
Effective Selection for Lower Mortality in Organic Pigs through Selection for Total Number Born and Number of Dead Piglets


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Simulated Traits
2.2. Data Collection
2.3. Parameter Estimation
2.4. Breeding Schemes
2.5. Breeding Goals
2.6. Simulated Breeding Schemes
2.7. Sensitivity Analysis
2.8. Data Analyses
3. Results
3.1. Simulations
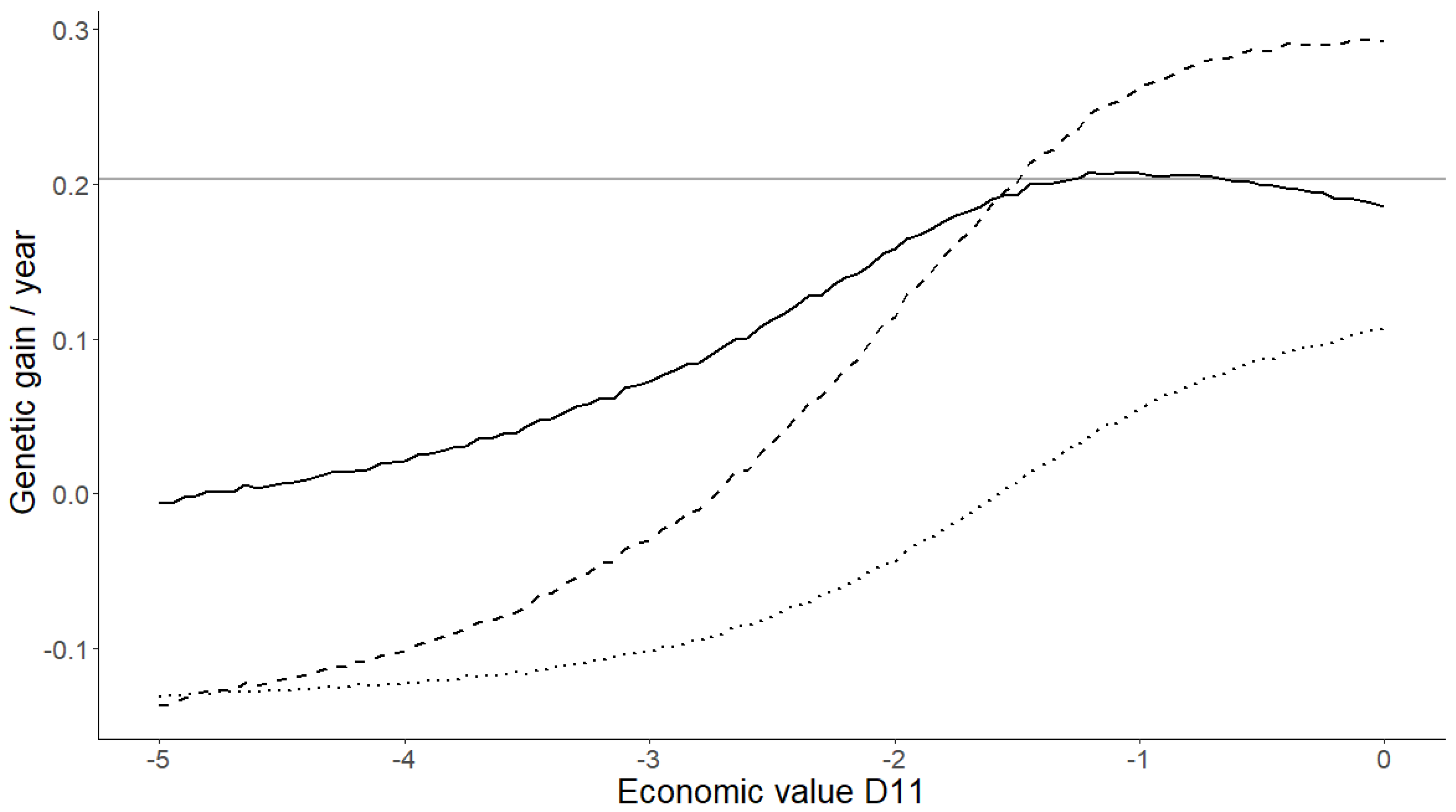
3.2. Sensitivity Analysis
4. Discussion
4.1. Opportunities of the Two-Trait Breeding Goal
4.2. Breeding for Lower Mortality in Organic Pig Production
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
| Breeding Goal | Economic Value | ΔF % per Generation | ||
|---|---|---|---|---|
| L11 1 | TNB 2 | D11 3 | ||
| Focus only on L11 | 1 | - | - | 0.89 |
| Focus only on TNB | - | 1 | 0 | 0.82 |
| Focus only on D11 | - | 0 | −1 | 0.85 |
| Equal focus on TNB and D11 | - | 1 | −1 | 0.86 |
| Emphasize D11 4 | - | 1 | −1.60 | 0.89 |
References
- Edwards, S.A.; Baxter, E.M. Piglet mortality: Causes and prevention. In The Gestating and Lactating Sow; Farmer, C., Ed.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2015; pp. 254–278. [Google Scholar]
- Leenhouwers, J.I.; Ten Napel, J.; Hanenberg, E.H.A.T.; Merks, J.W.M. Breeding replacement gilts for organic pig herds. Animal 2011, 5, 615–621. [Google Scholar] [CrossRef] [Green Version]
- Prunier, A.; Dipper, S.; Bochicchio, D.; Edwards, S.; Leeb, C.; Lindgren, K.; Sundrum, A.; Dietze, K.; Bonde, M. Characteristics of organic pig farms in selected European countries and their possible influence on litter size and piglet mortality. Org. Agric. 2014, 4, 163–173. [Google Scholar] [CrossRef]
- Rangstrup-Christensen, L.; Schild, S.-L.A.; Pedersen, L.J.; Sørensen, J.T. Causes of preweaning mortality in organic outdoor sow herds. Res. Vet. Sci. 2018, 118, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Grandinson, K.; Lund, M.S.; Rydhmer, L.; Strandberg, E. Genetic parameters for the piglet mortality traits crushing, stillbirth and total mortality, and their relation to birth weight. Acta Agric. Scand. Sect. A Anim. Sci. 2002, 52, 167–173. [Google Scholar] [CrossRef]
- IFOAM. Principles of Organic Agriculture 2005. Available online: https://www.ifoam.bio/en/organic-landmarks/principles-organic-agriculture (accessed on 25 November 2021).
- Gäde, S.; Bennewitz, J.; Kirchner, K.; Looft, H.; Knap, P.W.; Thaller, G.; Kalm, E. Genetic parameters for maternal behaviour traits in sows. Livest. Sci. 2008, 114, 31–41. [Google Scholar] [CrossRef]
- Damgaard, L.H.; Rydhmer, L.; Løvendahl, P.; Grandinson, K. Genetic parameters for within-litter variation in piglet birth weight and change in within-litter variation during suckling. J. Anim. Sci. 2003, 81, 604–610. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, B.; Su, G.; Lund, M.S.; Madsen, P. Selection for increased number of piglets at d5 after farrowing has increased litter size and reduced piglet mortality. J. Anim. Sci. 2013, 91, 2575–2582. [Google Scholar] [CrossRef] [PubMed]
- Baxter, E.M.; Edwards, S.A. Piglet mortality and morbidity: Inevitable or unacceptable. In Advances in Pig Welfare; Špinka, M., Ed.; Elsevier: Cambridge, MA, USA, 2018. [Google Scholar]
- Hansen, C.; SEGES. Landsgennemsnit for Produktivitet i Produktionen af Grise i 2020. 2021. Available online: https://svineproduktion.dk/Publikationer/Kilder/Notater/2021/2115 (accessed on 30 March 2022).
- Chu, T.T.; Zaalberg, R.M.; Bovbjerg, H.; Jensen, J.; Villumsen, T.M. Genetic variation in piglet mortality in outdoor organic production systems. Animal 2022, 16, 100529. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, H.M.; Olesen, I.; Navrud, S.; Kolstad, K.; Amer, P. How to consider the value of farm animals in breeding goals. A review of current status and future challenges. J. Agric. Environ. Ethics 2011, 24, 309–330. [Google Scholar] [CrossRef]
- Leite, N.G.; Knol, E.F.; Garcia, A.L.S.; Lopes, M.S.; Zak, L.; Tsuruta, S.; E Silva, F.F.; Lourenco, D. Investigating pig survival in different production phases using genomic models. J. Anim. Sci. 2021, 99, skab217. [Google Scholar] [CrossRef] [PubMed]
- Quiniou, N.; Dagorn, J.; Gaudré, D. Variation of piglets’ birth weight and consequences on subsequent performance. Livest. Prod. Sci. 2002, 78, 63–70. [Google Scholar] [CrossRef]
- Wientjes, J.G.M.; Soede, N.M.; Van der Peet-Schwering, C.M.C.; Van den Brand, H.; Kemo, B. Piglet uniformity and mortality in large organic litters: Effects of parity and pre-mating diet composition. Livest. Sci. 2012, 144, 218–229. [Google Scholar] [CrossRef]
- Vernersen, A.; Ask, B.; Sørensen, A.C.; Serup, T.; SEGES. Differentietet avl af grise—Via Danavl. 2015. Available online: https://www.landbrugsinfo.dk/-/media/landbrugsinfo/public/0/0/9/diff_avl_af_oeko_grise_via_danavl.pdf (accessed on 10 November 2021).
- Madsen, P.; Jensen, J. A User’s Guide to DMU; Version 6, Release 5.2. Aarhus University: Foulum, Denmark, 2013. [Google Scholar]
- Zaalberg, R.M.; Nielsen, H.M.; Sørensen, A.C.; Chu, T.T.; Jensen, J.; Villumsen, T.M. The effect of using organic or conventional sires on genetic gain in organic pigs: A simulation study. Animals 2022, 12, 455. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, L.D.; Sørensen, A.C.; Henryon, M.; Ansari-Mahyari, S.; Berg, P. ADAM: A computer program to simulate selective breeding schemes for animals. Livest. Sci. 2009, 121, 343–344. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.R-project.org/ (accessed on 12 June 2022).
- Hazel, L.N.; Dickerson, G.E.; Freeman, A.E. The selection index—Then, now, and for the future. J. Dairy Sci. 1994, 77, 3236–3251. [Google Scholar] [CrossRef]
- Cole, J.B.; VanRaden, P.M. Symposium review: Possibilities in an age of genomics: The future of selection indices. J. Dairy Sci. 2018, 101, 3686–3701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lund, V. Natural living—A precondition for animal welfare in organic farming. Livest. Sci. 2006, 100, 71–83. [Google Scholar] [CrossRef] [Green Version]
- Merks, J.W.M.; Mathur, P.K.; Knol, E.F. New phenotypes for new breeding goals in pigs. Animal 2012, 6, 535–543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallenbeck, A.; Rydhmer, L.; Lundeheim, N. GxE interactions for growth and carcass leanness: Re-ranking of boars in organic and conventional pig production. Livest. Sci. 2009, 123, 154–160. [Google Scholar] [CrossRef]
- Rangstrup-Christensen, L.; Krogh, M.A.; Pedersen, L.J.; Sørensen, J.T. Sow level risk factors for early piglet mortality and crushing in organic outdoor production. Animal 2018, 12, 810–818. [Google Scholar] [CrossRef] [PubMed]

| Trait | L11 | TNB | D11 | SDP | Mean |
|---|---|---|---|---|---|
| Alive at day 11 | 0.06 | 0.80 | −0.18 | 3.33 | 12.92 |
| Total number born | 0.87 | 0.09 | 0.45 | 3.67 | 14.61 |
| Dead at day 11 | 0.20 | 0.66 | 0.06 | 2.23 | 1.69 |
| Breeding Goal | Economic Value | Annual Genetic Gain 1 | ||||
|---|---|---|---|---|---|---|
| L11 | TNB | D11 | L11 | TNB | D11 | |
| Focus only on L11 | 1 | - | - | 0.201 | 0.227 | 0.026 |
| Focus only on TNB | - | 1 | 0 | 0.185 | 0.292 | 0.107 |
| Focus only on D11 | - | 0 | −1 | −0.082 | −0.223 | −0.141 |
| Equal focus on TNB and D11 | - | 1 | −1 | 0.207 | 0.262 | 0.055 |
| Emphasize D11 2 | - | 1 | −1.60 | 0.190 | 0.187 | −0.003 |
| Scenario | Annual Genetic Gain L11 | |||
|---|---|---|---|---|
| Genetic Correlation 1 | Heritability | Single-Trait BG | Two-Trait BG | p-Value 2 |
| original 3 | original 6 | 0.201 | 0.207 | >0.001 |
| strong 4 | Original | 0.133 | 0.158 | >0.001 |
| weak 5 | Original | 0.356 | 0.353 | 0.236 |
| original | equal 7 | 0.201 | 0.206 | 0.012 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zaalberg, R.M.; Villumsen, T.M.; Jensen, J.; Chu, T.T. Effective Selection for Lower Mortality in Organic Pigs through Selection for Total Number Born and Number of Dead Piglets. Animals 2022, 12, 1796. https://doi.org/10.3390/ani12141796
Zaalberg RM, Villumsen TM, Jensen J, Chu TT. Effective Selection for Lower Mortality in Organic Pigs through Selection for Total Number Born and Number of Dead Piglets. Animals. 2022; 12(14):1796. https://doi.org/10.3390/ani12141796
Chicago/Turabian StyleZaalberg, Roos M., Trine M. Villumsen, Just Jensen, and Thinh T. Chu. 2022. "Effective Selection for Lower Mortality in Organic Pigs through Selection for Total Number Born and Number of Dead Piglets" Animals 12, no. 14: 1796. https://doi.org/10.3390/ani12141796