1. Introduction
The cost of fish meal has increased in recent years. Therefore, the need for alternative protein sources has increased, prompting research into non-conventional feed ingredients. So far, insect protein has shown great promise as an alternative protein source [
1,
2]. Insects require less space for breeding, have short reproduction cycles, consume a wide range of food, and exhibit high conversion rates. Further, the negative environmental impact of insect breeding is much smaller than that of livestock breeding [
3]. Moreover, insect protein has a high nutritional value [
4]. Therefore, insect protein has gradually become a research hotspot for protein alternatives in animal feed [
5]. A variety of insects have been used to derive feed ingredients, with
Hermetia illucens [
6],
Tenebrio molitor [
7,
8], and
Musca domestica [
9] showing the highest potential.
Fly maggot protein comes from maggots cultured in a medium composed of microorganisms, milk powder, bean products, brown sugar, and starch. Fly maggot protein feed is affordable and costs less than fish meal, which is becoming increasingly more expensive. It contains a variety of amino acids, including essential amino acids required for animal growth [
10]. Similar to fish meal, fly maggot protein is a high-quality animal protein source [
11]. Compared to the broiler (
Gallus gallus) fed the basal diet, feeding diets containing 10–15% fly maggots can improve the carcass quality and growth performance of broiler chickens [
12]. Song et al. found that a feeding diet with 4.7–6.4% of fly maggot powder significantly improved the growth performance, immune indexes, and antioxidant enzyme activity in broiler chickens [
13]. In Wang’s study, a dietary maggot meal was also proved to be positively influential in flesh quality, and replacing 270 g∙kg
−1 fish meal did not have an impact on the growth performance and ingredient utilization of tilapia (
Oreochromis niloticus) [
14].
The scale of blue fox (
Alopex lagopus) breeding is increasing steadily. Blue foxes are carnivorous canines; hence, in captivity, most of the animal protein in their diet comes from fish meal. Accordingly, there is a high demand for fish meal in the blue fox breeding industry [
15,
16]. However, there are little data on the use of insect protein sources as a feed alternative for the blue fox diet. Studies examining the effects of insect protein on growth and plasma parameters have only been conducted on dogs [
17]. Furthermore, information on the microbiome of this species is very limited, although studies indicate that insect meal can positively influence the microbiota composition and improve the gastrointestinal health in these animals [
18,
19].
Accordingly, this study aimed to evaluate the effect of replacing fish meal with fly maggot protein feed on growth, the blood immune indexes, and the intestinal microbiota in weaned blue foxes. The results of the present study could contribute to explain the value of fly maggot protein as a substitute for fish meal in weaned blue fox diets.
4. Discussion
Fly maggots contain 59–65% crude protein [
21] and are a high-quality source of animal protein, comparable to fish meal [
22]. Fly maggot protein is known to improve growth in weaned piglets (
Sus scrofa), reducing the cost of growth and making pig breeding more economic [
23]. A study showed that when Chinese soft-shelled turtles (
Pelodiscus sinensis) were fed an appropriate amount of fly maggot protein, their feed coefficient effectively reduced, while their growth, immune indexes, and survival rates improved [
24,
25,
26]. Replacing fish meal with fly maggot meal not only improves growth and survival rates in tilapia but also increases their immunity and crude protein content [
5,
27]. These studies together indicate that the use of fly maggot protein as a feed supplement or animal protein alternative has no negative effect on growth and improves production efficiency.
In this study, the replacement of fish meal with fly maggot protein had a significant effect on the average daily weight gain of weaned foxes at various time points. However, when the entire experimental period was considered, no significant difference in this value was observed. The body weight of group M was higher than that of group F at 7 d and 14 d but lower at 21 d and 28 d, although the differences were not significant. Therefore, when weaned blue foxes grow to a certain stage, replacing fish meal with fly maggot protein may provide a better feeding effect and improve their growth. The experimental results suggest that fly maggot protein can replace fish meal for blue foxes at the weaning stage and reduce the cost of breeding.
In addition, although the incidence of diarrhea in group M was significantly higher than that in group F, there was no significant difference in the diarrhea index. The reasons for the high incidence of diarrhea may be as follows: (1) Monogastric animals may find it difficult to digest the chitin in insect proteins. Chitin has an anti-nutritional effect and can reduce protein utilization to a certain extent [
28,
29]. (2) The ratio of fish meal replacement is not optimal, and excessive addition of insect protein may have negative effects on animals [
30]. (3) The experiments were conducted on weaned foxes, who may have digestive organs that are not completely developed. Although the change in feed ingredients does not affect palatability, it may affect digestion in young blue foxes. (4) The stress of weaning and separation from companions may lead to disordered bodily functions in young blue foxes. However, our findings indicated that fly maggot protein can reduce the incidence of diarrhea in blue foxes at the middle and late stages. Therefore, further research is needed to determine the proportion of fish meal replacement with insect protein and the optimal feeding time to maximize the benefits of protein replacement.
IgG, IgA, and IgM are the main effector molecules mediating humoral immunity and reflect the immune status of the body [
31]. IgG has antiviral and antitoxin activities in vivo and is the most important immunoglobulin in humoral immunity [
32]. IgM is involved in intestinal immunity. The IgM levels in group M were always higher than those in group F, even at the early stages of the experiment. This may be because the fly maggot protein improved the immune status of young blue foxes during the pre-feeding period. Therefore, the IgM content on d 0 was already higher than that in group F, and this difference was maintained during the subsequent test period. IgA represents an important line of defense against the adhesion and colonization of pathogens in the intestinal mucosa, and it is also the most common and most secreted immunoglobulin in the body [
33]. In this study, there was no significant difference in IgA levels between group M and group F. This indicated that the replacement of fish meal with fly maggot protein did not create any immune stress in blue foxes during the weaning period.
Weaned blue foxes are prone to immune stress for the following reasons: discontinuation of breast milk, immature immune system, presence of protein antigens in the feed, and environmental changes. During conditions of immune stress, the levels of IL-1β, IL-6, and TNF-α increase. Instead of being used to maintain growth, the nutrients in the body are used to mount an immune response, inhibiting growth and metabolism. IL-1 is an important cytokine secreted by macrophages. Its excessive secretion can stimulate a variety of immune cells and inflammatory cells, thereby leading to the productions of cytokines, such as IL-6 and TNF-α, that induce inflammatory responses and cell damage. IL-2, an important factor in the series of lymphokines released during an immune response, is considered a marker of T cell activation. Our study found that the level of IL-2 was significantly higher in the fly maggot protein group than in the control group, while no significant difference in TNF-α was observed.
Therefore, the results of this study indicate that feed protein may cause stress responses in young foxes. The effects of fly protein feed were clearly more positive than those of fish meal. This is consistent with findings from studies on weaned piglets, loaches, and poultry [
23,
29,
34]. Fly maggot protein contains bioactive substances, such as fly maggot chitosan, fly maggot peptide, and lectin, which may improve immune function in young blue foxes.
Redox reactions are normal physiological and biochemical reactions and can lead to the generation of free radicals. MDA is an important product of lipid peroxidation and indicates the rate and intensity of lipid peroxidation. Thus, it can be used to indirectly estimate oxidative damage [
35]. T-AOC reflects the overall activity of enzymatic and non-enzymatic antioxidant systems [
36]. It is usually used to directly examine the activity of antioxidant enzymes in the body. SOD and CAT are the most important antioxidant enzymes and serve as a defense system against O
2− radicals. SOD can prevent excessive O
2− invasion and its activity is an indirect indicator of oxidative damage.
Antioxidant systems of organisms are affected differently by feeding insect proteins. Studies have shown that 4.7–6.4% of fly maggot protein in feed can positively influence immunity and antioxidant capacity in broiler chickens [
11]. However, other studies have found different results. The CAT levels in Nile tilapia were not significantly affected after feeding with fly maggot protein [
37]. Hence, the response to different protein sources can vary from species to species, and the defense strategies may also vary. In this experiment, the levels of SOD and T-AOC in group M had no significant difference in the later stage of the experiment. MDA levels did not show significant between-group differences throughout the experimental period. These findings suggest that feeding fly maggot protein to weaned blue foxes has no negative effect on their antioxidant capacity.
Microbes play an important role in the development of the intestinal mucosal system and tissues [
38]. Microbial colonization is a complex process. Each host develops a unique bacterial community structure through long-term evolution, which is maintained in a state of equilibrium via interaction with environmental factors. Moreover, there are significant differences in the microbiota between species. Various factors, such as maternal microflora composition, age, dietary changes, and antibiotics, all influence gut microflora composition [
39]. In this experiment, the Shannon index of group M was significantly lower than that of group F on day 0. However, no significant difference in ACE, Chao1, Simpson, and Shannon index values of the fecal microflora was observed between the two groups at 28 d. Hence, after feeding with fly maggot protein, the richness of the microbial community in weaned foxes could be increased, although the diversity remains unaffected.
The genus
Prevotella contains highly active bacteria that decompose hemicellulose [
40]. As part of the intestinal microflora, these bacteria play an important role in the degradation of hemicellulose and promote the degradation of non-structural carbohydrates [
41,
42], improving digestion and absorption. Hence, they have a positive impact on health.
Prevotella spp. play a key role in the utilization of starch, xylan, and pectin during digestion in cattle and sheep [
43,
44,
45]. In addition,
Prevotella can degrade plant cell walls [
42]. The abundance of
Prevotella in the rumen is associated with fiber digestibility [
46], and these bacteria also secrete proteases [
47]. Although
Prevotella is abundant in blue fox feces, the exact role of these bacteria in blue foxes remains to be determined. Blue foxes show a high utilization rate of protein- and fat-rich animal feed [
48]. However, whether and how their intestinal bacteria participate in the metabolism of nutrients, such as protein and fat, is still unclear.
Lactic acid bacteria produce a large amount of lactic acid, which can reduce the pH of the intestinal tract. These bacteria can also produce bacteriostatic metabolites and thereby, inhibit the reproduction and invasion of pathogenic bacteria. Therefore, lactic acid bacteria have a positive impact on immunity and intestinal health [
49].
Weissella spp. are present in most fermentation products [
50,
51]. In the early stage of fermentation,
Weisseria can even dominate in numbers. At this stage, many species of
Weisseria have been found to have the advantages of probiotics, with various probiotic properties, including: the production of bacteriocins, which have been found to inhibit pathogenic bacteria [
52]; prevent it from adhering to cells [
53]; produce exopolysaccharides, such as glucan, etc. [
54,
55,
56]; generate digestive enzymes [
57]; and have antioxidant capacity [
52,
58,
59,
60].
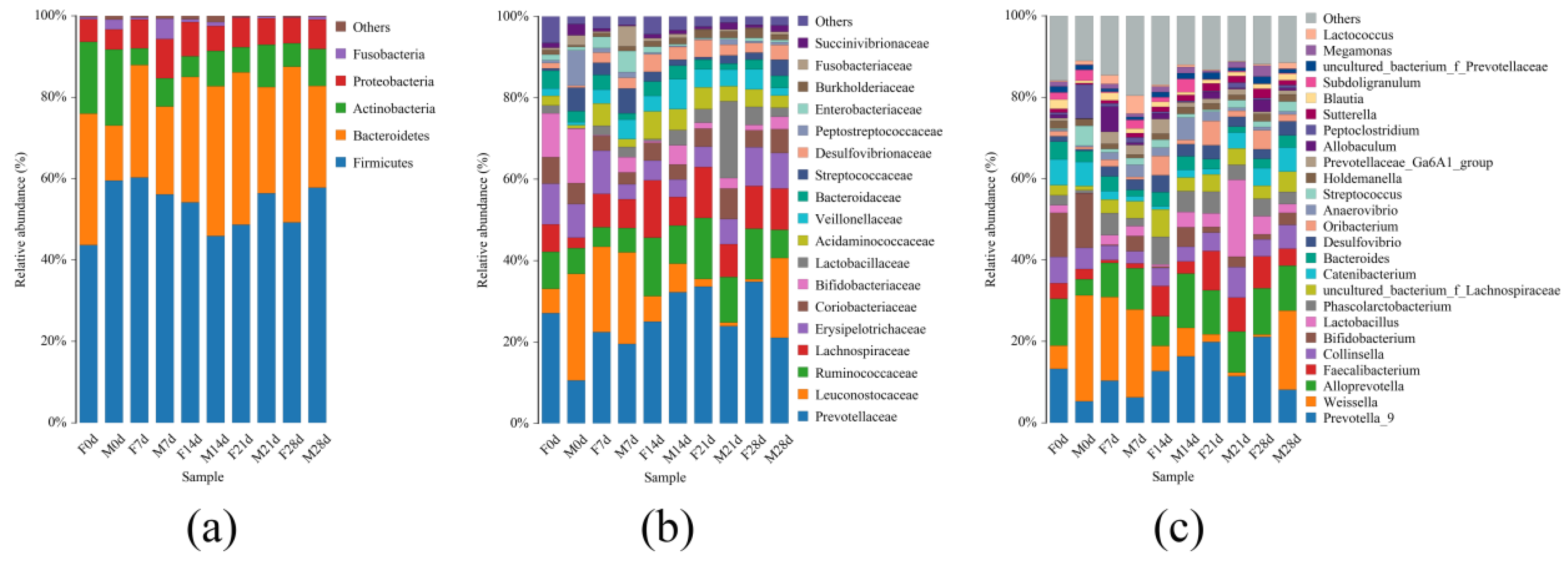
Consistent with Chen’s findings, this study showed that the fecal flora in weaned foxes mainly consists of
Firmicutes,
Bacteroidetes,
Actinobacteria,
Proteobacteria, and
Fusobacteria. Together, these bacteria account for 97% of the microbiota [
61]. At the family level,
Prevotellaceae was the dominant bacterial group. No significant difference in the relative abundance of
Prevotellaceae was observed between groups M and F overall, although this value was higher in group F on day 0. At the genus level,
Weissella and
Prevotella_9 made up the dominant flora. The presence of fly maggot protein in feed increased the relative abundance of
Weissella in weaned blue foxes, although the difference was not significant. In contrast, blue foxes consuming fish meal had a significantly higher relative abundance of
Prevotella_9. The effect of fly maggot protein on the relative abundance of other fecal microbes was not significant. However, the abundance of each group showed different trends of change over time. This suggests that when fly maggot protein was used to replace fish meal, there was no adverse effect on the structure of the fecal flora. Hence, fly maggot protein could replace fish meal in the diet of blue foxes during the weaning period.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}