
Effects of Dietary Nucleotide Supplementation on Performance, Profitability, and Disease Resistance of Litopenaeus vannamei Cultured in Indonesia under Intensive Outdoor Pond Conditions




Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Diets
2.2. Growth Trial
2.3. Body Composition Analysis
2.4. Water Quality and Growth Sampling
2.5. Total Hemocyte Count
2.6. Lysozyme Activity Analysis
2.7. Histology
2.8. Challenge Test
2.9. Organoleptic Analysis
2.10. Profitability Analysis
2.11. Statistical Analysis
3. Results
3.1. Water Quality
3.2. Growth Performance
3.3. Body Composition Analysis and Organoleptic Evaluation
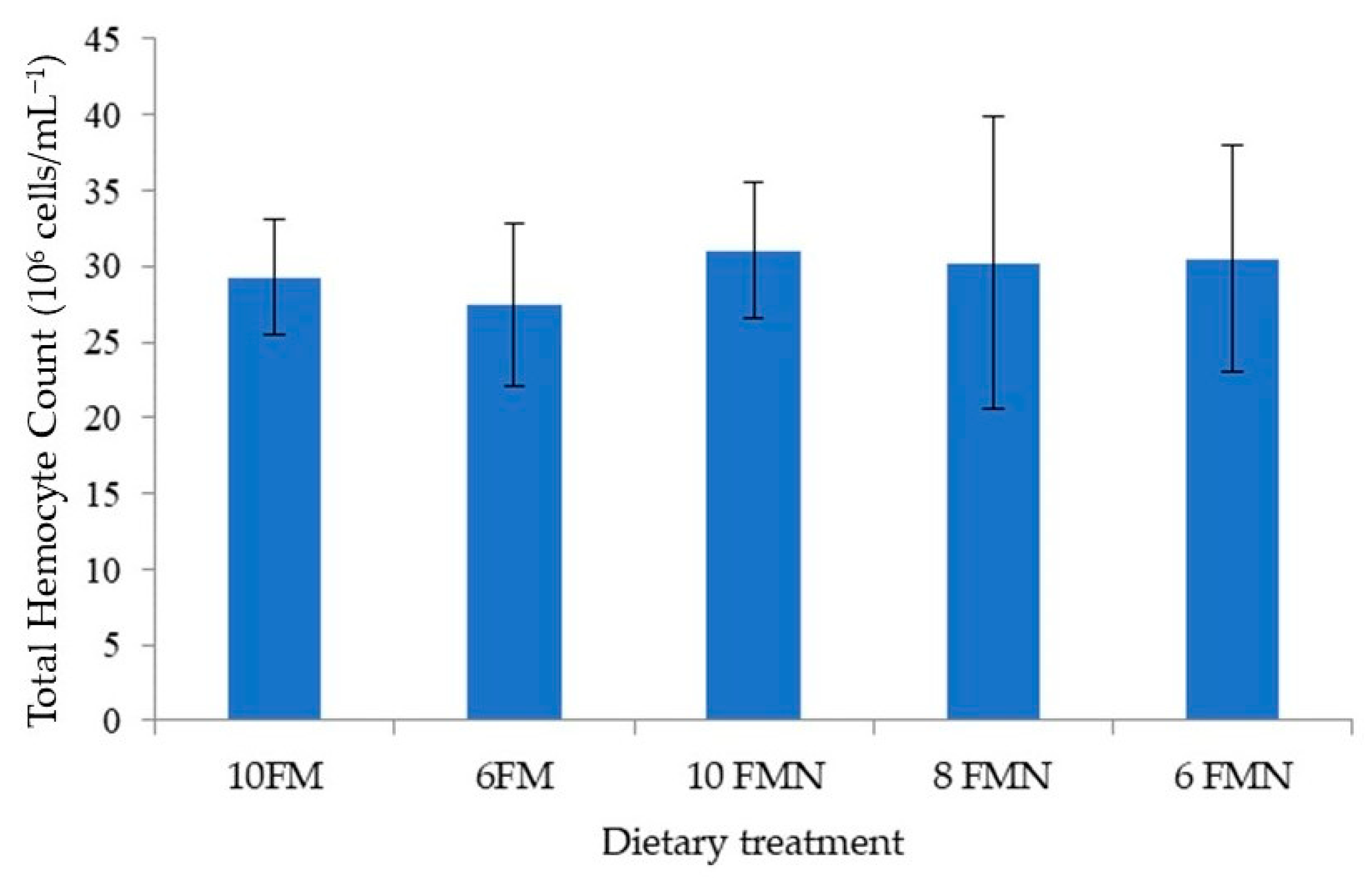
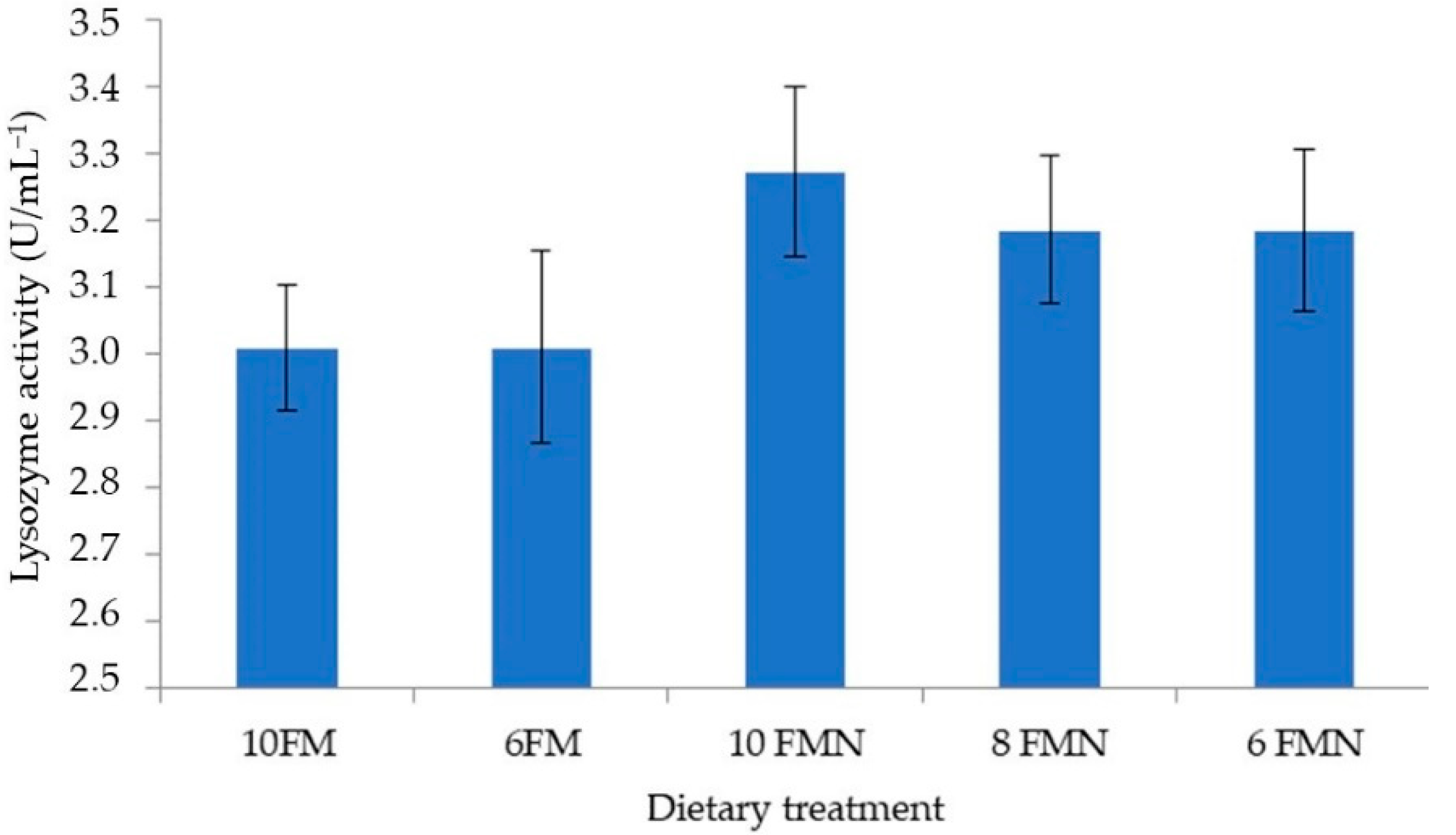
3.4. Total Hemocyte Counts and Lysozyme Activity
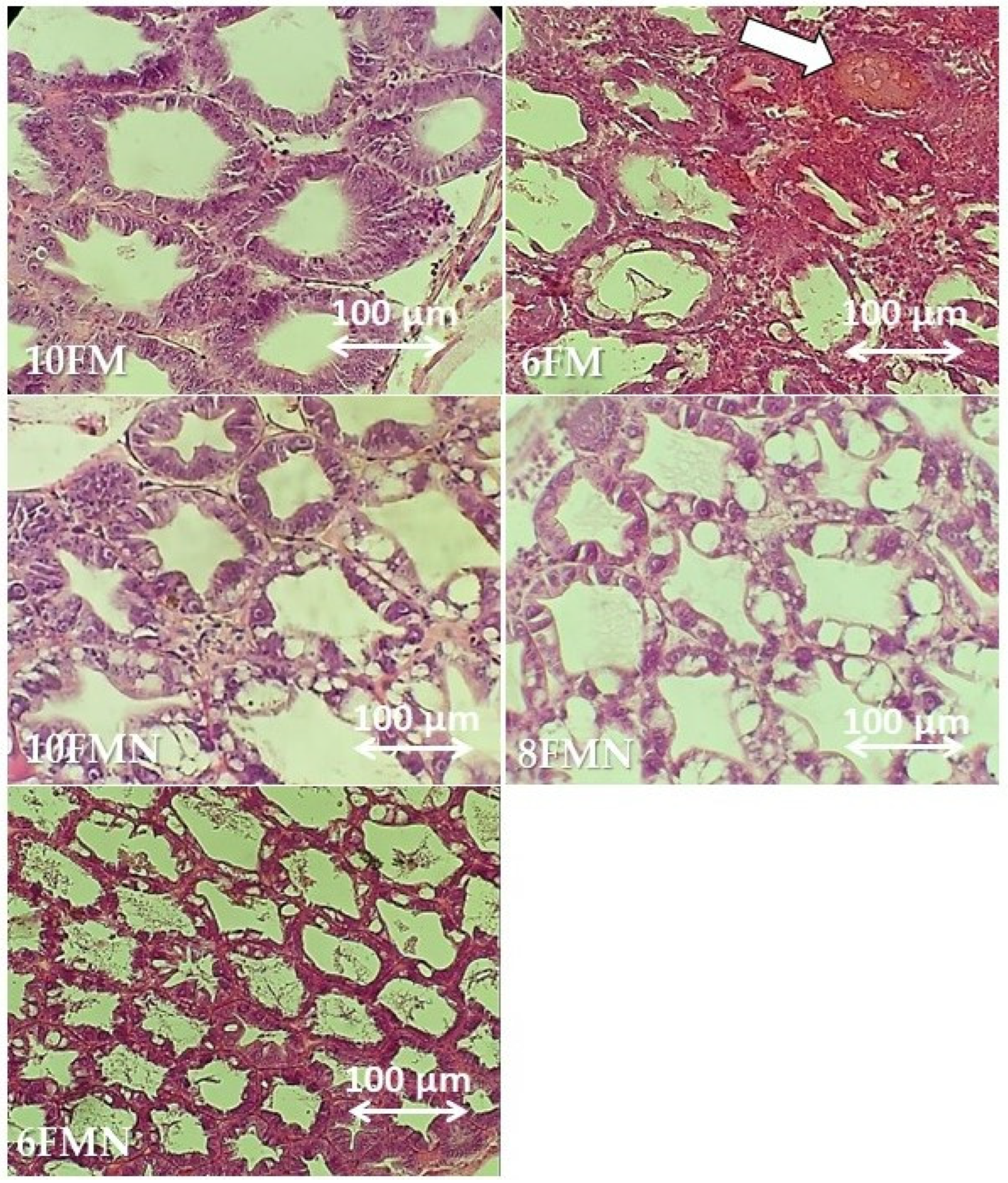
3.5. Histomorphological Condition of Shrimp Hepatopancreas
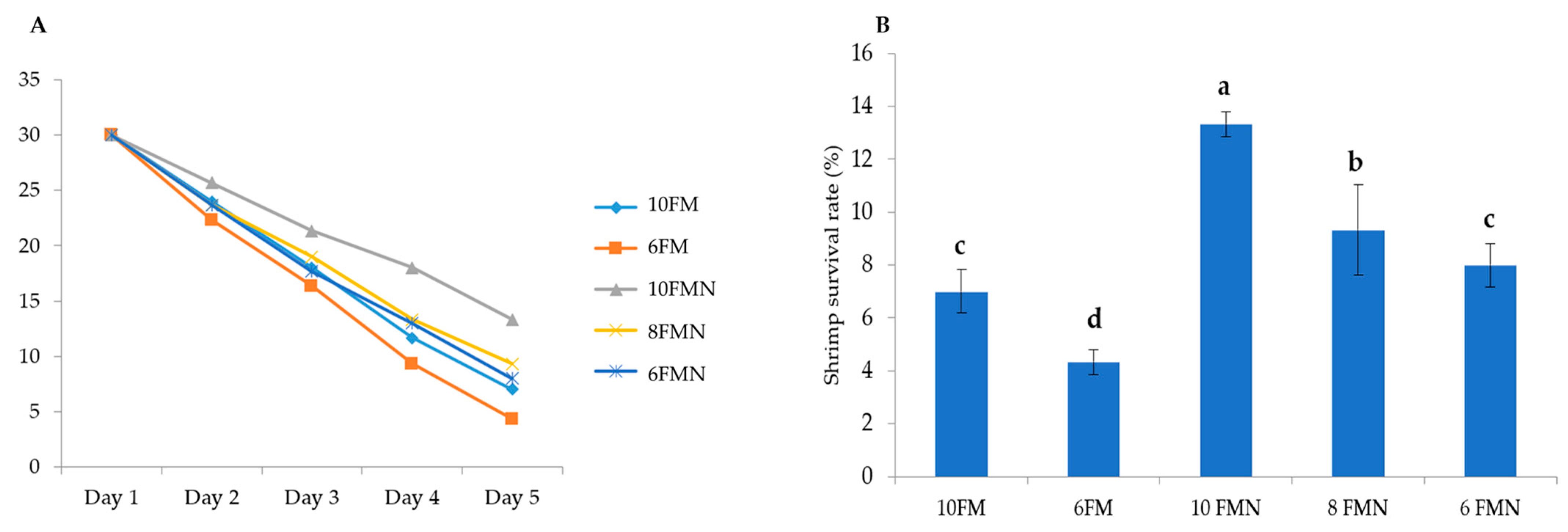
3.6. Challenge Test
3.7. Organoleptic Evaluation
3.8. Profitability
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liao, I.C.; Chien, Y.-H. The pacific white shrimp, Litopenaeus vannamei, in Asia: The world’s most widely cultured alien crustacean. In In the Wrong Place-Alien Marine Crustaceans: Distribution, Biology and Impacts; Springer: Dordrecht, The Netherlands, 2011; pp. 489–519. [Google Scholar]
- Farfante, I.P.; Kensley, B. Penaeoid and Sergestoid Shrimps and Prawns of the World. Keys and Diagnoses for the Families and Genera; Editions du Museum national d’Histoire naturelle: Paris, France, 1997. [Google Scholar]
- Paquotte, P.; Chim, L.; Martin, J.-L.; Lemos, E.; Stern, M.; Tosta, G. Intensive culture of shrimp Penaeus vannamei in floating cages: Zootechnical, economic and environmental aspects. Aquaculture 1998, 164, 151–166. [Google Scholar] [CrossRef]
- Sookying, D.; Davis, D.; da Silva, F.S. A review of the development and application of soybean-based diets for Pacific white shrimp Litopenaeus vannamei. Aquac. Nutr. 2013, 19, 441–448. [Google Scholar] [CrossRef]
- Zulkarnain, R.; Adiyana, K.; Nugroho, H.; Nugraha, B.; Thesiana, L.; Supriyono, E. Selection of intensive shrimp farming technology for small farmers with analytical hierarchy process: A case for whiteleg shrimp (Litopenaeus vannamei). In Proceedings of the IOP Conference Series: Earth and Environmental Science, Changchun, China, 21–23 August 2020; p. 012017. [Google Scholar]
- Nurhafizah, W.W.I.; Lee, K.L.; Nadirah, M.; Danish-Daniel, M.; Zainathan, S.C.; Najiah, M. Virulence properties and pathogenicity of multidrug-resistant Vibrio harveyi associated with luminescent vibriosis in Pacific white shrimp, Penaeus vannamei. J. Invertebr. Pathol. 2021, 186, 107594. [Google Scholar] [CrossRef]
- Novriadi, R.; Albasri, H.; Eman, C.M. Tinjauan indikator kesiapan produksi udang putih Litopenaeus vannamei (Boone, 1931) di sistem intensif. Sains Akuakultur Trop. Indones. J. Trop. Aquac. 2021, 5, 252–271. [Google Scholar]
- Sookying, D.; Davis, D.A. Pond production of Pacific white shrimp (Litopenaeus vannamei) fed high levels of soybean meal in various combinations. Aquaculture 2011, 319, 141–149. [Google Scholar] [CrossRef]
- Fox, J.M.; Lawrence, A.L.; Smith, F. Development of a low-fish meal feed formulation for commercial production of Litopenaeus vannamei. Avances En Nutrición Acuicola 2004. Available online: https://nutricionacuicola.uanl.mx/index.php/acu/article/view/199 (accessed on 1 August 2022).
- Braga, A.; Magalhães, V.; Hanson, T.; Morris, T.C.; Samocha, T.M. The effects of feeding commercial feed formulated for semi-intensive systems on Litopenaeus vannamei production and its profitability in a hyper-intensive biofloc-dominated system. Aquac. Rep. 2016, 3, 172–177. [Google Scholar] [CrossRef] [Green Version]
- Quyen, N.T.K.; Hien, H.V.; Khoi, L.N.D.; Yagi, N.; Karia Lerøy Riple, A. Quality management practices of intensive whiteleg shrimp (Litopenaeus vannamei) farming: A study of the Mekong Delta, Vietnam. Sustainability 2020, 12, 4520. [Google Scholar] [CrossRef]
- Flegel, T.W. Current status of viral diseases in Asian shrimp aquaculture. Isr. J. Aquac. Bamidgeh 2009, 61, 229–239. [Google Scholar] [CrossRef]
- Thitamadee, S.; Prachumwat, A.; Srisala, J.; Jaroenlak, P.; Salachan, P.V.; Sritunyalucksana, K.; Flegel, T.W.; Itsathitphaisarn, O. Review of current disease threats for cultivated penaeid shrimp in Asia. Aquaculture 2016, 452, 69–87. [Google Scholar] [CrossRef]
- Olmos, J.; Ochoa, L.; Paniagua-Michel, J.; Contreras, R. Functional feed assessment on Litopenaeus vannamei using 100% fish meal replacement by soybean meal, high levels of complex carbohydrates and Bacillus probiotic strains. Mar. Drugs 2011, 9, 1119–1132. [Google Scholar] [CrossRef] [Green Version]
- Encarnação, P. Functional feed additives in aquaculture feeds. In Aquafeed Formulation; Elsevier: Amsterdam, The Netherlands, 2016; pp. 217–237. [Google Scholar]
- Novriadi, R.; Fadhilah, R.; Wahyudi, A.E.; Trullas, C. Effects of hydrolysable tannins on the growth performance, total haemocyte counts and lysozyme activity of Pacific white leg shrimp Litopenaeus vannamei. Aquac. Rep. 2021, 21, 100796. [Google Scholar] [CrossRef]
- Novriadi, R.; Albasri, H.; Wahyudi, A.E.; Fadhilah, R.; Ali, A.; Trullàs, C. Effects of the addition of oak (Quercus robur L.) and yucca (Yucca schidigera) on the water quality and growth performance of pacific white shrimp (Litopenaeus vannamei) cultured intensively in concrete tanks. J. World Aquac. Soc. 2022. [Google Scholar] [CrossRef]
- Andrino, K.G.S.; Augusto, E.; Serrano, J.; Valeriano, L.; Corre, J. Effects of dietary nucleotides on the immune response and growth of juvenile Pacific white shrimp Litopenaeus vannamei (Boone, 1931). Asian Fish. Sci. 2012, 25, 180–192. [Google Scholar] [CrossRef]
- Li, P.; Lawrence, A.L.; Castille, F.L.; Gatlin, D.M., III. Preliminary evaluation of a purified nucleotide mixture as a dietary supplement for Pacific white shrimp Litopenaeus vannamei (Boone). Aquac. Res. 2007, 38, 887–890. [Google Scholar] [CrossRef]
- Segarra, S. Nutritional Modulation of the Immune Response Mediated by Nucleotides in Canine Leishmaniosis. Microorganisms 2021, 9, 2601. [Google Scholar] [CrossRef]
- Novriadi, R.; Ilham, I.; Roigé, O.; Segarra, S. Effects of dietary nucleotides supplementation on growth, total haemocyte count, lysozyme activity and survival upon challenge with Vibrio harveyi in pacific white shrimp, Litopenaeus vannamei. Aquac. Rep. 2021, 21, 100840. [Google Scholar] [CrossRef]
- Soto-Rodriguez, S.A.; Gomez-Gil, B.; Lozano, R. ‘Bright-red’syndrome in Pacific white shrimp Litopenaeus vannamei is caused by Vibrio harveyi. Dis. Aquat. Org. 2010, 92, 11–19. [Google Scholar] [CrossRef] [Green Version]
- Austin, B.; Zhang, X.H. Vibrio harveyi: A significant pathogen of marine vertebrates and invertebrates. Lett. Appl. Microbiol. 2006, 43, 119–124. [Google Scholar] [CrossRef]
- Novriadi, R. Vibriosis in aquaculture. Omni-Akuatika 2016, 12, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Fang, W.; Yang, X.; Zhou, S.; Hu, L.; Li, X.; Qi, X.; Su, H.; Xie, L. A nonluminescent and highly virulent Vibrio harveyi strain is associated with “bacterial white tail disease” of Litopenaeus vannamei shrimp. PLoS ONE 2012, 7, e29961. [Google Scholar] [CrossRef]
- Montero, A.; Austin, B. Characterization of extracellular products from an isolate of Vibrio harveyi recovered from diseased post-larval Penaeus vannamei (Bonne). J. Fish Dis. 1999, 22, 377–386. [Google Scholar] [CrossRef]
- Franco, A.; Ferreira, J.; Nobre, A. Development of a growth model for penaeid shrimp. Aquaculture 2006, 259, 268–277. [Google Scholar] [CrossRef]
- Helrich, K. Association of Official Analytical Chemists. In Official Methods of Analysis of the Association of Official Analytical Chemists, 15th ed.; Association of Official Analytical Chemists: Arlington, VA, USA, 1990. [Google Scholar]
- Boyd, C.E.; Tucker, C.S. Water quality and pond soil analyses for aquaculture. In Water Quality and Pond Soil Analyses for Aquaculture; The University of Alabama Press: Tuscaloosa, AL, USA, 1992. [Google Scholar]
- Del Carmen Flores-Miranda, M.; Luna-González, A.; Campa-Córdova, Á.I.; González-Ocampo, H.A.; Fierro-Coronado, J.A.; Partida-Arangure, B.O. Microbial immunostimulants reduce mortality in whiteleg shrimp (Litopenaeus vannamei) challenged with Vibrio sinaloensis strains. Aquaculture 2011, 320, 51–55. [Google Scholar] [CrossRef]
- Seong, M.; Lee, S.; Lee, S.; Song, Y.; Bae, J.; Chang, K.; Bai, S.C. The effects of different levels of dietary fermented plant-based protein concentrate on growth, hematology and non-specific immune responses in juvenile olive flounder, Paralichthys olivaceus. Aquaculture 2018, 483, 196–202. [Google Scholar] [CrossRef]
- Hasson, K.; Lightner, D.; Poulos, B.; Redman, R.; White, B.; Brock, J.; Bonami, J. Taura syndrome in Penaeus vannamei: Demonstration of a viral etiology. Dis. Aquat. Org. 1995, 23, 115–126. [Google Scholar] [CrossRef] [Green Version]
- Le, N.T.; Doan, N.K.; Ba, T.N.; Tran, T.V.T. Towards improved quality benchmarking and shelf life evaluation of black tiger shrimp (Penaeus monodon). Food Chem. 2017, 235, 220–226. [Google Scholar] [CrossRef]
- Hossain, M.S.; Koshio, S.; Kestemont, P. Recent advances of nucleotide nutrition research in aquaculture: A review. Rev. Aquac. 2020, 12, 1028–1053. [Google Scholar] [CrossRef]
- Ringø, E.; Olsen, R.E.; Vecino, J.G.; Wadsworth, S.; Song, S. Use of immunostimulants and nucleotides in aquaculture: A review. J. Mar. Sci. Res. Dev. 2012, 2, 104. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.; Guo, B.; Zhang, H.; Xu, W.; Zhang, W.; Mai, K. Effects of nucleotides on growth performance, immune response, disease resistance and intestinal morphology in shrimp Litopenaeus vannamei fed with a low fish meal diet. Aquac. Int. 2016, 24, 1007–1023. [Google Scholar] [CrossRef]
- Xiong, J.; Jin, M.; Yuan, Y.; Luo, J.X.; Lu, Y.; Zhou, Q.C.; Liang, C.; Tan, Z.L. Dietary nucleotide-rich yeast supplementation improves growth, innate immunity and intestinal morphology of Pacific white shrimp (Litopenaeus vannamei). Aquac. Nutr. 2018, 24, 1425–1435. [Google Scholar] [CrossRef]
- Burrells, C.; Williams, P.; Forno, P. Dietary nucleotides: A novel supplement in fish feeds: 1. Effects on resistance to disease in salmonids. Aquaculture 2001, 199, 159–169. [Google Scholar] [CrossRef]
- Hossain, M.S.; Koshio, S.; Ishikawa, M.; Yokoyama, S.; Sony, N.M.; Dawood, M.A.; Kader, M.A.; Bulbul, M.; Fujieda, T. Efficacy of nucleotide related products on growth, blood chemistry, oxidative stress and growth factor gene expression of juvenile red sea bream, Pagrus major. Aquaculture 2016, 464, 8–16. [Google Scholar] [CrossRef]
- Lin, Y.H.; Wang, H.; Shiau, S.Y. Dietary nucleotide supplementation enhances growth and immune responses of grouper, Epinephelus malabaricus. Aquac. Nutr. 2009, 15, 117–122. [Google Scholar] [CrossRef]
- Metailler, R.; Cadena-Roa, M.; Ruyet, J.P.L. Attractive chemical substances for the weaning of Dover sole (Solea vulgaris): Qualitative and quantitative approach. J. World Maric. Soc. 1983, 14, 679–684. [Google Scholar] [CrossRef]
- Johansson, M.; Soderhall, K. Cellular immunity in crustaceans and the proPO system. Parasitol. Today 1989, 5, 171–176. [Google Scholar] [CrossRef]
- Cerenius, L.; Söderhäll, K. The prophenoloxidase-activating system in invertebrates. Immunol. Rev. 2004, 198, 116–126. [Google Scholar] [CrossRef]
- Xu, Z.; Guan, W.; Xie, D.; Lu, W.; Ren, X.; Yuan, J.; Mao, L. Evaluation of immunological response in shrimp Penaeus vannamei submitted to low temperature and air exposure. Dev. Comp. Immunol. 2019, 100, 103413. [Google Scholar] [CrossRef]
- Pan, L.-Q.; Hu, F.-W.; Jing, F.-T.; Liu, H.-J. The effect of different acclimation temperatures on the prophenoloxidase system and other defence parameters in Litopenaeus vannamei. Fish Shellfish Immunol. 2008, 25, 137–142. [Google Scholar] [CrossRef]
- Murthy, H.S.; Li, P.; Lawrence, A.; Gatlin, D., III. Dietary β-glucan and nucleotide effects on growth, survival and immune responses of pacific white shrimp, Litopenaeus vannamei. J. Appl. Aquac. 2009, 21, 160–168. [Google Scholar] [CrossRef]
- Shankar, R.; Murthy, H.S.; Sujatha, H.; Jayaraj, E.; Tejpal, C.; Chinthamani, V. Effect of nucleotide on growth, immune responses and resistance of Macrobrachium rosenbergii (De Man) to Macrobrachium rosenbergii nodavirus (MrNV) and extra small virus (XSV) and Aeromonas hydrophila infection. Aquac. Int. 2012, 20, 1–12. [Google Scholar] [CrossRef]
- De-La-Re-Vega, E.; García-Orozco, K.D.; Calderón-Arredondo, S.A.; Romo-Figueroa, M.G.; Islas-Osuna, M.A.; Yepiz-Plascencia, G.M.; Sotelo-Mundo, R.R. Recombinant expression of marine shrimp lysozyme in Escherichia coli. Electron. J. Biotechnol. 2004, 7, 12–13. [Google Scholar]
- Biswas, G.; Korenaga, H.; Nagamine, R.; Kono, T.; Shimokawa, H.; Itami, T.; Sakai, M. Immune stimulant effects of a nucleotide-rich baker’s yeast extract in the kuruma shrimp, Marsupenaeus japonicus. Aquaculture 2012, 366, 40–45. [Google Scholar] [CrossRef]
- Kurland, C. The role of guanine nucleotides in protein biosynthesis. Biophys. J. 1978, 22, 373–392. [Google Scholar] [CrossRef] [Green Version]
- Tacon, A.; Cooke, D. Nutritional value of dietary nucleic acids to trout. Nutr. Rep. Int. 1980, 22, 631–640. [Google Scholar]
- Council, N.R. Nutrient Requirements of Fish and Shrimp; National Academies Press: Washington, DC, USA, 2011. [Google Scholar]
- Feng, J.; Liu, X.; Xu, Z.; Liu, Y.; Lu, Y. Effects of Aspergillus oryzae 3.042 fermented soybean meal on growth performance and plasma biochemical parameters in broilers. Anim. Feed Sci. Technol. 2007, 134, 235–242. [Google Scholar] [CrossRef]
- Yao, W.; Zhang, C.; Li, X.; He, M.; Wang, J.; Leng, X. The replacement of fish meal with fermented soya bean meal or soya bean meal in the diet of Pacific white shrimp (Litopenaeus vannamei). Aquac. Res. 2020, 51, 2400–2409. [Google Scholar] [CrossRef]
- Soller, F.; Rhodes, M.; Davis, D. Replacement of fish oil with alternative lipid sources in plant-based practical feed formulations for marine shrimp (Litopenaeus vannamei) reared in outdoor ponds and tanks. Aquac. Nutr. 2017, 23, 63–75. [Google Scholar] [CrossRef]
- Amaya, E.A.; Davis, D.A.; Rouse, D.B. Replacement of fish meal in practical diets for the Pacific white shrimp (Litopenaeus vannamei) reared under pond conditions. Aquaculture 2007, 262, 393–401. [Google Scholar] [CrossRef]
- Dios, D. Fishmeal replacement with feather-enzymatic hydrolyzates co-extruded with soya-bean meal in practical diets for the Pacific white shrimp (Litopenaeus vannamei). Aquac. Nutr. 2001, 7, 143–151. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient (%, as Is) | Diet | ||||
|---|---|---|---|---|---|
| 10FM | 6FM | 10FMN | 8FMN | 6FMN | |
| Menhaden fish meal 1 | 10.00 | 6.00 | 10.00 | 8.00 | 6.00 |
| Soybean meal 1 | 43.00 | 49.50 | 43.00 | 44.80 | 49.50 |
| Corn gluten meal 1 | 10.00 | 10.00 | 10.00 | 10.00 | 10.00 |
| Menhaden fish oil 1 | 5.64 | 5.64 | 5.64 | 5.64 | 5.64 |
| Soy-Lecithin 2 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 |
| Nucleotides 3 | 0.00 | 0.00 | 0.10 | 0.10 | 0.10 |
| Corn starch 1 | 8.06 | 5.56 | 7.96 | 8.16 | 5.46 |
| Wheat products 4 | 17.00 | 17.00 | 17.00 | 17.00 | 17.00 |
| Mineral premix 2* | 0.70 | 0.70 | 0.70 | 0.70 | 0.70 |
| Vitamin premix 2# | 1.90 | 1.90 | 1.90 | 1.90 | 1.90 |
| KP-dibasic 2 | 2.50 | 2.50 | 2.50 | 2.50 | 2.50 |
| Choline chloride 2 | 0.20 | 0.20 | 0.20 | 0.20 | 0.20 |
| Formulation cost (IDR/Kg) | 13.137 | 12.727 | 13.688 | 13.489 | 13.278 |
| Formulation cost (USD/Kg) 5 | 0.9178 | 0.8892 | 0.9553 | 0.9414 | 0.9267 |
| Diet Code | |||||
|---|---|---|---|---|---|
| 10FM | 6FM | 10FMN | 8FMN | 6FMN | |
| Proximate analysis (%, as is) * | |||||
| Protein content | 36.55 | 36.07 | 36.95 | 36.57 | 35.99 |
| Fat content | 6.75 | 6.00 | 6.07 | 6.28 | 6.22 |
| Moisture content | 7.64 | 7.58 | 8.70 | 9.08 | 9.23 |
| Crude fiber | 2.69 | 3.40 | 1.75 | 1.91 | 1.63 |
| Calories (kcal/100 g) * | |||||
| Calories from fat | 60.75 | 54.00 | 54.63 | 56.52 | 64.98 |
| Total calories | 362.47 | 350.32 | 343.67 | 345.32 | 350.82 |
| Amino acid profile (%, as is) * | |||||
| L-Serine | 1.92 | 2.09 | 2.28 | 2.18 | 1.97 |
| L-Glutamic acid | 6.87 | 6.16 | 4.96 | 5.49 | 5.89 |
| L-Phenylalanine | 3.32 | 2.75 | 3.78 | 3.27 | 2.43 |
| L-Isoleusine | 1.26 | 1.41 | 1.41 | 1.39 | 1.39 |
| L-Valine | 1.37 | 1.53 | 1.56 | 1.51 | 1.50 |
| L-Alanine | 1.63 | 1.68 | 1.61 | 1.61 | 1.69 |
| L-Arginine | 2.48 | 2.88 | 3.49 | 3.21 | 2.63 |
| Glycine | 1.73 | 1.81 | 2.13 | 1.93 | 1.74 |
| L-Lysine | 2.86 | 1.71 | 1.60 | 1.80 | 2.23 |
| L-Aspartic acid | 3.00 | 3.25 | 2.52 | 2.79 | 3.27 |
| L-Leusine | 2.65 | 2.88 | 2.94 | 2.90 | 2.81 |
| L-Tyrosine | 1.36 | 1.63 | 2.30 | 2.05 | 1.45 |
| L-Proline | 2.02 | 2.05 | 2.00 | 2.01 | 2.01 |
| L-Threonine | 1.65 | 1.81 | 2.09 | 1.93 | 1.71 |
| L-Histidine | 1.05 | 1.22 | 1.62 | 1.43 | 1.12 |
| L-Cystine | 0.35 | 0.52 | 0.48 | 0.44 | 0.58 |
| L-Methionine | 0.65 | 0.67 | 0.68 | 0.68 | 0.68 |
| L-Tryptophan | 0.32 | 0.36 | 0.33 | 0.34 | 0.33 |
| Diet | Final Biomass (g) | FBW (g) | Survival Pre-Challenge (%) | PWG (%) | FCR | TGC | ADG |
|---|---|---|---|---|---|---|---|
| 10FM | 7604.0 | 20.05 a | 83.31 | 1904.66 a | 1.36 a | 0.05429 a | 0.173 a |
| 6FM | 7405.5 | 19.44 b | 83.91 | 1844.15 b | 1.40 b | 0.05341 b | 0.167 b |
| 10FMN | 7652.5 | 20.28 a | 83.84 | 1928.38 a | 1.34 a | 0.05462 a | 0.175 a |
| 8FMN | 7521.0 | 19.98 a | 84.67 | 1897.90 a | 1.37 a | 0.05419 a | 0.173 a |
| 6FMN | 7534.0 | 19.95 a | 83.93 | 1895.46 a | 1.37 a | 0.05415 a | 0.172 a |
| p-value | 0.1827 | <0.0001 | 0.9336 | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| PSE | 116.0961 | 0.1449 | 1.3703 | 14.4975 | 0.0106 | 0.0002 | 0.0013 |
| Parameter | Unit | Nutritional Composition | |||||
|---|---|---|---|---|---|---|---|
| Control | 10FM | 6FM | 10FMN | 8FMN | 6FMN | ||
| Protein content | % | 24.89 | 22.42 | 22.29 | 24.72 | 24.88 | 23.65 |
| Total fat | % | 1.01 | 0.28 | 0.35 | 0.31 | 0.33 | 0.29 |
| Moisture content | % | 79.69 | 68.72 | 72.77 | 69.43 | 69.96 | 69.72 |
| Carbohydrate | % | 0.65 | 1.56 | 2.45 | 1.88 | 1.70 | 2.14 |
| Ash content | % | 3.76 | 1.59 | 1.78 | 1.71 | 2.27 | 1.82 |
| Calories from fat | KCal 100 g−1 | 4.09 | 4.41 | 4.38 | 4.20 | 4.41 | 4.58 |
| Total calories | KCal 100 g−1 | 71.25 | 113.53 | 109.34 | 111.45 | 115.77 | 110.66 |
| Crude fiber | % | 0.44 | <0.02 | <0.02 | <0.02 | <0.02 | <0.02 |
| L-Cystine | % | 0.23 | 6.44 | 5.31 | 6.02 | 6.14 | 5.89 |
| L-Methionine | % | 0.09 | 4.55 | 4.38 | 4.47 | 4.78 | 4.66 |
| L-Serine | % | 0.37 | 11.53 | 11.02 | 11.15 | 11.53 | 11.44 |
| L-Glutamic acid | % | 1.39 | 31.36 | 29.35 | 32.52 | 32.25 | 31.89 |
| L-Phenylalanine | % | 0.71 | 15.25 | 13.74 | 15.29 | 15.44 | 15.12 |
| L-Isoleucine | % | 0.39 | 11.15 | 10.95 | 11.44 | 10.87 | 10.99 |
| L-Valine | % | 0.45 | 10.77 | 10.82 | 11.34 | 11.33 | 10.39 |
| L-Alanine | % | 0.74 | 12.63 | 12.68 | 12.65 | 12.72 | 12.77 |
| L-Arginine | % | 0.40 | 28.44 | 23.11 | 28.65 | 28.23 | 26.44 |
| Glycine | % | 1.10 | 17.13 | 17.74 | 17.63 | 19.13 | 18.66 |
| L-Lysine | % | 0.53 | 14.56 | 14.22 | 15.07 | 14.37 | 14.44 |
| L-Aspartic acid | % | 0.75 | 16.89 | 17.02 | 18.45 | 17.23 | 16.89 |
| L-Leucine | % | 0.71 | 18.66 | 17.74 | 16.55 | 17.75 | 17.66 |
| L-Tyrosine | % | 0.52 | 11.41 | 10.39 | 10.45 | 10.48 | 10.59 |
| L-Proline | % | 0.41 | 18.78 | 18.85 | 19.02 | 19.11 | 18.59 |
| L-Threonine | % | 0.58 | 11.30 | 10.54 | 11.33 | 11.40 | 10.77 |
| L-Histidine | % | 0.40 | 7.77 | 7.42 | 7.51 | 7.96 | 7.88 |
| L-Tryptophan | % | 0.11 | 1.65 | 1.86 | 1.91 | 1.79 | 1.82 |
| Treatment | Observed Parameter | |||
|---|---|---|---|---|
| Color | Aroma | Flavor | Texture | |
| 10FM | 3.45 | 3.45 | 3.60 | 3.66 |
| 6FM | 3.35 | 3.35 | 3.46 | 3.35 |
| 10FMN | 3.65 | 3.55 | 3.66 | 3.65 |
| 8FMN | 3.40 | 3.46 | 3.55 | 3.55 |
| 6FMN | 3.45 | 3.66 | 3.55 | 3.35 |
| p-value | >0.05 | >0.05 | >0.05 | >0.05 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Novriadi, R.; Roigé, O.; Segarra, S. Effects of Dietary Nucleotide Supplementation on Performance, Profitability, and Disease Resistance of Litopenaeus vannamei Cultured in Indonesia under Intensive Outdoor Pond Conditions. Animals 2022, 12, 2036. https://doi.org/10.3390/ani12162036
Novriadi R, Roigé O, Segarra S. Effects of Dietary Nucleotide Supplementation on Performance, Profitability, and Disease Resistance of Litopenaeus vannamei Cultured in Indonesia under Intensive Outdoor Pond Conditions. Animals. 2022; 12(16):2036. https://doi.org/10.3390/ani12162036
Chicago/Turabian StyleNovriadi, Romi, Oriol Roigé, and Sergi Segarra. 2022. "Effects of Dietary Nucleotide Supplementation on Performance, Profitability, and Disease Resistance of Litopenaeus vannamei Cultured in Indonesia under Intensive Outdoor Pond Conditions" Animals 12, no. 16: 2036. https://doi.org/10.3390/ani12162036