
Identification of Body Size Determination Related Candidate Genes in Domestic Pig Using Genome-Wide Selection Signal Analysis

 ,
,  ,
, {kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
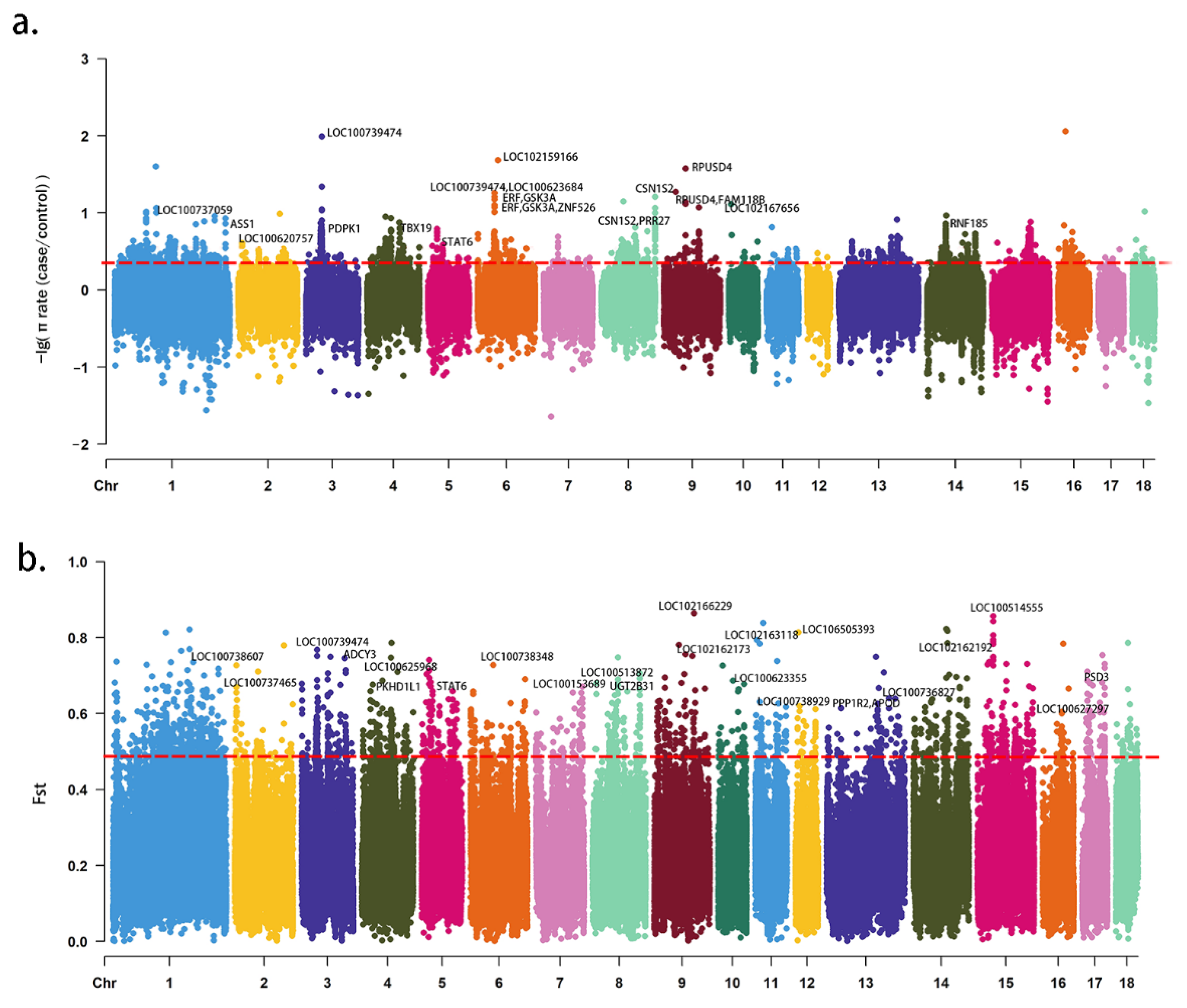
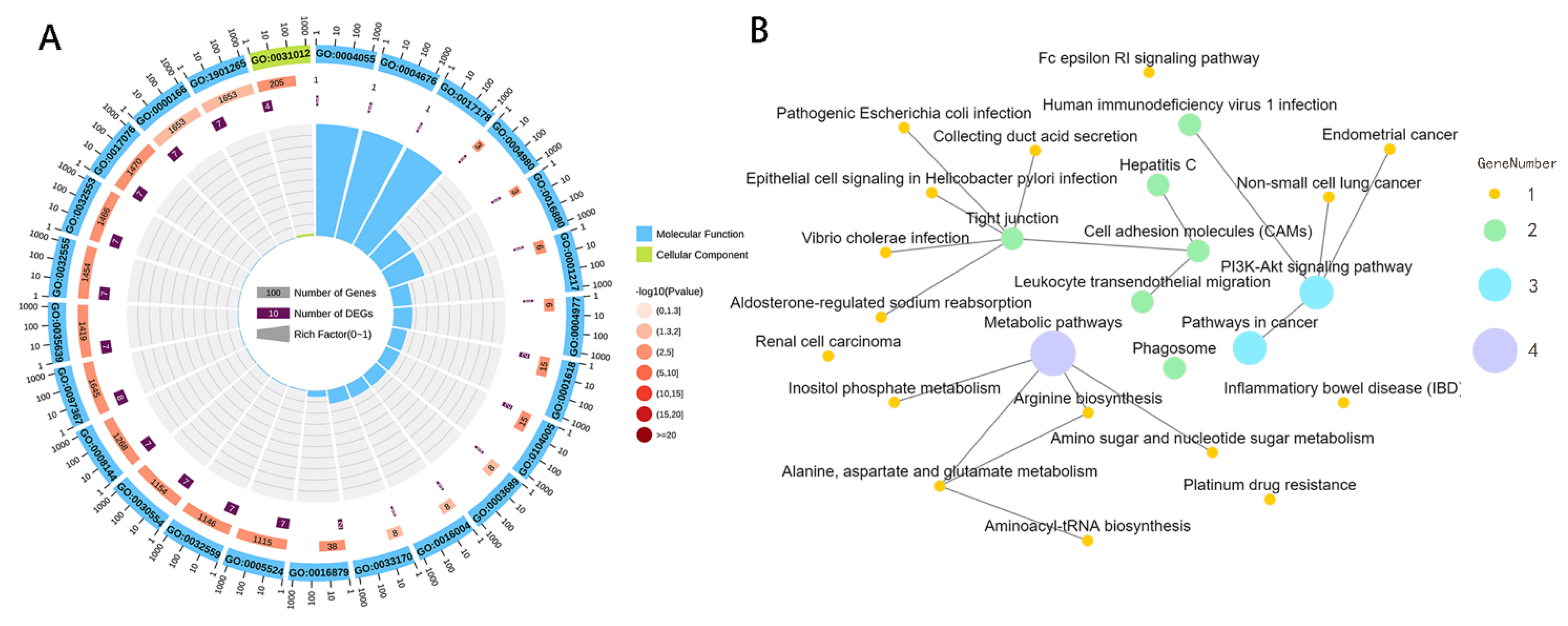
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Sutherland, A.; Myburgh, J.; Steyn, M.; Becker, P.J. The effect of body size on the rate of decomposition in a temperate region of South Africa. Forensic. Sci. Int. 2013, 231, 258–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hannon, B.M.; Murphy, M.R. Toward a theory of energetically optimal body size in growing animals. J. Anim. Sci. 2016, 94, 2532–2541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Zhuang, Y.; Ji, H.; Teng, G. Pig Weight and Body Size Estimation Using a Multiple Output Regression Convolutional Neural Network: A Fast and Fully Automatic Method. Sensors 2021, 21, 3218. [Google Scholar] [CrossRef] [PubMed]
- National Commission on Livestock and Poultry Genetic Resources. Animal Genetic Re-sources in China: Pigs, 1st ed.; China Agricuture Press: Beijing, China, 2008. [Google Scholar]
- Tynes, V.V. Miniature Pet Pig Behavioral Medicine. Vet. Clin. N. Am. Exot. Anim. Pract. 2021, 24, 63–86. [Google Scholar] [CrossRef] [PubMed]
- Musial, F.; Kowalski, A.; Treiber, A.; Enck, P. Temporal characteristics of feeding behavior in the Munich miniature pig. Physiol. Behav. 2006, 87, 206–218. [Google Scholar] [CrossRef] [PubMed]
- Swindle, M.M.; Makin, A.; Herron, A.J.; Clubb, F.J.; Frazier, K.S. Swine as models in biomedical research and toxicology testing. Vet. Pathol. 2012, 49, 344–356. [Google Scholar] [CrossRef]
- Ryan, M.C.; Kochunov, P.; Sherman, P.M.; Rowland, L.M.; Wijtenburg, S.A.; Acheson, A.; Hong, L.E.; Sladky, J.; McGuire, S. Miniature pig magnetic resonance spectroscopy model of normal adolescent brain development. J. Neurosci. Methods 2018, 308, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Stricker-Krongrad, A.; Shoemake, C.R.; Bouchard, G.F. The Miniature Swine as a Model in Experimental and Translational Medicine. Toxicol. Pathol. 2016, 44, 612–623. [Google Scholar] [CrossRef] [Green Version]
- Van Metre, D.C.; Angelos, S.M. Miniature pigs. Vet. Clin. N. Am. Exot. Anim. Pract. 1999, 2, 519–537. [Google Scholar] [CrossRef]
- Willens, S.; Cox, D.M.; Braue, E.H.; Myers, T.M.; Wegner, M.D. Novel technique for retroperitoneal implantation of telemetry transmitters for physiologic monitoring in Gottingen minipigs (Sus scrofa ssp. domesticus). Comp. Med. 2014, 64, 464–470. [Google Scholar]
- Saxena, A.; Hastings, N.B.; Sun, W.; Dabisch, P.A.; Hulet, S.W.; Jakubowski, E.M.; Mioduszewski, R.J.; Doctor, B.P. Prophylaxis with human serum butyrylcholinesterase protects Gottingen minipigs exposed to a lethal high-dose of sarin vapor. Chem. Biol. Interact. 2015, 238, 161–169. [Google Scholar] [CrossRef] [Green Version]
- Hey, A.; Hill, M.; Calonder, C.; DeLise, A.M.; Grossi, A.B.; Pique, C.; Marsden, E. "Simulect" as a model compound for assessing placental transfer of monoclonal antibodies in minipigs. Reprod. Toxicol. 2020, 91, 142–146. [Google Scholar] [CrossRef]
- Wang, D.; Xu, X.; Zhao, M.; Wang, X. Accelerated miniature swine models of advanced atherosclerosis: A review based on morphology. Cardiovasc. Pathol. 2020, 49, 107241. [Google Scholar] [CrossRef]
- Zhao, Y.; Xiang, L.; Liu, Y.; Niu, M.; Yuan, J.; Chen, H. Atherosclerosis Induced by a High-Cholesterol and High-Fat Diet in the Inbred Strain of the Wuzhishan Miniature Pig. Anim. Biotechnol. 2018, 29, 110–118. [Google Scholar] [CrossRef]
- Niu, M.; Liu, Y.; Xiang, L.; Zhao, Y.; Yuan, J.; Jia, Y.; Dai, X.; Chen, H. Long-term case study of a Wuzhishan miniature pig with diabetes. Anim. Models Exp. Med. 2020, 3, 22–31. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Huang, Y.; Wang, M.; Guo, Y.; Liang, J.; Yang, X.; Qi, W.; Wu, Y.; Si, J.; Zhu, S.; et al. Development and Genome Sequencing of a Laboratory-Inbred Miniature Pig Facilitates Study of Human Diabetic Disease. iScience 2019, 19, 162–176. [Google Scholar] [CrossRef] [Green Version]
- Dolezalova, D.; Hruska-Plochan, M.; Bjarkam, C.R.; Sorensen, J.C.; Cunningham, M.; Weingarten, D.; Ciacci, J.D.; Juhas, S.; Juhasova, J.; Motlik, J.; et al. Pig models of neurodegenerative disorders: Utilization in cell replacement-based preclinical safety and efficacy studies. J. Comp. Neurol. 2014, 522, 2784–2801. [Google Scholar] [CrossRef]
- Hoffe, B.; Holahan, M.R. The Use of Pigs as a Translational Model for Studying Neurodegenerative Diseases. Front. Physiol. 2019, 10, 838. [Google Scholar] [CrossRef]
- Xiaokai, L.; Quankui, H.; Lingki, F.; Yafeng, G.; Jing, L.; Ganqiu, L. Sequence and expression differences of BMP2 and, FGFR3 genes in Guangxi Bama mini pig and Landrace pig. Guangxi Agric. Sci. 2021, 52, 1709–1718. [Google Scholar] [CrossRef]
- Wanbo, L.; Yaling, Z.; Huashui, A.; Tianfu, G. Identifying signatures of Selection Related to Small Body Size in Pigs. Chin. J. Anim. Vet. Sci. 2016, 47, 1977–1985. [Google Scholar] [CrossRef]
- Ornitz, D.M.; Legeai-Mallet, L. Achondroplasia: Development, pathogenesis, and therapy. Dev. Dyn. 2017, 246, 291–309. [Google Scholar] [CrossRef] [Green Version]
- Ijiri, M.; Lai, Y.C.; Kawaguchi, H.; Fujimoto, Y.; Miura, N.; Matsuo, T.; Tanimoto, A. NR6A1 Allelic Frequencies as an Index for both Miniaturizing and Increasing Pig Body Size. In Vivo 2021, 35, 163–167. [Google Scholar] [CrossRef]
- Kim, H.; Song, K.D.; Kim, H.J.; Park, W.; Kim, J.; Lee, T.; Shin, D.H.; Kwak, W.; Kwon, Y.J.; Sung, S.; et al. Exploring the genetic signature of body size in Yucatan miniature pig. PLoS ONE 2015, 10, e121732. [Google Scholar] [CrossRef] [Green Version]
- Reimer, C.; Rubin, C.J.; Sharifi, A.R.; Ha, N.T.; Weigend, S.; Waldmann, K.H.; Distl, O.; Pant, S.D.; Fredholm, M.; Schlather, M.; et al. Analysis of porcine body size variation using re-sequencing data of miniature and large pigs. BMC Genom. 2018, 19, 687. [Google Scholar] [CrossRef]
- Rubin, C.J.; Megens, H.J.; Martinez, B.A.; Maqbool, K.; Sayyab, S.; Schwochow, D.; Wang, C.; Carlborg, O.; Jern, P.; Jorgensen, C.B.; et al. Strong signatures of selection in the domestic pig genome. Proc. Natl. Acad. Sci. USA 2012, 109, 19529–19536. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Tian, S.; Jin, L.; Zhou, G.; Li, Y.; Zhang, Y.; Wang, T.; Yeung, C.K.; Chen, L.; Ma, J.; et al. Genomic analyses identify distinct patterns of selection in domesticated pigs and Tibetan wild boars. Nat. Genet. 2013, 45, 1431–1438. [Google Scholar] [CrossRef]
- Kim, J.; Cho, S.; Caetano-Anolles, K.; Kim, H.; Ryu, Y.C. Genome-wide detection and characterization of positive selection in Korean Native Black Pig from Jeju Island. BMC Genet. 2015, 16, 3. [Google Scholar] [CrossRef] [Green Version]
- Moon, S.; Kim, T.H.; Lee, K.T.; Kwak, W.; Lee, T.; Lee, S.W.; Kim, M.J.; Cho, K.; Kim, N.; Chung, W.H.; et al. A genome-wide scan for signatures of directional selection in domesticated pigs. BMC Genom. 2015, 16, 130. [Google Scholar] [CrossRef] [Green Version]
- Jeong, H.; Song, K.D.; Seo, M.; Caetano-Anolles, K.; Kim, J.; Kwak, W.; Oh, J.D.; Kim, E.; Jeong, D.K.; Cho, S.; et al. Exploring evidence of positive selection reveals genetic basis of meat quality traits in Berkshire pigs through whole genome sequencing. BMC Genet. 2015, 16, 104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Y.; Li, C.; Tang, Q.; Tian, S.; Jin, L.; Chen, J.; Li, M.; Li, C. Genomic analysis reveals selection in Chinese native black pig. Sci. Rep. 2016, 6, 36354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hudson, R.R.; Mand, S.; Maddison, W.P. Estimation of levels of gene flow from DNA sequence data. Genetics 1992, 132, 2. [Google Scholar] [CrossRef] [PubMed]
- Nei, M.; Li, W.H. Mathematical model for studying genetic variation in terms of restriction endonucleases. Proc. Natl. Acad. Sci. USA 1979, 76, 10. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.Y.; Li, A.; Otecko, N.O.; Liu, Y.H.; Irwin, D.M.; Wang, L.; Adeola, A.C.; Zhang, J.; Xie, H.B.; Zhang, Y.P. PigVar: A database of pig variations and positive selection signatures. Database 2017, 2017, bax048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danecek, P.; Auton, A.; Abecasis, G.; Albers, C.A.; Banks, E.; DePristo, M.A.; Handsaker, R.E.; Lunter, G.; Marth, G.T.; Sherry, S.T.; et al. The variant call format and, VCFtools. Bioinformatics 2011, 27, 2156–2158. [Google Scholar] [CrossRef]
- Wang, G.; Li, M.; Zhou, J.; An, X.; Bai, F.; Gao, Y.; Yu, J.; Li, H.; Lei, C.; Dang, R. A novel A > G polymorphism in the intron 2 of TBX3 gene is significantly associated with body size in donkeys. Gene 2021, 785, 145602. [Google Scholar] [CrossRef]
- Gui, L.S.; Zhang, Y.R.; Liu, G.Y.; Zan, L.S. Expression of the SIRT2 gene and its relationship with body size traits in Qinchuan cattle (Bos taurus). Int. J. Mol. Sci. 2015, 16, 2458–2471. [Google Scholar] [CrossRef] [Green Version]
- Metzger, J.; Schrimpf, R.; Philipp, U.; Distl, O. Expression levels of, L.CORL are associated with body size in horses. PLoS ONE 2013, 8, e56497. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Hu, H.; Tian, Y.; Li, J.; Scheben, A.; Zhang, C.; Li, Y.; Wu, J.; Yang, L.; Fan, X.; et al. The Chicken Pan-Genome Reveals Gene Content Variation and a Promoter Region Deletion in IGF2BP1 Affecting Body Size. Mol. Biol. Evol. 2021, 38, 5066–5081. [Google Scholar] [CrossRef]
- Niswender, K.D. Basal insulin: Physiology, pharmacology, and clinical implications. Postgrad. Med. 2011, 123, 17–26. [Google Scholar] [CrossRef]
- Stratton, M. Genome resequencing and genetic variation. Nat. Biotechnol. 2008, 26, 65–66. [Google Scholar] [CrossRef]
- Sharma, R.; Kopchick, J.J.; Puri, V.; Sharma, V.M. Effect of growth hormone on insulin signaling. Mol. Cell Endocrinol. 2020, 518, 111038. [Google Scholar] [CrossRef]
- Gardner, L.B.; Li, Q.; Park, M.S.; Flanagan, W.M.; Semenza, G.L.; Dang, C.V. Hypoxia inhibits G1/S transition through regulation of p27 expression. J. Biol. Chem. 2001, 276, 7919–7926. [Google Scholar] [CrossRef] [Green Version]
- Carmeliet, P.; Dor, Y.; Herbert, J.M.; Fukumura, D.; Brusselmans, K.; Dewerchin, M.; Neeman, M.; Bono, F.; Abramovitch, R.; Maxwell, P.; et al. Role of, H.IF-1alpha in hypoxia-mediated apoptosis, cell proliferation and tumour angiogenesis. Nature 1998, 394, 485–490. [Google Scholar] [CrossRef]
- Biswas, S.; Roy, S.; Banerjee, J.; Hussain, S.R.; Khanna, S.; Meenakshisundaram, G.; Kuppusamy, P.; Friedman, A.; Sen, C.K. Hypoxia inducible microRNA 210 attenuates keratinocyte proliferation and impairs closure in a murine model of ischemic wounds. Proc. Natl. Acad. Sci. USA 2010, 107, 6976–6981. [Google Scholar] [CrossRef] [Green Version]
- Kyun, S.; Yoo, C.; Park, H.Y.; Kim, J.; Lim, K. The Effects of Exogenous Lactate Administration on the, I.GF1/Akt/mTOR Pathway in Rat Skeletal Muscle. Int. J. Environ. Res. Public Health 2020, 17, 7805. [Google Scholar] [CrossRef]
- Aiello, D.; Patel, K.; Lasagna, E. The myostatin gene: An overview of mechanisms of action and its relevance to livestock animals. Anim. Genet. 2018, 49, 505–519. [Google Scholar] [CrossRef] [Green Version]
- McCroskery, S.; Thomas, M.; Maxwell, L.; Sharma, M.; Kambadur, R. Myostatin negatively regulates satellite cell activation and self-renewal. J. Cell Biol. 2003, 162, 1135–1147. [Google Scholar] [CrossRef] [Green Version]
- Hubbi, M.E.; Kshitiz; Gilkes, D.M.; Rey, S.; Wong, C.C.; Luo, W.; Kim, D.H.; Dang, C.V.; Levchenko, A.; Semenza, G.L. A nontranscriptional role for HIF-1alpha as a direct inhibitor of DNA replication. Sci. Signal. 2013, 6, a10. [Google Scholar] [CrossRef] [Green Version]
- Karalaki, M.; Fili, S.; Philippou, A.; Koutsilieris, M. Muscle regeneration: Cellular and molecular events. In Vivo 2009, 23, 779–796. [Google Scholar]
- Heydemann, A.; Ceco, E.; Lim, J.E.; Hadhazy, M.; Ryder, P.; Moran, J.L.; Beier, D.R.; Palmer, A.A.; McNally, E.M. Latent T.GF-beta-binding protein 4 modifies muscular dystrophy in mice. J. Clin. Investig. 2009, 119, 3703–3712. [Google Scholar] [CrossRef] [Green Version]
- McGlory, C.; van Vliet, S.; Stokes, T.; Mittendorfer, B.; Phillips, S.M. The impact of exercise and nutrition on the regulation of skeletal muscle mass. J. Physiol. 2019, 597, 1251–1258. [Google Scholar] [CrossRef] [Green Version]
- Stinckens, A.; Luyten, T.; Bijttebier, J.; Van den Maagdenberg, K.; Dieltiens, D.; Janssens, S.; De Smet, S.; Georges, M.; Buys, N. Characterization of the complete porcine MSTN gene and expression levels in pig breeds differing in muscularity. Anim. Genet. 2008, 39, 586–596. [Google Scholar] [CrossRef] [PubMed]
- Matsakas, A.; Otto, A.; Elashry, M.I.; Brown, S.C.; Patel, K. Altered primary and secondary myogenesis in the myostatin-null mouse. Rejuvenation Res. 2010, 13, 717–727. [Google Scholar] [CrossRef]
- Sternstein, I.; Reissmann, M.; Maj, D.; Bieniek, J.; Brockmann, G.A. A new single nucleotide polymorphism in the rabbit (Oryctolagus cuniculus) myostatin (MSTN) gene is associated with carcass composition traits. Anim. Genet. 2014, 45, 596–599. [Google Scholar] [CrossRef]
- Lamar, K.M.; Bogdanovich, S.; Gardner, B.B.; Gao, Q.Q.; Miller, T.; Earley, J.U.; Hadhazy, M.; Vo, A.H.; Wren, L.; Molkentin, J.D.; et al. Overexpression of Latent TGFbeta Binding Protein 4 in Muscle Ameliorates Muscular Dystrophy through Myostatin and, T.GFbeta. PLoS Genet. 2016, 12, e1006019. [Google Scholar] [CrossRef] [Green Version]
- Thomson, D.M. The Role of AMPK in the Regulation of Skeletal Muscle Size, Hypertrophy, and Regeneration. Int. J. Mol. Sci. 2018, 19, 3125. [Google Scholar] [CrossRef] [Green Version]
- Williamson, D.L.; Bolster, D.R.; Kimball, S.R.; Jefferson, L.S. Time course changes in signaling pathways and protein synthesis in C2C12 myotubes following AMPK activation by AICAR. Am. J. Physiol. Endocrinol. Metab. 2006, 291, E80–E89. [Google Scholar] [CrossRef]
- Bolster, D.R.; Crozier, S.J.; Kimball, S.R.; Jefferson, L.S. AMP-activated protein kinase suppresses protein synthesis in rat skeletal muscle through down-regulated mammalian target of rapamycin (mTOR) signaling. J. Biol. Chem. 2002, 277, 23977–23980. [Google Scholar] [CrossRef] [Green Version]
- Sandrine, H.; Gareth, J.B.; Ulrike, K.; Jigna, V.P.; Didier, V.; Luc, B.; Alain, L.; Louis, H.; Christopher, G.P.; Mark, H.R. Activation of AMP-Activated Protein Kinase Leads to the Phosphorylation of Elongation Factor 2 and an Inhibition of Protein Synthesis. Curr. Biol. 2002, 12, 1419–1423. [Google Scholar] [CrossRef] [Green Version]
- Reiter, A.K.; Bolster, D.R.; Crozier, S.J.; Kimball, S.R.; Jefferson, L.S. Repression of protein synthesis and mTOR signaling in rat liver mediated by the AMPK activator aminoimidazole carboxamide ribonucleoside. Am. J. Physiol. Endocrinol. Metab. 2005, 288, 980–988. [Google Scholar] [CrossRef]
- McLeod, L.E.; Proud, C.G. ATP depletion increases phosphorylation of elongation factor eEF2 in adult cardiomyocytes independently of inhibition of mTOR signalling. Febs. Lett. 2002, 531, 448–452. [Google Scholar] [CrossRef] [Green Version]
- Chan, A.Y.; Soltys, C.L.; Young, M.E.; Proud, C.G.; Dyck, J.R. Activation of AMP-activated protein kinase inhibits protein synthesis associated with hypertrophy in the cardiac myocyte. J. Biol. Chem. 2004, 279, 32771–32779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiang, X.; Saha, A.K.; Wen, R.; Ruderman, N.B.; Luo, Z. AMP-activated protein kinase activators can inhibit the growth of prostate cancer cells by multiple mechanisms. Biochem. Biophys. Res. Commun. 2004, 321, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Fay, J.R.; Steele, V.; Crowell, J.A. Energy homeostasis and cancer prevention: The AMP-activated protein kinase. Cancer Prev. Res. 2009, 2, 301–309. [Google Scholar] [CrossRef] [Green Version]
- Figueiredo, V.C.; Markworth, J.F.; Cameron-Smith, D. Considerations on mTOR regulation at serine 2448: Implications for muscle metabolism studies. Cell Mol. Life Sci. 2017, 74, 2537–2545. [Google Scholar] [CrossRef]
- Bodine, S.C.; Latres, E.; Baumhueter, S.; Lai, V.K.; Nunez, L.; Clarke, B.A.; Poueymirou, W.T.; Panaro, F.J.; Na, E.; Dharmarajan, K.; et al. Identification of ubiquitin ligases required for skeletal muscle atrophy. Science 2001, 294, 1704–1708. [Google Scholar] [CrossRef]
- Morita, M.; Gravel, S.P.; Hulea, L.; Larsson, O.; Pollak, M.; St-Pierre, J.; Topisirovic, I. mTOR coordinates protein synthesis, mitochondrial activity and proliferation. Cell Cycle 2015, 14, 473–480. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, N.; Motohashi, N.; Uezumi, A.; Fukada, S.; Yoshimura, T.; Itoyama, Y.; Aoki, M.; Miyagoe-Suzuki, Y.; Takeda, S. NO production results in suspension-induced muscle atrophy through dislocation of neuronal NOS. J. Clin. Investig. 2007, 117, 2468–2476. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Su, S.; Zhang, Y.; Qian, J.; Liu, P. Control of mTOR signaling by ubiquitin. Oncogene 2019, 38, 3989–4001. [Google Scholar] [CrossRef]
- Aramburo, C.; Alba-Betancourt, C.; Luna, M.; Harvey, S. Expression and function of growth hormone in the nervous system: A brief review. Gen. Comp. Endocrinol. 2014, 203, 35–42. [Google Scholar] [CrossRef]
- Chen, L.; Chen, L.; Wan, L.; Huo, Y.; Huang, J.; Li, J.; Lu, J.; Xin, B.; Yang, Q.; Guo, C. Matrine improves skeletal muscle atrophy by inhibiting E3 ubiquitin ligases and activating the Akt/mTOR/FoxO3alpha signaling pathway in C2C12 myotubes and mice. Oncol. Rep. 2019, 42, 479–494. [Google Scholar] [CrossRef]
- Jorgensen, S.B.; Wojtaszewski, J.F.; Viollet, B.; Andreelli, F.; Birk, J.B.; Hellsten, Y.; Schjerling, P.; Vaulont, S.; Neufer, P.D.; Richter, E.A.; et al. Effects of alpha-AMPK knockout on exercise-induced gene activation in mouse skeletal muscle. Faseb. J. 2005, 19, 1146–1148. [Google Scholar] [CrossRef]
- Nystrom, G.J.; Lang, C.H. Sepsis and AMPK Activation by AICAR Differentially Regulate FoxO-1, -3 and -4 mRNA in Striated Muscle. Int. J. Clin. Exp. Med. 2008, 1, 50–63. [Google Scholar]
- Porter, R.K.; Brand, M.D. Causes of differences in respiration rate of hepatocytes from mammals of different body mass. Am. J. Physiol. 1995, 269, R1213–R1224. [Google Scholar] [CrossRef]
- Cooper, C.E.; Giulivi, C. Nitric oxide regulation of mitochondrial oxygen consumption, I.I: Molecular mechanism and tissue physiology. Am. J. Physiol. Cell Physiol. 2007, 292, C1993–C2003. [Google Scholar] [CrossRef] [Green Version]
- Kelly, D.P.; Scarpulla, R.C. Transcriptional regulatory circuits controlling mitochondrial biogenesis and function. Genes Dev. 2004, 18, 357–368. [Google Scholar] [CrossRef] [Green Version]
- Skotland, T.; Hessvik, N.P.; Sandvig, K.; Llorente, A. Exosomal lipid composition and the role of ether lipids and phosphoinositides in exosome biology. J. Lipid Res. 2019, 60, 9–18. [Google Scholar] [CrossRef] [Green Version]
- Porta, C.; Paglino, C.; Mosca, A. Targeting PI3K/Akt/mTOR Signaling in Cancer. Front. Oncol. 2014, 4, 64. [Google Scholar] [CrossRef] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pan, B.; Long, H.; Yuan, Y.; Zhang, H.; Peng, Y.; Zhou, D.; Liu, C.; Xiang, B.; Huang, Y.; Zhao, Y.; et al. Identification of Body Size Determination Related Candidate Genes in Domestic Pig Using Genome-Wide Selection Signal Analysis. Animals 2022, 12, 1839. https://doi.org/10.3390/ani12141839
Pan B, Long H, Yuan Y, Zhang H, Peng Y, Zhou D, Liu C, Xiang B, Huang Y, Zhao Y, et al. Identification of Body Size Determination Related Candidate Genes in Domestic Pig Using Genome-Wide Selection Signal Analysis. Animals. 2022; 12(14):1839. https://doi.org/10.3390/ani12141839
Chicago/Turabian StylePan, Bing, Haoyuan Long, Ying Yuan, Haoyuan Zhang, Yangyang Peng, Dongke Zhou, Chengli Liu, Baiju Xiang, Yongfu Huang, Yongju Zhao, and et al. 2022. "Identification of Body Size Determination Related Candidate Genes in Domestic Pig Using Genome-Wide Selection Signal Analysis" Animals 12, no. 14: 1839. https://doi.org/10.3390/ani12141839