
Shipping Temperature, Time and Media Effects on Equine Wharton’s Jelly and Adipose Tissue Derived Mesenchymal Stromal Cells Characteristics


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Samples
2.2. Cell Isolation and Culture
2.3. Study Design
2.4. CFU (Colony Forming Unit) Assay
2.5. Spheroid Formation Assays
2.6. Multi Lineage In Vitro Differentiation
2.7. RT-PCR
2.8. Statistical Analysis
3. Results
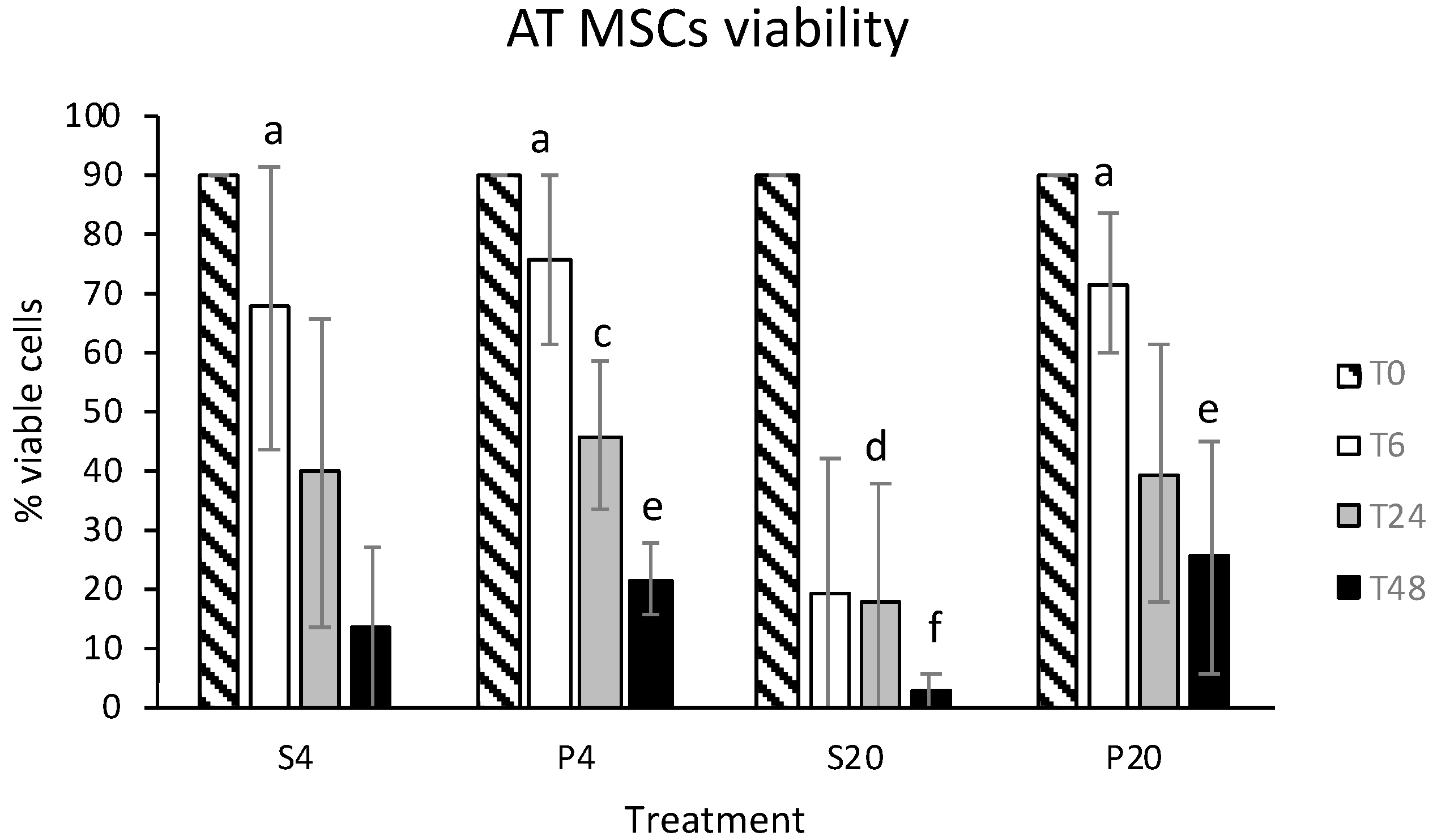
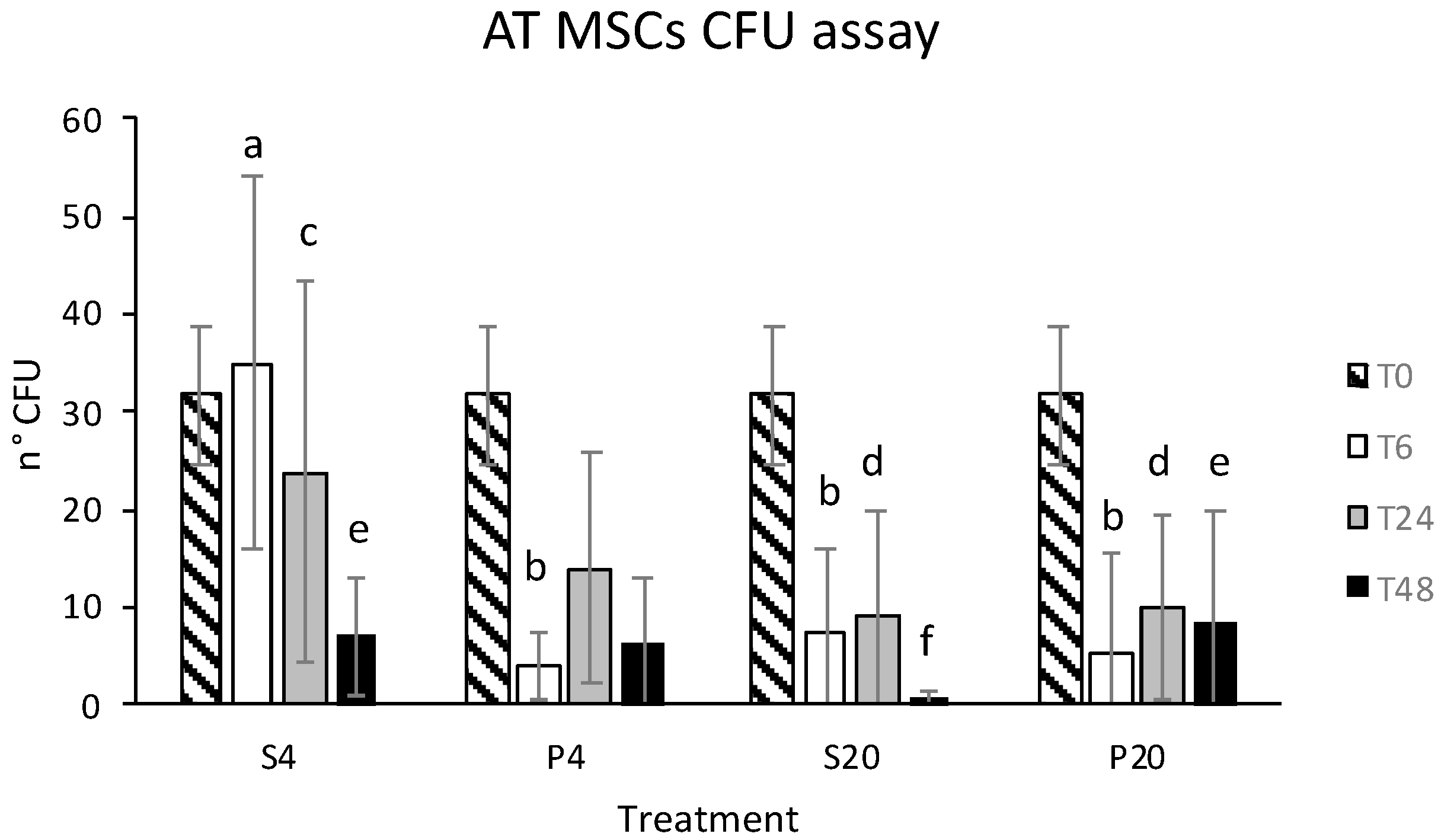
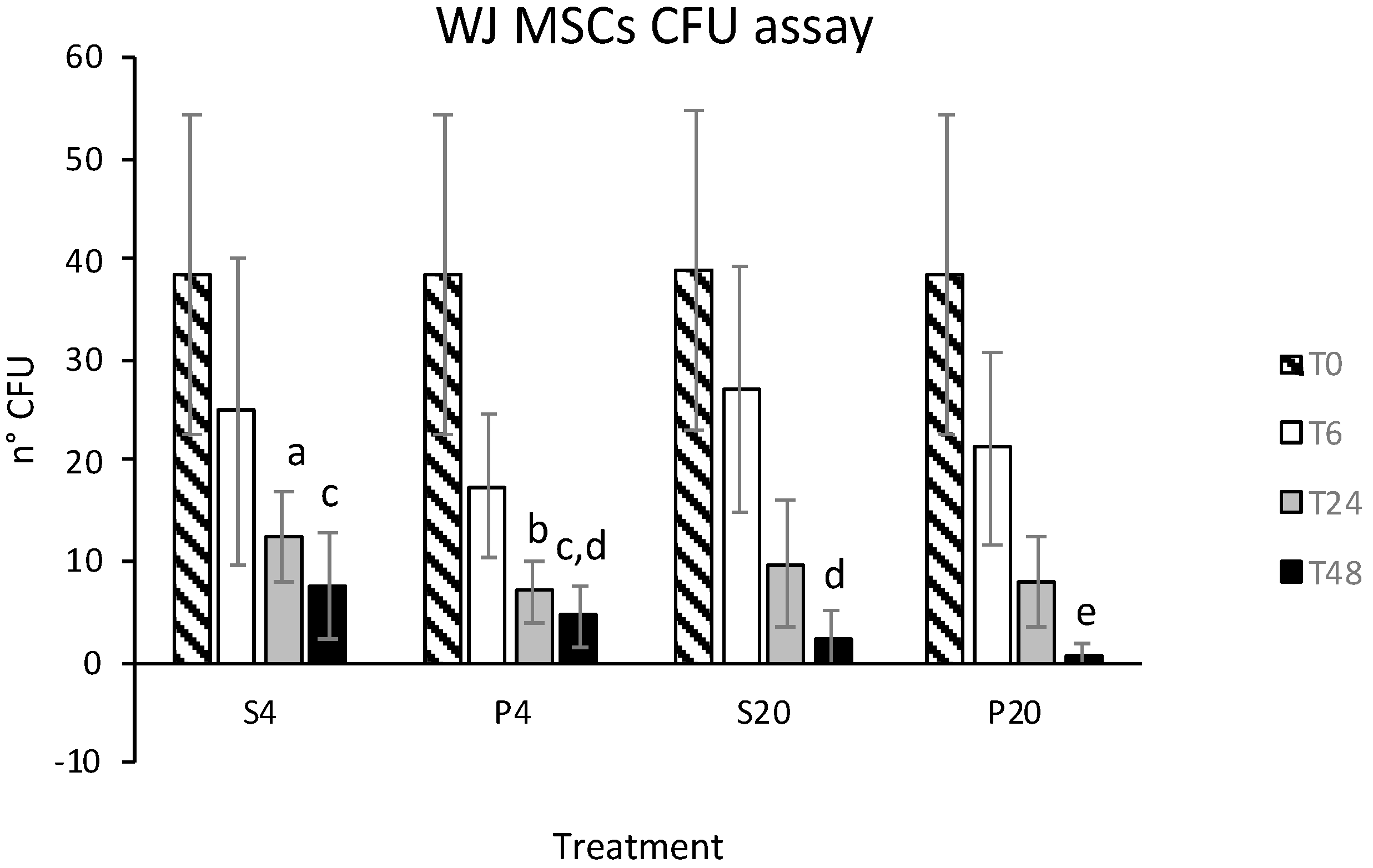
3.1. Vitality and CFU Assay
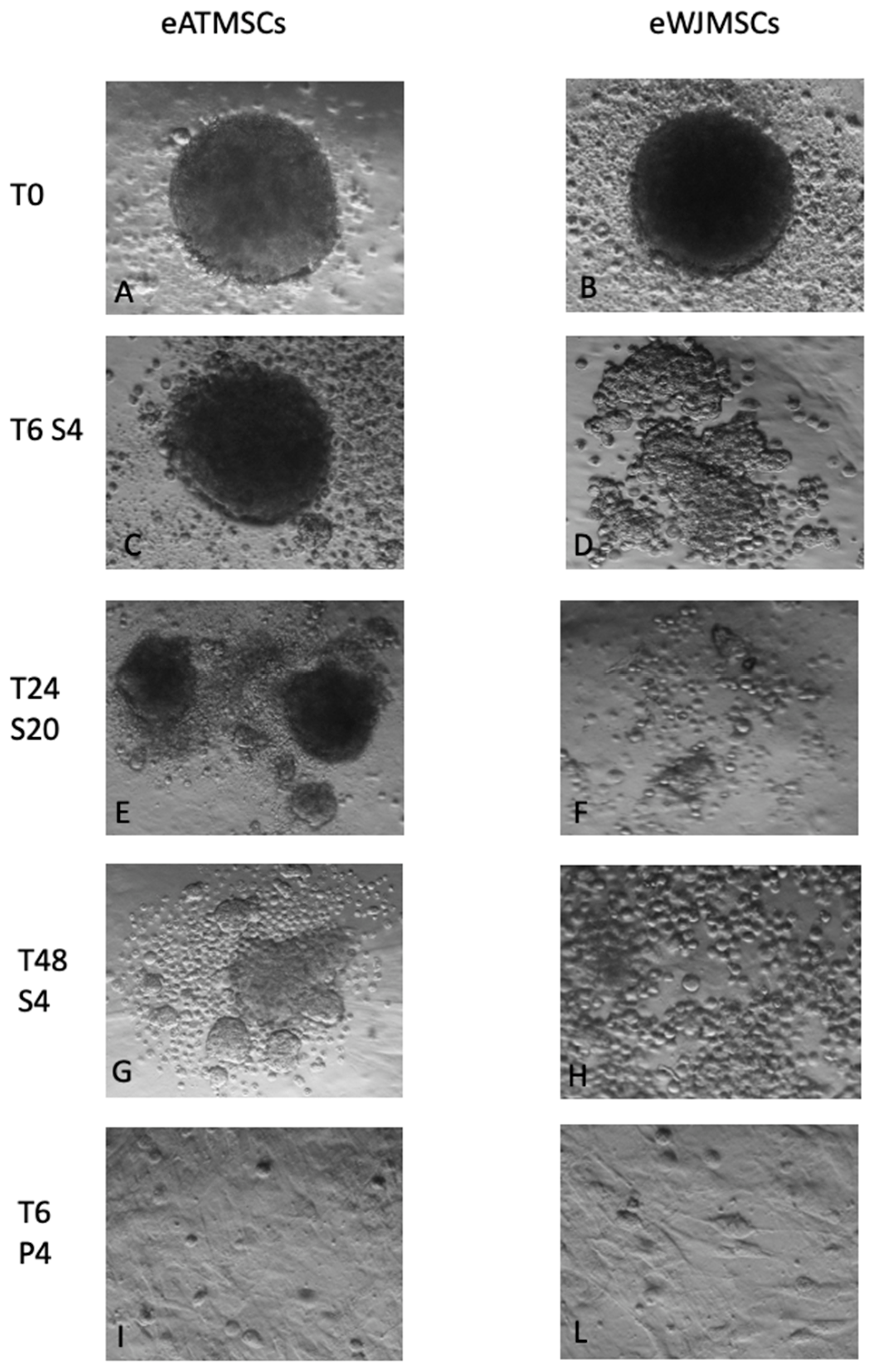
3.2. Spheroid Formation Assays
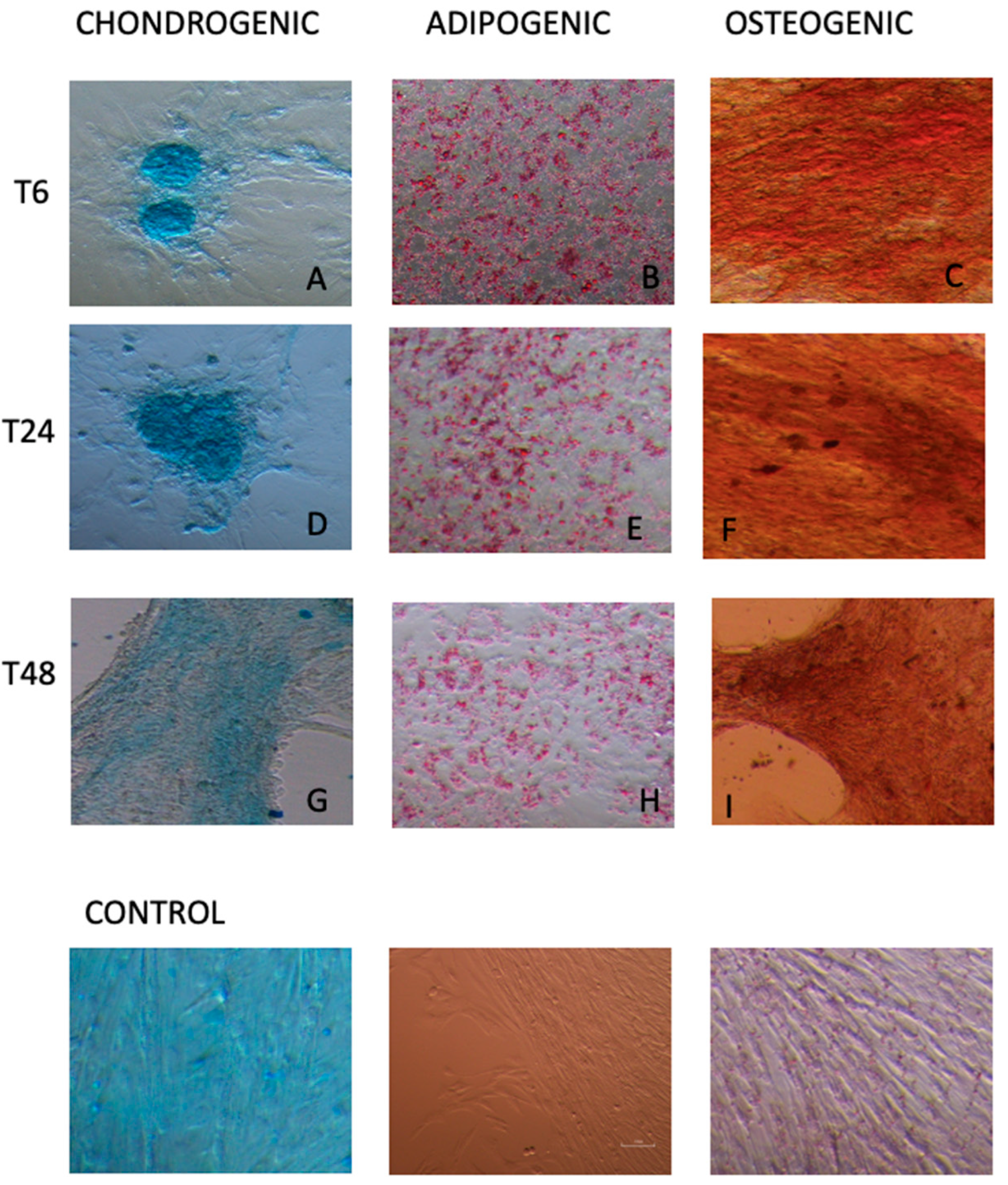
3.3. Multi Lineage In Vitro Differentiation and RT-PCR
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AF | Amniotic Fluid |
| eAFMSCs | Equine Amniotic Fluid Mesenchymal Stromal Cells |
| AM | Amniotic Membrane |
| AT | Adipose Tissue |
| eATMSCs | Equine Adipose Tissue Mesenchymal Stromal Cells |
| BM | Bone Marrow |
| eBMMSCs | Equine Bone Marrow Mesenchymal Stromal cells |
| CD | Cluster Designation |
| DMEM | Dulbecco’s Modified Eagle’s Medium |
| DMSO | dimethyl sulfoxide |
| EV | Extracellular Vesicles |
| F cells | Fibroblastic cells |
| FBS | Fetal Bovine Serum |
| Hrs | Hours |
| IL | Interleukin |
| ISCT | International Society For Cytotherapy |
| MHC | Major Histocompatibility Complex |
| MSCs | Mesenchymal Stromal Cells |
| NSAIDs | Non-Steroidal Anti-Inflammatory Drug |
| P | In vitro culture Passage |
| DPBS | Dulbecco’s Phosphate Buffer Solution |
| ePBMSCs | Equine Peripheral Blood Mesenchymal Stromal Cells |
| ePMSCs | Equine Placenta Mesenchymal Stromal Cells |
| PRP | Platelet Rich Plasma |
| RT | Room Temperature |
| RT-PCR | Real Time Polymerase Chain reaction |
| TGF | Transforming Growth Factor |
| UC | Umbilical Cord |
| UCB | Umbilical Cord Blood |
| eUCBMSCs | Equine Umbilical Cord Blood Mesenchymal Stromal Cells |
| VEGF | Vascular Endothelial Growth Factor |
| WJ | Wharton’s Jelly |
| eWJMSCs | Equine Wharton’s Jelly Mesenchymal Stromal Cells |
References
- Meirelles, L.d.S.; Fontes, A.M.; Covas, D.T.; Caplan, A.I. Mechanisms Involved in the Therapeutic Properties of Mesenchymal Stem Cells. Cytokine Growth Factor Rev. 2009, 20, 419–427. [Google Scholar] [CrossRef]
- Barrachina, L.; Romero, A.; Zaragoza, P.; Rodellar, C.; Vázquez, F.J. Practical Considerations for Clinical Use of Mesenchymal Stem Cells: From the Laboratory to the Horse. Vet. J. Lond. Engl. 1997 2018, 238, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Iacono, E.; Merlo, B.; Romagnoli, N.; Rossi, B.; Ricci, F.; Spadari, A. Equine Bone Marrow and Adipose Tissue Mesenchymal Stem Cells: Cytofluorimetric Characterization, In Vitro Differentiation, and Clinical Application. J. Equine Vet. Sci. 2015, 35, 130–140. [Google Scholar] [CrossRef]
- Colleoni, S.; Bottani, E.; Tessaro, I.; Mari, G.; Merlo, B.; Romagnoli, N.; Spadari, A.; Galli, C.; Lazzari, G. Isolation, Growth and Differentiation of Equine Mesenchymal Stem Cells: Effect of Donor, Source, Amount of Tissue and Supplementation with Basic Fibroblast Growth Factor. Vet. Res. Commun. 2009, 33, 811–821. [Google Scholar] [CrossRef] [PubMed]
- Lange-Consiglio, A.; Tassan, S.; Corradetti, B.; Meucci, A.; Perego, R.; Bizzaro, D.; Cremonesi, F. Investigating the Efficacy of Amnion-Derived Compared with Bone Marrow-Derived Mesenchymal Stromal Cells in Equine Tendon and Ligament Injuries. Cytotherapy 2013, 15, 1011–1020. [Google Scholar] [CrossRef]
- Iacono, E.; Merlo, B.; Pirrone, A.; Antonelli, C.; Brunori, L.; Romagnoli, N.; Castagnetti, C. Effects of Mesenchymal Stem Cells Isolated from Amniotic Fluid and Platelet-Rich Plasma Gel on Severe Decubitus Ulcers in a Septic Neonatal Foal. Res. Vet. Sci. 2012, 93, 1439–1440. [Google Scholar] [CrossRef] [PubMed]
- Corradetti, B.; Correani, A.; Romaldini, A.; Marini, M.G.; Bizzaro, D.; Perrini, C.; Cremonesi, F.; Lange-Consiglio, A. Amniotic Membrane-Derived Mesenchymal Cells and Their Conditioned Media: Potential Candidates for Uterine Regenerative Therapy in the Horse. PLoS ONE 2014, 9, e111324. [Google Scholar] [CrossRef] [Green Version]
- Lanci, A.; Merlo, B.; Mariella, J.; Castagnetti, C.; Iacono, E. Heterologous Wharton’s Jelly Derived Mesenchymal Stem Cells Application on a Large Chronic Skin Wound in a 6-Month-Old Filly. Front. Vet. Sci. 2019, 6, 9. [Google Scholar] [CrossRef] [Green Version]
- Lange-Consiglio, A.; Funghi, F.; Cantile, C.; Idda, A.; Cremonesi, F.; Riccaboni, P. Case Report: Use of Amniotic Microvesicles for Regenerative Medicine Treatment of a Mare With Chronic Endometritis. Front. Vet. Sci. 2020, 7, 347. [Google Scholar] [CrossRef]
- De Coppi, P.; Bartsch, G.; Siddiqui, M.M.; Xu, T.; Santos, C.C.; Perin, L.; Mostoslavsky, G.; Serre, A.C.; Snyder, E.Y.; Yoo, J.J.; et al. Isolation of Amniotic Stem Cell Lines with Potential for Therapy. Nat. Biotechnol. 2007, 25, 100–106. [Google Scholar] [CrossRef]
- Merlo, B.; Teti, G.; Lanci, A.; Burk, J.; Mazzotti, E.; Falconi, M.; Iacono, E. Comparison between Adult and Foetal Adnexa Derived Equine Post-Natal Mesenchymal Stem Cells. BMC Vet. Res. 2019, 15, 277. [Google Scholar] [CrossRef]
- Iacono, E.; Pascucci, L.; Rossi, B.; Bazzucchi, C.; Lanci, A.; Ceccoli, M.; Merlo, B. Ultrastructural Characteristics and Immune Profile of Equine MSCs from Fetal Adnexa. Reprod. Camb. Engl. 2017, 154, 509–519. [Google Scholar] [CrossRef] [Green Version]
- Becerra, P.; Valdés Vázquez, M.A.; Dudhia, J.; Fiske-Jackson, A.R.; Neves, F.; Hartman, N.G.; Smith, R.K.W. Distribution of Injected Technetium(99m)-Labeled Mesenchymal Stem Cells in Horses with Naturally Occurring Tendinopathy. J. Orthop. Res. Off. Publ. Orthop. Res. Soc. 2013, 31, 1096–1102. [Google Scholar] [CrossRef]
- Garvican, E.R.; Cree, S.; Bull, L.; Smith, R.K.; Dudhia, J. Viability of Equine Mesenchymal Stem Cells during Transport and Implantation. Stem Cell Res. Ther. 2014, 5, 1. [Google Scholar] [CrossRef] [Green Version]
- Bronzini, I.; Patruno, M.; Iacopetti, I.; Martinello, T. Influence of Temperature, Time and Different Media on Mesenchymal Stromal Cells Shipped for Clinical Application. Vet. J. Lond. Engl. 1997 2012, 194, 121–123. [Google Scholar] [CrossRef]
- Mercati, F.; Pascucci, L.; Curina, G.; Scocco, P.; Tardella, F.M.; Dall’aglio, C.; Marini, C.; Ceccarelli, P. Evaluation of Storage Conditions on Equine Adipose Tissue-Derived Multipotent Mesenchymal Stromal Cells. Vet. J. Lond. Engl. 1997 2014, 200, 339–342. [Google Scholar] [CrossRef]
- Espina, M.; Jülke, H.; Brehm, W.; Ribitsch, I.; Winter, K.; Delling, U. Evaluation of Transport Conditions for Autologous Bone Marrow-Derived Mesenchymal Stromal Cells for Therapeutic Application in Horses. PeerJ 2016, 4, e1773. [Google Scholar] [CrossRef] [Green Version]
- Merlo, B.; Pirondi, S.; Iacono, E.; Rossi, B.; Ricci, F.; Mari, G. Viability, in vitro differentiation and molecular characterization of equine adipose tissue-derived mesenchymal stem cells cryopreserved in serum and serum-free medium. Cryo Lett. 2016, 37, 243–252. [Google Scholar]
- Mizuno, H.; Hyakusoku, H. Mesengenic Potential and Future Clinical Perspective of Human Processed Lipoaspirate Cells. J. Nippon Med. Sch. Nippon Ika Daigaku Zasshi 2003, 70, 300–306. [Google Scholar] [CrossRef] [Green Version]
- Iacono, E.; Brunori, L.; Pirrone, A.; Pagliaro, P.P.; Ricci, F.; Tazzari, P.L.; Merlo, B. Isolation, Characterization and Differentiation of Mesenchymal Stem Cells from Amniotic Fluid, Umbilical Cord Blood and Wharton’s Jelly in the Horse. Reprod. Camb. Engl. 2012, 143, 455–468. [Google Scholar] [CrossRef] [Green Version]
- Mohanty, N.; Gulati, B.R.; Kumar, R.; Gera, S.; Kumar, P.; Somasundaram, R.K.; Kumar, S. Immunophenotypic Characterization and Tenogenic Differentiation of Mesenchymal Stromal Cells Isolated from Equine Umbilical Cord Blood. In Vitro Cell. Dev. Biol. Anim. 2014, 50, 538–548. [Google Scholar] [CrossRef] [PubMed]
- Desmarais, J.A.; Demers, S.-P.; Suzuki, J.; Laflamme, S.; Vincent, P.; Laverty, S.; Smith, L.C. Trophoblast Stem Cell Marker Gene Expression in Inner Cell Mass-Derived Cells from Parthenogenetic Equine Embryos. Reprod. Camb. Engl. 2011, 141, 321–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal Criteria for Defining Multipotent Mesenchymal Stromal Cells. The International Society for Cellular Therapy Position Statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Ling, L.; Camilleri, E.T.; Helledie, T.; Samsonraj, R.M.; Titmarsh, D.M.; Chua, R.J.; Dreesen, O.; Dombrowski, C.; Rider, D.A.; Galindo, M.; et al. Effect of Heparin on the Biological Properties and Molecular Signature of Human Mesenchymal Stem Cells. Gene 2016, 576, 292–303. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Adipogenic | Chondrogenic | Osteogenic |
|---|---|---|
| DMEM | DMEM | DMEM |
| 10% FBS | 1% FBS | 10% Rabbit Serum |
| 0.5 mM IBMX (removed after 3 days) | 6.25 μg/mL insulin | 50 μM AA2P |
| 1 μM DXM (removed after 6 days) | 50 nM AA2P | 0.1 μM DXM |
| 10 μg/mL insulin | 0.1 μM DXM | 10 mM BGP |
| 0.1 mM indomethacin | 10 ng/mL hTGF-β1 |
| Primers | References | Sequences (5′→3′) | bp |
|---|---|---|---|
| MSC marker | |||
| CD90 | [21] | FW: TGCGAACTCCGCCTCTCT | 93 |
| RW: GCTTATGCCCTCGCACTTG | |||
| Ematopoietic markers | |||
| CD45 | [21] | FW: TGATTCCCAGAAATGACATGTA | 101 |
| RW: ACATTTTGGGCTTGTCCTTAAC | |||
| Housekeeping | |||
| GAPDH | [22] | FW: GTCCATGCCATCACTGCCAC | 262 |
| RW: CCTGCTTCACCACCTTCTTG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iacono, E.; Lanci, A.; Gugole, P.; Merlo, B. Shipping Temperature, Time and Media Effects on Equine Wharton’s Jelly and Adipose Tissue Derived Mesenchymal Stromal Cells Characteristics. Animals 2022, 12, 1967. https://doi.org/10.3390/ani12151967
Iacono E, Lanci A, Gugole P, Merlo B. Shipping Temperature, Time and Media Effects on Equine Wharton’s Jelly and Adipose Tissue Derived Mesenchymal Stromal Cells Characteristics. Animals. 2022; 12(15):1967. https://doi.org/10.3390/ani12151967
Chicago/Turabian StyleIacono, Eleonora, Aliai Lanci, Penelope Gugole, and Barbara Merlo. 2022. "Shipping Temperature, Time and Media Effects on Equine Wharton’s Jelly and Adipose Tissue Derived Mesenchymal Stromal Cells Characteristics" Animals 12, no. 15: 1967. https://doi.org/10.3390/ani12151967