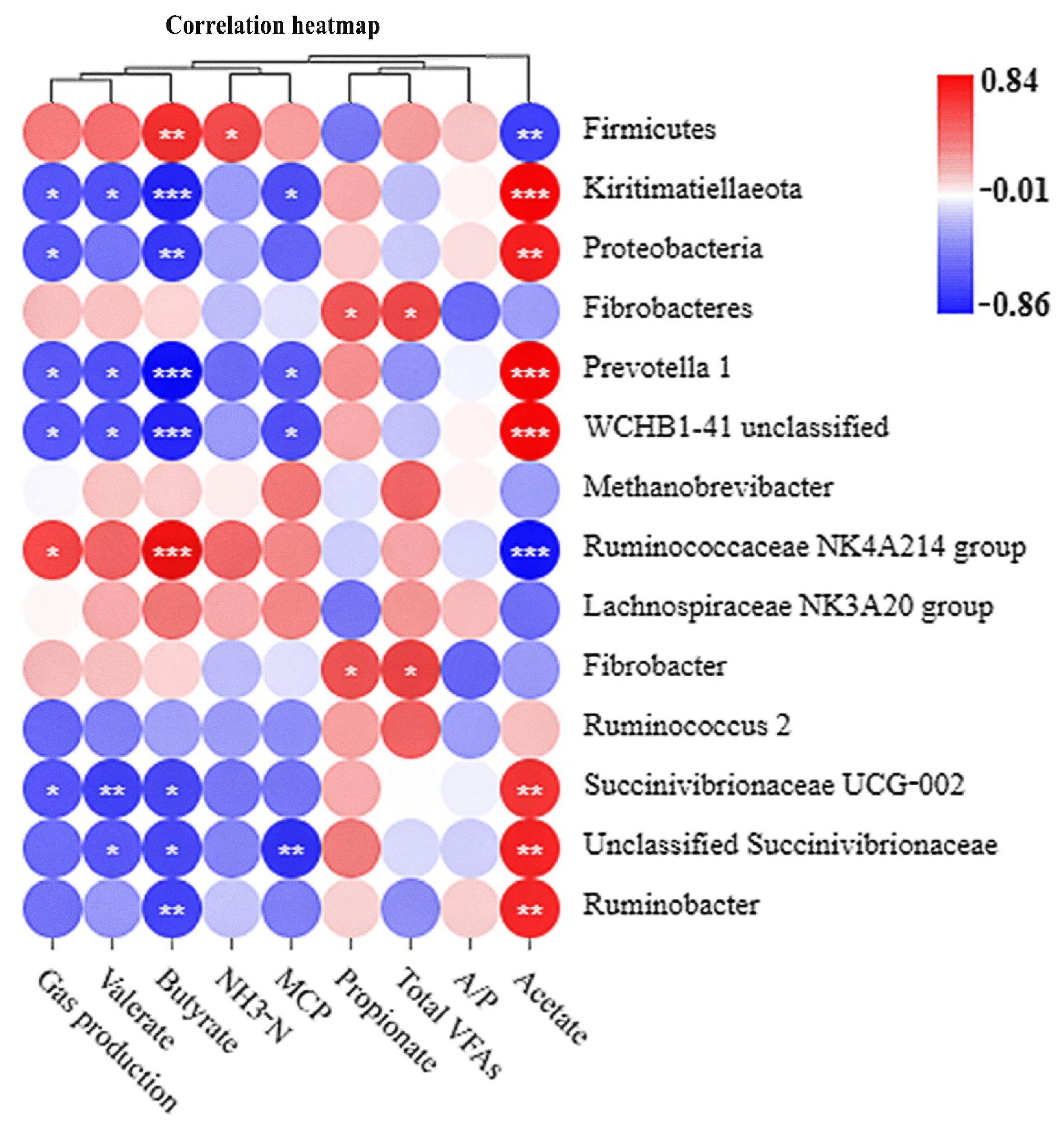
64]. Supplementation of XP yeast culture with 10 g/kg of substrate increased the relative abundance of the phylum Firmicutes while decreased the relative abundance of the phyla Kiritimatiellaeota and Proteobacteria, compared with the control group. Supplementation of ECP with 6 and 18 g/kg of substrate decreased the relative abundance of the phyla Kiritimatiellaeota and Proteobacteria while supplementation of ECP with 6 and 12 g/kg of substrate increased the relative abundance of the phylum Fibrobacteres, compared with the control group. The results of correlation analysis showed that the phylum Firmicutes was positively associated with the molar butyrate proportion while negatively associated with the molar butyrate proportion. In contrast, the phyla Kiritimatiellaeota and Proteobacteria were negatively associated with the molar butyrate proportion while positively associated with the molar butyrate proportion. These results indicated that supplementation with XP yeast culture or ECP had different effects on microbial diversity during in vitro fermentation.
Members of the genus
Prevotella are considered to be the most abundant group of ruminal bacteria [
65,
66], and several enzymes involved in fiber digestion have been identified in members of this genus [
67,
68]. In this study, the genus
Prevotella 1 was positively associated with the molar acetate proportion while negatively associated with gas production, MCP concentration, and the molar proportions of butyrate and valerate. The low abundance of
Prevotella 1 could be the reason for the low molar acetate proportion in the XP treatment. The lowest function percentage of energy metabolism and concentration of total VFAs in the XP treatment suggested that supplementation of XP yeast culture with 10 g/kg of substrate may not be beneficial to energy metabolism activities of rumen bacteria. The members of the family Ruminococcaceae are another major component of Firmicutes in the degradation of fiber [
69]. The genus
Ruminococcaceae NK4A214 group belongs to the family Ruminococcaceae, and although it was first identified and cultured in 2011 [
70], its precise function in the rumen is still not clear [
71]. In this study, the genus
Ruminococcaceae NK4A214 group was positively associated with gas production and the molar butyrate proportion while negatively associated with the molar acetate proportion. Previous study has shown that high dietary soluble carbohydrates increased the content of butyrate in the rumen of dairy cows [
72]. Thus, we speculate that the function of the genus
Ruminococcaceae NK4A214 group may be related to degradation of soluble carbohydrates. Although accounting for a relatively small proportion, the high fibrolytic activity makes genera
Fibrobacter and
Ruminococcus play a nonnegligible role in fiber degradation [
73]. Supplementation of ECP with 6 g/kg of substrate increased the relative abundance of the genera
Fibrobacter and
Ruminococcus, which could result in the increase of the concentration of total VFAs at 24 h (63.8 vs. 71.6 mmol/L). In fact, microbes that account for a larger proportion of abundance contribute significantly to the function of the rumen microbial ecosystem, and the small group of bacteria in the rumen community may possess important but as yet unrecognized ecological functions [
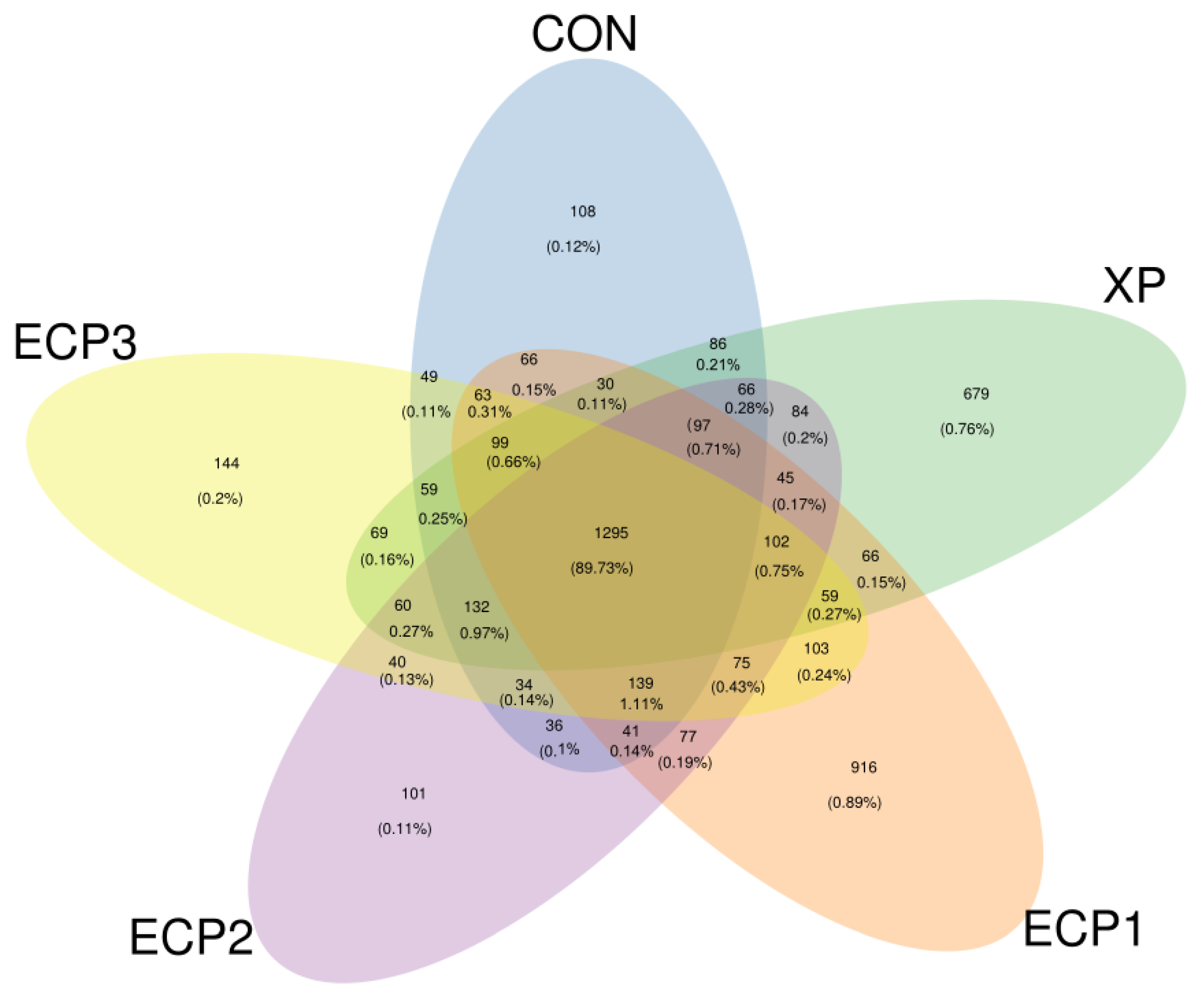
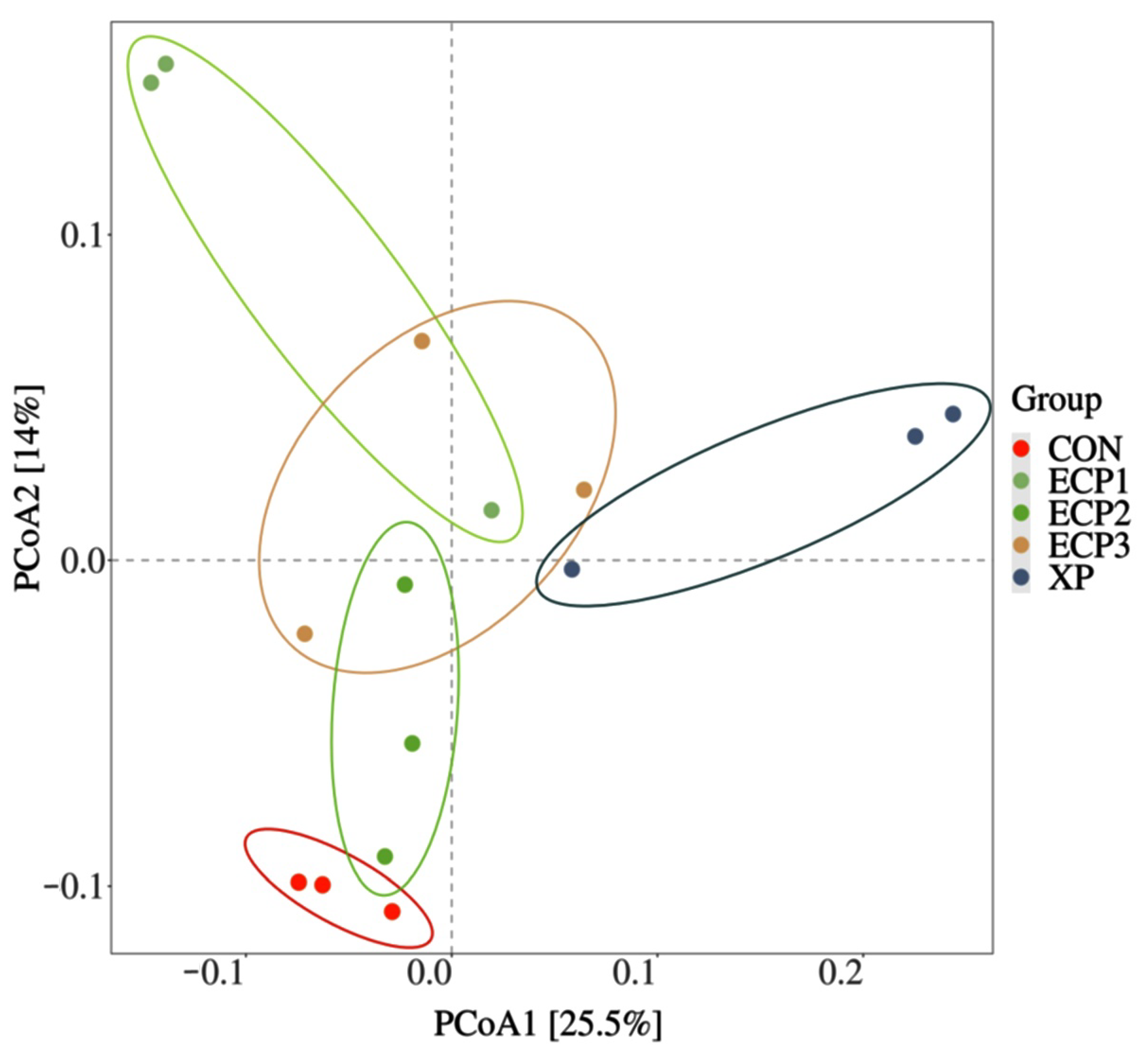
69]. One of the limitations of this study is the low number of repetitions, which should be noted in future experiments. Meanwhile, the bacterial diversity at 24 h may be not enough to reveal the relationship between bacteria and rumen fermentation parameters.


 ,
, 






{kind=link}
{kind=link}
{kind=link}
{kind=link}