
Effects of High Ambient Temperature on Small Intestinal Morphology and Colonic Microbiota in Weaned Piglets

 , , , , ,
, , , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Animals, Diets and Experimental Design
2.3. Samples Collection
2.4. Organ Index Calculation Formula
2.5. Intestinal Morphology and Structure
2.6. DNA Extraction and 16S rRNA Gene Sequencing
2.7. Bioinformatics Analysis for Colonic Microbiota
2.8. Statistical Analysis
3. Results
3.1. Growth Performance
3.2. Organs Index
3.3. Intestinal Morphology
3.4. Alpha Diversity
3.5. Beta Diversity
3.6. Colonic Bacterial Community
3.7. Predicted Functional Profiles of Microbiota Communities Using Tax4Fun
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- St-Pierre, N.R.; Cobanov, B.; Schnitkey, G. Economic losses from heat stress by US livestock industries. J. Dairy Sci. 2003, 86, E52–E77. [Google Scholar] [CrossRef] [Green Version]
- Fl, A.; Wz, B.; Hhl, B.; Jjc, B.; Mpg, C.; Bjl, B.; Frdb, D.; Awb, E. What have we learned about the effects of heat stress on the pig industry? Animal 2021, 16, 100349. [Google Scholar]
- Gardiner, G.E.; Metzler-Zebeli, B.U.; Lawlor, P.G. Impact of intestinal microbiota on growth and feed efficiency in pigs: A review. Microorganisms 2020, 8, 1886. [Google Scholar] [CrossRef] [PubMed]
- Yadav, S.; Jha, R. Strategies to modulate the intestinal microbiota and their effects on nutrient utilization, performance, and health of poultry. J. Anim. Sci. Biotechnol. 2019, 10, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Tarini, J.; Wolever, T.M.S. The fermentable fibre inulin increases postprandial serum short-chain fatty acids and reduces free-fatty acids and ghrelin in healthy subjects. Appl. Physiol. Nutr. Metab. 2010, 35, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Bhattarai, Y.; Muniz Pedrogo, D.A.; Kashyap, P.C. Irritable bowel syndrome: A gut microbiota-related disorder? Am. J. Physiol.-Gastrointest. Liver Physiol. 2017, 312, G52–G62. [Google Scholar] [CrossRef]
- Plaza-Díaz, J.; Solís-Urra, P.; Rodríguez-Rodríguez, F.; Olivares-Arancibia, J.; Navarro-Oliveros, N.; Abadía-Molina, F.; Álvarez-Mercado, A.I. The gut barrier, intestinal microbiota, and liver disease: Molecular mechanisms and strategies to manage. Int. J. Mol. Sci. 2020, 21, 8351. [Google Scholar] [CrossRef]
- Ferreira, R.S.; Mendonca, L.A.B.M.; Ribeiro, C.F.A.; Calças, N.C.; Guimarães, R.C.A.; Nascimento, V.A.; Gielow, K.C.F.; Carvalho, C.M.E.; Castroa, A.P.; Franco, O.L. Relationship between intestinal microbiota, diet and biological systems: An integrated view. Crit. Rev. Food Sci. Nutr. 2022, 62, 1166–1186. [Google Scholar] [CrossRef]
- Jiang, H.; Chen, W.; Su, L.; Huang, M.; Lin, L.; Su, Q.; Li, G.; Ahmad, H.I.; Li, L.; Zhang, X.; et al. Impact of host intraspecies genetic variation, diet, and age on bacterial and fungal intestinal microbiota in tigers. Microbiologyopen 2020, 9, e1050. [Google Scholar] [CrossRef]
- Oliveira, R.F.; Moreira, R.H.R.; Ribeiro, B.; Resende, M.; Chaves, R.F.; Gionbelli, M.P.; Ferreira, R.A. Acute heat stress compromises the physiology of growing pigs. Arch. De Zootec. 2019, 68, 300–302. [Google Scholar] [CrossRef] [Green Version]
- Renaudeau, D.; Francès, G.; Dubois, S.; Gilbert, H.; Noblet, J. Effect of thermal heat stress on energy utilization in two lines of pigs divergently selected for residual feed intake. J. Anim. Sci. 2013, 91, 1162–1175. [Google Scholar] [CrossRef] [PubMed]
- Burkholder, K.M.; Thompson, K.L.; Einstein, M.E.; Applegate, T.J.; Patterson, J.A. Influence of stressors on normal intestinal microbiota, intestinal morphology, and susceptibility to Salmonella enteritidis colonization in broilers. Poult. Sci. 2008, 87, 1734–1741. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Xiao, K.; Ke, Y.L.; Jiao, L.F.; Hu, C.H.; Diao, Q.Y.; Shi, B.; Zou, X.T. Effect of a probiotic mixture on intestinal microflora, morphology, and barrier integrity of broilers subjected to heat stress. Poult. Sci. 2014, 93, 581–588. [Google Scholar] [CrossRef] [PubMed]
- Mathilde, L.S.; Olivier, Z.; Isabelle, H.; Juliette, R.; Hélène, G.; Mario, G.; Yvon, B.; Jean-Luc, G.; David, R. Effect of chronic and acute heat challenges on fecal microbiota composition, production, and thermoregulation traits in growing pigs. J. Anim. Sci. 2019, 97, 3845–3858. [Google Scholar]
- Yu, J.; Song, Y.; Yu, B.; He, J.; Chen, D.W. Tannic acid prevents post-weaning diarrhea by improving intestinal barrier integrity and function in weaned piglets. J. Anim. Sci. Biotechnol. 2020, 11, 1–11. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. S1), 4516–4522. [Google Scholar] [CrossRef] [Green Version]
- Youssef, N.; Sheik, C.S.; Krumholz, L.R.; Najar, F.Z.; Roe, B.A.; Elshahed, M.S. Comparison of species richness estimates obtained using nearly complete fragments and simulated pyrosequencing-generated fragments in 16S rRNA gene-based environmental surveys. Appl. Environ. Microbiol. 2009, 75, 5227–5236. [Google Scholar] [CrossRef] [Green Version]
- Hess, M.; Sczyrba, A.; Egan, R.; Kim, T.W.; Chokhawala, H.; Schroth, G.; Luo, S.J.; Clark, D.S.; Chen, F.; Zhang, T.; et al. Metagenomic discovery of biomass-degrading genes and genomes from cow rumen. Science 2011, 331, 463–467. [Google Scholar] [CrossRef] [Green Version]
- Lundberg, D.S.; Yourstone, S.; Mieczkowski, P.; Jones, C.D.; Dangl, J.L. Practical innovations for high-throughput amplicon sequencing. Nat. Methods 2013, 10, 999–1002. [Google Scholar] [CrossRef]
- Li, B.; Zhang, X.; Guo, F.; Wu, W.; Zhang, T. Characterization of tetracycline resistant bacterial community in saline activated sludge using batch stress incubation with high-throughput sequencing analysis. Water Res. 2013, 47, 4207–4216. [Google Scholar] [CrossRef]
- Abdi, H.; Williams, L.J. Principal component analysis. Wiley Interdiscip. Rev. Comput. Stat. 2010, 2, 433–459. [Google Scholar] [CrossRef]
- Chapman, M.G.; Underwood, A.J. Ecological patterns in multivariate assemblages: Information and interpretation of negative values in ANOSIM tests. Mar. Ecol. Prog. Ser. 1999, 180, 257–265. [Google Scholar] [CrossRef]
- Qin, J.; Li, Y.; Cai, Z.; Li, S.H.; Zhu, J.F.; Zhang, F.; Liang, S.S.; Zhang, W.W.; Guan, Y.L.; Shen, D.Q.; et al. A metagenome-wide association study of gut microbiota in type 2 diabetes. Nature 2012, 490, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Baumgard, L.H.; Rhoads, R.P., Jr. Effects of heat stress on postabsorptive metabolism and energetics. Annu. Rev. Anim. Biosci. 2013, 1, 311–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayorga, E.J.; Kvidera, S.K.; Horst, E.A.; Al-Qaisi, M.; McCarthy, C.S.; Abeyta, M.A.; Lei, S.; Elsasser, T.H.; Kahl, S.; Kiros, T.G.; et al. Effects of dietary live yeast supplementation on growth performance and biomarkers of metabolism and inflammation in heat-stressed and nutrient-restricted pigs. Transl. Anim. Sci. 2021, 5, txab072. [Google Scholar] [CrossRef]
- Oliveira, A.C.F.; Vanelli, K.; Sotomaior, C.S.; Weber, S.H.; Costa, L.B. Impacts on performance of growing-finishing pigs under heat stress conditions: A meta-analysis. Vet. Res. Commun. 2019, 43, 37–43. [Google Scholar] [CrossRef]
- Fu, C.; Zhang, Y.; Jing, Q.; Shi, T.; Wei, X.; Liu, X. Effect of Chinese herbal medicine on growth performance, immune organ index and antioxidant functions in broiler chickens. Int. J. Agric. Biol. 2018, 20, 1677–1681. [Google Scholar]
- Jiang, M.H.; Zhu, L.; Jiang, J.G. Immunoregulatory actions of polysaccharides from Chinese herbal medicine. Expert Opin. Ther. Targets 2010, 14, 1367–1402. [Google Scholar] [CrossRef]
- Collin, A.; Lebreton, Y.; Fillaut, M.; Vincent, A.; Thomas, F.; Herpin, P. Effects of exposure to high temperature and feeding level on regional blood flow and oxidative capacity of tissues in piglets. Exp. Physiol. 2001, 86, 83–91. [Google Scholar] [CrossRef] [Green Version]
- Kwon, O.; Han, T.S.; Son, M.Y. Intestinal morphogenesis in development, regeneration, and disease: The potential utility of intestinal organoids for studying compartmentalization of the crypt-villus structure. Front. Cell Dev. Biol. 2020, 8, 593969. [Google Scholar] [CrossRef]
- Morales, A.; Hernández, L.; Buenabad, L.; Avelar, E.; Bernal, H.; Baumgard, L.H.; Cervantes, M. Effect of heat stress on the endogenous intestinal loss of amino acids in growing pigs. J. Anim. Sci. 2016, 94, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Ortega, A.D.S.V.; Szabó, C. Adverse effects of heat stress on the intestinal integrity and function of pigs and the mitigation capacity of dietary antioxidants: A review. Animals 2021, 11, 1135. [Google Scholar] [CrossRef]
- Yu, J.; Yin, P.; Liu, F.; Cheng, G.; Guo, K.J.; Lu, A.; Zhu, X.Y.; Luan, W.L.; Xu, J.Q. Effect of heat stress on the porcine small intestine: A morphological and gene expression study. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2010, 156, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Yin, J.; Du, M.; Yan, P.; Xu, J.; Zhu, X.; Yu, J. Heat-stress-induced damage to porcine small intestinal epithelium associated with downregulation of epithelial growth factor signaling. J. Anim. Sci. 2009, 87, 1941–1949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heller, F.; Duchmann, R. Intestinal flora and mucosal immune responses. Int. J. Med. Microbiol. 2003, 293, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Flint, H.J.; Scott, K.P.; Louis, P.; Duncan, S.H. The role of the gut microbiota in nutrition and health. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 577–589. [Google Scholar] [CrossRef]
- Vrieze, A.; Holleman, F.; Zoetendal, E.G.; Vos, W.M.; Hoekstra, J.B.L.; Nieuwdorp, M. The environment within: How gut microbiota may influence metabolism and body composition. Diabetologia 2010, 53, 606–613. [Google Scholar] [CrossRef] [Green Version]
- Rojo, D.; Méndez-García, C.; Raczkowska, B.A.; Bargiela, R.; Moya, A.; Ferrer, M.; Barbas, C. Exploring the human microbiome from multiple perspectives: Factors altering its composition and function. FEMS Microbiol. Rev. 2017, 41, 453–478. [Google Scholar] [CrossRef] [Green Version]
- Rowan, F.; Docherty, N.G.; Murphy, M.; Murphy, B.; Coffey, J.C.; O’Connell, P.R. Desulfovibrio bacterial species are increased in ulcerative colitis. Dis. Colon Rectum 2010, 53, 1530–1536. [Google Scholar] [CrossRef]
- Lam, Y.Y.; Ha CW, Y.; Campbell, C.R.; Mitchell, A.J.; Dinudom, A.; Oscarsson, J.; Cook, D.I.; Hunt, N.H.; Caterson, D.; Holmes, A.J.; et al. Increased gut permeability and microbiota change associate with mesenteric fat inflammation and metabolic dysfunction in diet-induced obese mice. PLoS ONE 2012, 7, e34233. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.S.; Li, W.N.; Wang, H.Y.; Ma, Y.M.; Zhao, X.Y.; Yang, H.; Qian, J.M.; Li, J.N. VSL# 3 can prevent ulcerative colitis-associated carcinogenesis in mice. World J. Gastroenterol. 2018, 24, 4254. [Google Scholar] [PubMed]
- Kanno, M.; Katayama, T.; Morita, N.; Tamaki1, H.; Hanada1, S.; Kamagata1, Y. Catenisphaera adipataccumulans gen. nov., sp. nov., a member of the family Erysipelotrichaceae isolated from an anaerobic digester. Int. J. Syst. Evol. Microbiol. 2015, 65 Pt 3, 805–810. [Google Scholar] [CrossRef] [PubMed]
- Cox, L.M.; Sohn, J.; Tyrrell, K.L.; Citron, D.M.; Lawson, P.A.; Patel, N.B.; Iizumi, T.; Perez-Perez, G.I.; Goldstein, E.J.C.; Blaser, M.J. Description of two novel members of the family Erysipelotrichaceae: Ileibacterium valens gen. nov., sp. nov. and Dubosiella newyorkensis, gen. nov., sp. nov., from the murine intestine, and emendation to the description of Faecalibacterium rodentium. Int. J. Syst. Evol. Microbiol. 2017, 67, 1247. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | % | Nutrients Levels 3 | % |
|---|---|---|---|
| Corn | 25.00 | DE, Mcal/kg | 14.82 |
| Extruded corn | 30.50 | CP | 18.35 |
| Peeling soybean meal | 10.00 | Ca | 0.62 |
| Extruded soybean | 7.00 | TP | 0.55 |
| Wheat bran | 1.45 | AP | 0.36 |
| Rice, broken | 7.90 | D-Lys | 1.35 |
| Soy protein concentrate | 3.50 | D-Met | 0.42 |
| Plasma protein powder | 2.00 | D-Trp | 0.24 |
| Fish meal | 3.50 | D-Thr | 0.79 |
| Whey powder | 3.00 | ||
| Soybean oil | 2.00 | ||
| Limestone | 0.57 | ||
| CaHPO4 | 0.38 | ||
| Salt | 0.30 | ||
| L-Lys•HCl | 0.39 | ||
| DL-Met | 0.12 | ||
| L-Thr | 0.04 | ||
| Chloride choline | 0.10 | ||
| Vitamin premix 1 | 0.05 | ||
| Mineral premix 2 | 0.20 | ||
| Sucrose | 2.00 | ||
| Total | 100.00 |
| Items | CON Group | HT Group | p-Value |
|---|---|---|---|
| Initial weight, kg | 8.61 ± 0.24 | 8.61 ± 0.24 | 0.971 |
| Final weight, kg | 17.20 ± 0.72 a | 12.86 ± 0.31 b | 0.001 |
| ADFI, g/d | 559.20 ± 37.42 a | 318.03 ± 17.05 b | 0.001 |
| ADG, g/d | 408.93 ± 25.51 a | 202.98 ± 13.00 b | 0.001 |
| F/G | 1.37 ± 0.02 a | 1.59 ± 0.08 b | 0.022 |
| Organ Index | CON Group | HT Group | p-Value |
|---|---|---|---|
| Heart | 5.05 ± 0.17 | 4.88 ± 0.14 | 0.467 |
| Liver | 27.28 ± 0.67 a | 23.41 ± 1.02 b | 0.007 |
| Spleen | 2.31 ± 0.23 a | 1.72 ± 0.08 b | 0.030 |
| Lung | 9.95 ± 0.55 | 8.87 ± 0.36 | 0.128 |
| Kidney | 5.58 ± 0.20 | 5.12 ± 0.25 | 0.167 |
| Thymus | 1.43 ± 0.10 a | 0.93 ± 0.08 b | 0.001 |
| Items | CON Group | HT Group | p-Value |
|---|---|---|---|
| Duodenum | |||
| Villous height, μm | 511.94 ± 28.20 a | 421.02 ± 21.54 b | 0.023 |
| Crypt depth, μm | 162.44 ± 11.50 a | 216.64 ± 22.18 b | 0.048 |
| VH/CD | 3.25 ± 0.27 a | 2.09 ± 0.23 b | 0.005 |
| Jejunum | |||
| Villous height, μm | 366.19 ± 19.22 a | 281.48 ± 19.11 b | 0.007 |
| Crypt depth, μm | 128.49 ± 15.65 | 137.20 ± 13.54 | 0.681 |
| VH/CD | 3.27 ± 0.52 | 2.18 ± 0.25 | 0.080 |
| Ileum | |||
| Villous height, μm | 389.26 ± 26.44 a | 291.34 ± 21.91 b | 0.013 |
| Crypt depth, μm | 134.11 ± 12.05 | 144.03 ± 12.51 | 0.577 |
| VH/CD | 3.13 ± 0.42 a | 2.08 ± 0.17 b | 0.037 |
| Items | CON Group | HT Group | t-Test | Rank Sum Test |
|---|---|---|---|---|
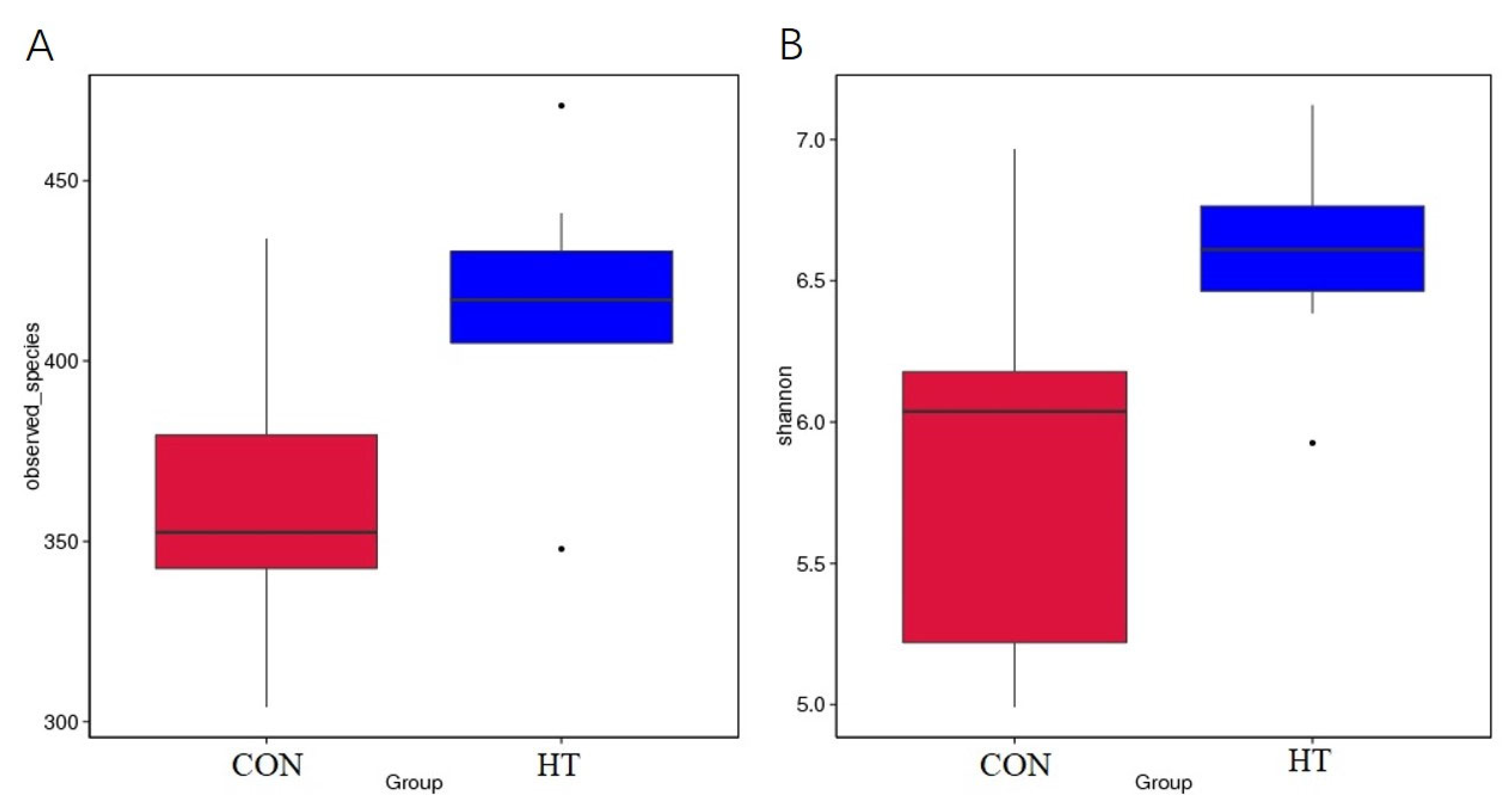
| Observed-species index | 362 | 416 | 0.016 | 0.045 |
| Shannon index | 5.855 | 6.595 | 0.019 | 0.028 |
| Groups | R-Value | p-Value |
|---|---|---|
| C-T | 0.3259 | 0.015 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xing, S.; Chen, S.; Zhao, Y.; Luo, Y.; Yu, B.; He, J.; Huang, Z.; Zheng, P.; Mao, X.; Luo, J.; et al. Effects of High Ambient Temperature on Small Intestinal Morphology and Colonic Microbiota in Weaned Piglets. Animals 2022, 12, 1743. https://doi.org/10.3390/ani12141743
Xing S, Chen S, Zhao Y, Luo Y, Yu B, He J, Huang Z, Zheng P, Mao X, Luo J, et al. Effects of High Ambient Temperature on Small Intestinal Morphology and Colonic Microbiota in Weaned Piglets. Animals. 2022; 12(14):1743. https://doi.org/10.3390/ani12141743
Chicago/Turabian StyleXing, Shuaibing, Shuai Chen, Ying Zhao, Yuheng Luo, Bing Yu, Jun He, Zhiqing Huang, Ping Zheng, Xiangbing Mao, Junqiu Luo, and et al. 2022. "Effects of High Ambient Temperature on Small Intestinal Morphology and Colonic Microbiota in Weaned Piglets" Animals 12, no. 14: 1743. https://doi.org/10.3390/ani12141743