Mast Cell Population and Histamine Content in Hypothyroid Rat Tissues


 ,
, {kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Experiments
2.2. T3 and TSH Serum Determinations
2.3. Histochemistry and Ultrastructure
2.4. Morphometric Analysis
2.5. Histamine Determination
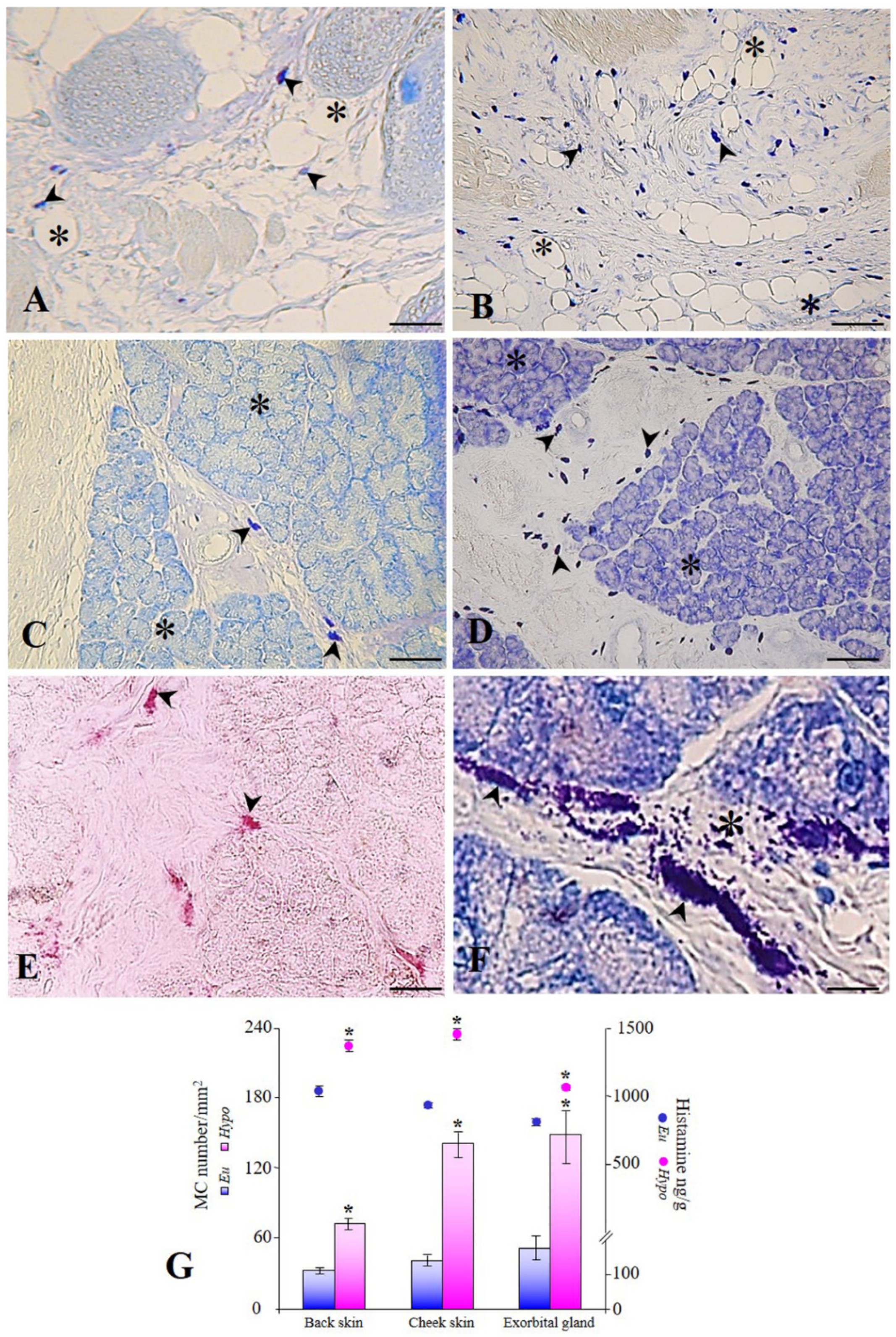
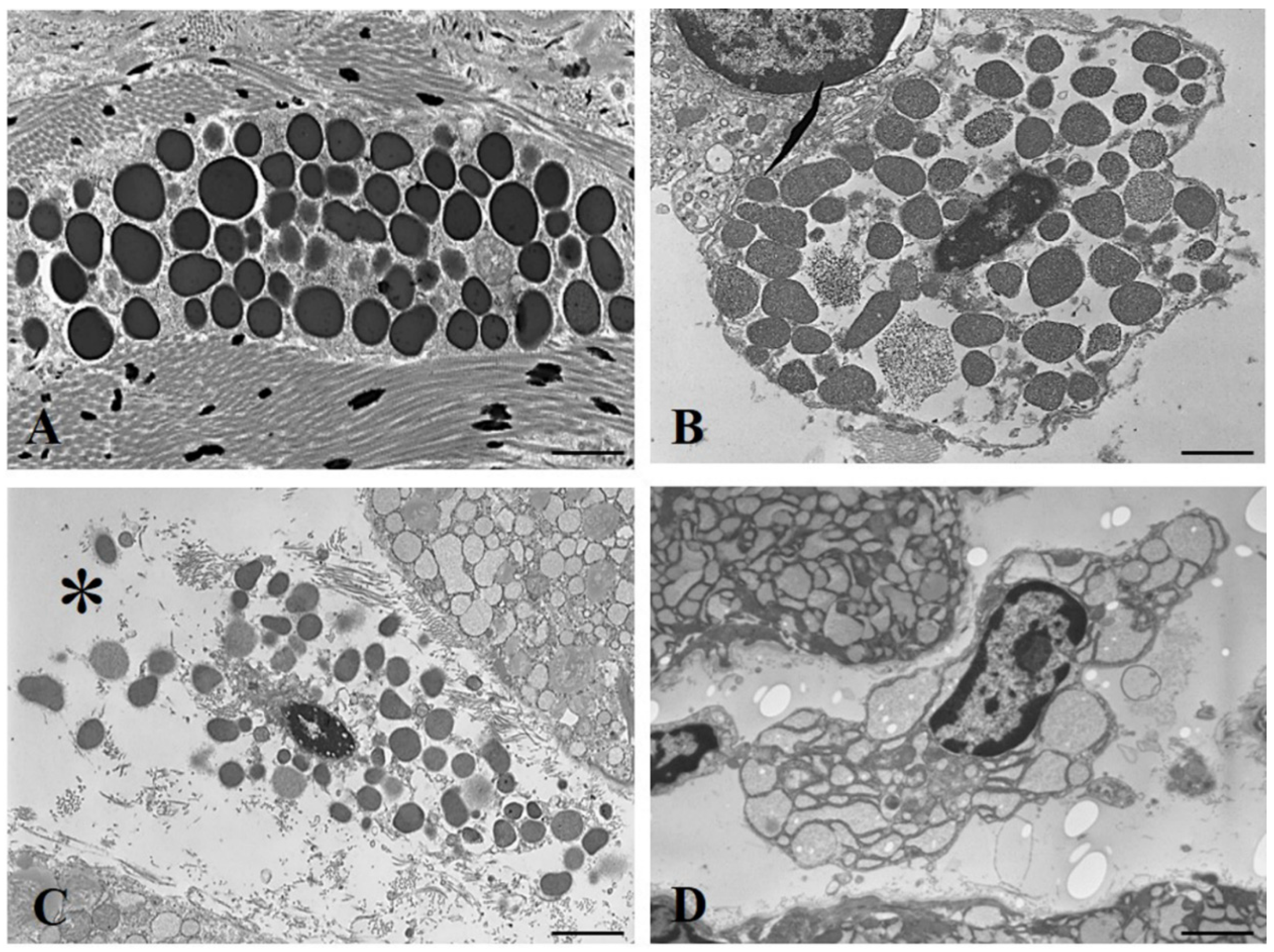
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Galli, S.J.; Tsai, M. IgE and mast cells in allergic disease. Nat. Med. 2012, 18, 693–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- da Silva, E.Z.; Jamur, M.C.; Oliver, C. Mast cell function: A new vision of an old cell. J. Histochem. Cytochem. 2014, 62, 698–738. [Google Scholar] [CrossRef] [PubMed]
- Varricchi, G.; de Paulis, A.; Marone, G.; Galli, S.J. Future Needs in Mast Cell Biology. Int. J. Mol. Sci. 2019, 20, 4397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitamura, Y. Heterogeneity of mast cells and phenotypic change between subpopulations. Ann. Rev. Immunol. 1989, 7, 59–76. [Google Scholar] [CrossRef]
- Galli, S.J.; Nakae, S.; Tsai, M. Mast cells in the development of adaptive immune responses. Nat. Immunol. 2005, 6, 135–142. [Google Scholar] [CrossRef]
- Metcalfe, D.D.; Boyce, J.A. Mast cell biology in evolution. J. Allergy Clin. Immunol. 2006, 117, 1227–1229. [Google Scholar] [CrossRef]
- Baccari, G.C.; Pinelli, C.; Santillo, A.; Minucci, S.; Rastogi, R.K. Mast cells in nonmammalian vertebrates: An overview. Int. Rev. Cell. Mol. Biol. 2011, 290, 1–53. [Google Scholar] [CrossRef]
- Beil, W.J.; Schulz, M.; Wefelmeyer, U. Mast cell granule composition and tissue location—A close correlation. Histol. Histopathol. 2000, 15, 937–946. [Google Scholar] [CrossRef]
- Nakamura, T. The roles of lipid mediators in type I hypersensitivity. J. Pharmacol. Sci. 2021, 147, 126–131. [Google Scholar] [CrossRef]
- Tainsh, K.R.; Pearce, F.L. Mast Cell Heterogeneity: Evidence that Mast Cells Isolated from Various Connective Tissue Locations in the Rat Display Markedly Graded Phenotypes. Int. Arch. Allergy Immunol. 1992, 98, 26–34. [Google Scholar] [CrossRef]
- Bienenstock, J. An update on mast cell heterogeneity. J. Allergy Clin. Immunol. 1988, 81, 763–769. [Google Scholar] [CrossRef]
- Akula, S.; Paivandy, A.; Fu, Z.; Thorpe, M.; Pejler, G.; Hellman, L. Quantitative In-Depth Analysis of the Mouse Mast Cell Transcriptome Reveals Organ-Specific Mast Cell Heterogeneity. Cells 2020, 9, 211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabria, J.; Ferrer, I.; Toledo, A.; Sentis, M.; Blanco, I. Effects of altered thyroid function on histamine levels and mast cell number in neonatal rat brain. J. Pharmacol. Exp. Ther. 1987, 240, 612–616. [Google Scholar] [PubMed]
- Siebler, T.; Robson, H.; Bromley, M.; Stevens, D.A.; Shalet, S.M.; Williams, G.R. Thyroid status affects number and localization of thyroid hormone receptor expressing mast cells in bone marrow. Bone 2002, 30, 259–266. [Google Scholar] [CrossRef]
- Artantaş, Ş.; Gül, Ü.; Kılıç, A.; Güler, S. Skin findings in thyroid diseases. Eur. J. Intern. Med. 2009, 20, 158–161. [Google Scholar] [CrossRef]
- Landucci, E.; Laurino, A.; Cinci, L.; Gencarelli, M.; Raimondi, L. Thyroid Hormone, Thyroid Hormone Metabolites and Mast Cells: A Less Explored Issue. Front. Cell. Neurosci. 2019, 13, 79. [Google Scholar] [CrossRef]
- Monteforte, R.; Santillo, A.; Lanni, A.; D’Aniello, S.; Baccari, G.C. Morphological and biochemical changes in the Harderian gland of hypothyroid rats. J. Exp. Biol. 2008, 211, 606–612. [Google Scholar] [CrossRef] [Green Version]
- Ribatti, D. The Staining of Mast Cells: A Historical Overview. Int. Arch. Allergy Immunol. 2018, 176, 55–60. [Google Scholar] [CrossRef]
- Baccari, G.C.; De Paulis, A.; Di Matteo, L.; Gentile, M.; Marone, G.; Minucci, S. In situ characterization of mast cells in the frog Rana esculenta. Cell Tissue Res. 1998, 292, 151–162. [Google Scholar] [CrossRef]
- Ferrara, D.; Monteforte, R.; Baccari, G.C.; Minucci, S.; Chieffi, G. Androgen and estrogen receptors expression in the rat exorbital lacrimal gland in relation to “harderianization”. J. Exp. Zool. Part A Comp. Exp. Biol. 2004, 301, 297–306. [Google Scholar] [CrossRef]
- Wilhelm, M.; King, B.; Silverman, A.J.; Silver, R. Gonadal steroids regulate the number and activational state of mast cells in the medial habenula. Endocrinology 2000, 141, 1178–1186. [Google Scholar] [CrossRef]
- Thangam, E.B.; Jemima, E.A.; Singh, H.; Baig, M.S.; Khan, M.; Mathias, C.B.; Church, M.K.; Saluja, R. The Role of Histamine and Histamine Receptors in Mast Cell-Mediated Allergy and Inflammation: The Hunt for New Therapeutic Targets. Front. Immunol. 2018, 9, 1873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Castelán, J.; Corona-Pérez, A.; Nicolás-Toledo, L.; Martínez-Gómez, M.; Castelán, F.; Cuevas-Romero, E. Hypothyroidism Induces a Moderate Steatohepatitis Accompanied by Liver Regeneration, Mast Cells Infiltration, and Changes in the Expression of the Farnesoid X Receptor. Exp. Clin. Endocrinol. Diabetes 2017, 125, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Csaba, G.; Pállinger, É. Thyrotropic hormone (TSH) regulation of triiodothyronine (T3) concentration in immune cells. Inflamm. Res. 2009, 58, 151–154. [Google Scholar] [CrossRef]
- Clayton, J.A.; Masuoka, D.T. TSH-Induced Mobilization of Serotonin from Perivascular Mast Cells in the Rat Thyroid. Endocrinology 1968, 83, 263–271. [Google Scholar] [CrossRef]
- Ericson, L.E.; Håkanson, R.; Melander, A.; Owman, C.; Sundler, F. TSH-Induced Release of 5-Hydroxytryptamine and Histamine from Rat Thyroid Mast Cells. Endocrinology 1972, 90, 795–801. [Google Scholar] [CrossRef]
- Melander, A.; Sundler, F. Significance of Thyroid Mast Cells in Thyroid Hormone Secretion. Endocrinology 1972, 90, 802–807. [Google Scholar] [CrossRef] [PubMed]
- Wynford-Thomas, D.; Stringer, B.M.J. Mast cell hyperplasia during goitrogen—Induced thyroid growth—A quantitative study. Eur. J. Endocrinol. 1982, 101, 365–370. [Google Scholar] [CrossRef]
- Monteforte, R.; Pinelli, C.; Santillo, A.; Rastogi, R.K.; Polese, G.; Baccari, G.C. Mast cell population in the frog brain: Distribution and influence of thyroid status. J. Exp. Biol. 2010, 213, 1762–1770. [Google Scholar] [CrossRef] [Green Version]
- Baccari, G.C.; Minucci, S.; Marmorino, C.; Izzo, I.V. Number of mast cells in the harderian gland of the green frog, Rana esculenta: The annual cycle and its relation to environmental and hormonal factors. J. Anat. 1991, 179, 75–83. [Google Scholar]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chieffi Baccari, G.; Falvo, S.; Lanni, A.; Di Fiore, M.M.; Cioffi, F.; Santillo, A. Mast Cell Population and Histamine Content in Hypothyroid Rat Tissues. Animals 2022, 12, 1840. https://doi.org/10.3390/ani12141840
Chieffi Baccari G, Falvo S, Lanni A, Di Fiore MM, Cioffi F, Santillo A. Mast Cell Population and Histamine Content in Hypothyroid Rat Tissues. Animals. 2022; 12(14):1840. https://doi.org/10.3390/ani12141840
Chicago/Turabian StyleChieffi Baccari, Gabriella, Sara Falvo, Antonia Lanni, Maria Maddalena Di Fiore, Federica Cioffi, and Alessandra Santillo. 2022. "Mast Cell Population and Histamine Content in Hypothyroid Rat Tissues" Animals 12, no. 14: 1840. https://doi.org/10.3390/ani12141840