
Effect of Dietary Sugarcane Bagasse Supplementation on Growth Performance, Immune Response, and Immune and Antioxidant-Related Gene Expressions of Nile Tilapia (Oreochromis niloticus) Cultured under Biofloc System

 ,
,  ,
,  ,
, 

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sugarcane Bagasse Powder Preparing
2.2. Diets Description
2.3. Experimental Design
2.4. Biofloc Water Preparation
2.5. Samples Preparation
2.6. Immunological Parameters and Growth Performance
2.7. Immune and Antioxidant-Related Genes Expression in Liver and Intestine
2.7.1. Tissue Sampling
2.7.2. RNA Extraction and cDNA Synthesis
2.7.3. Quantitative PCR
2.8. Statistical Analysis
3. Results
3.1. Growth Performance
3.2. Skin Mucus Immunity
3.3. Serum Immunity
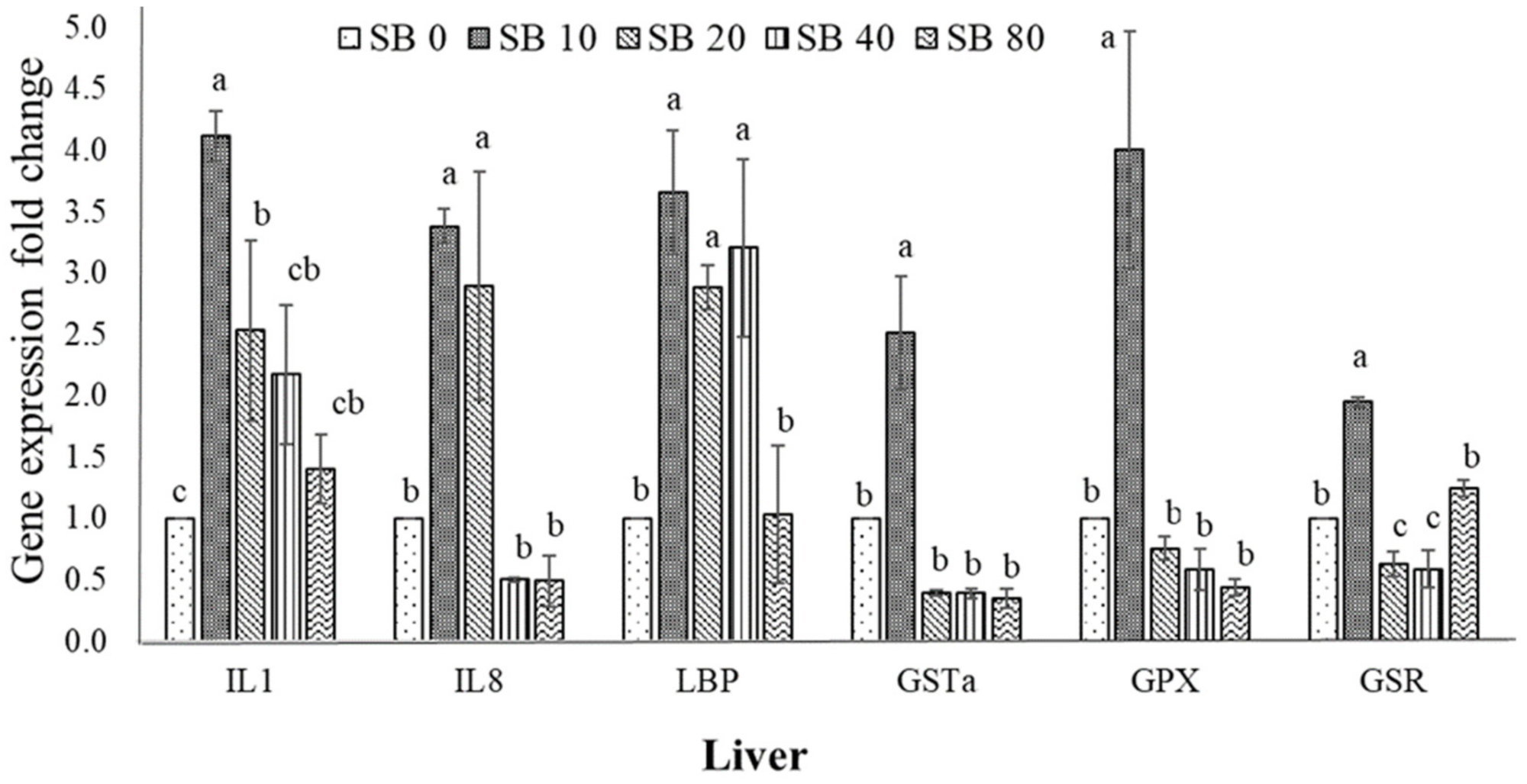
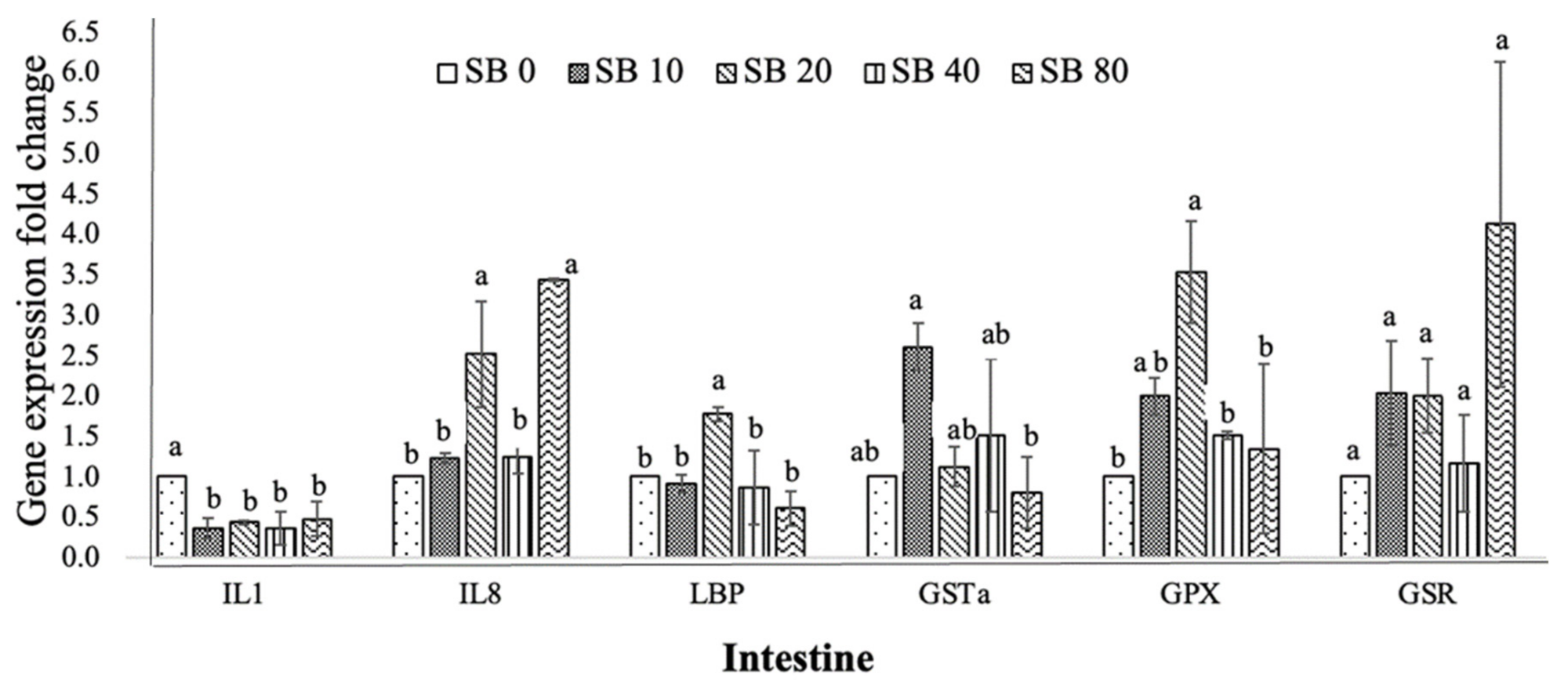
3.4. Expression of Immune-Related and Antioxidant Genes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture (SOFIA); Food and Agriculture Organization, Ed.; FAO: Rome, Italy, 2020. [Google Scholar]
- Sampantamit, T.; Ho, L.; Lachat, C.; Sutummawong, N.; Sorgeloos, P.; Goethals, P. Aquaculture Production and Its Environmental Sustainability in Thailand: Challenges and Potential Solutions. Sustainability 2020, 12, 2010. [Google Scholar] [CrossRef] [Green Version]
- Dan, N.C.; Little, D.C. Overwintering performance of Nile tilapia Oreochromis niloticus (L.) broodfish and seed at ambient temperatures in northern Vietnam. Aquac. Res. 2000, 31, 485–493. [Google Scholar] [CrossRef]
- Prabu, E.; Rajagopalsamy, C.; Ahilan, B.; Jeevagan, I.J.M.A.; Renuhadevi, M. Tilapia—An Excellent Candidate Species for World Aquaculture: A Review. Annu. Res. Rev. Biol. 2019, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Nicholson, P.; Mon-on, N.; Jaemwimol, P.; Tattiyapong, P.; Surachetpong, W. Coinfection of tilapia lake virus and Aeromonas hydrophila synergistically increased mortality and worsened the disease severity in tilapia (Oreochromis spp.). Aquaculture 2020, 520, 734746. [Google Scholar] [CrossRef]
- Piamsomboon, P.; Thanasaksiri, K.; Murakami, A.; Fukuda, K.; Takano, R.; Jantrakajorn, S.; Wongtavatchai, J. Streptococcosis in freshwater farmed seabass Lates calcarifer and its virulence in Nile tilapia Oreochromis niloticus . Aquaculture 2020, 523, 735189. [Google Scholar] [CrossRef]
- Chen, S.-W.; Liu, C.-H.; Hu, S.-Y. Dietary administration of probiotic Paenibacillus ehimensis NPUST1 with bacteriocin-like activity improves growth performance and immunity against Aeromonas hydrophila and Streptococcus iniae in Nile tilapia (Oreochromis niloticus). Fish Shellfish Immunol. 2019, 84, 695–703. [Google Scholar] [CrossRef]
- Rico, A.; Oliveira, R.; McDonough, S.; Matser, A.; Khatikarn, J.; Satapornvanit, K.; Nogueira, A.J.A.; Soares, A.M.V.M.; Domingues, I.; Van den Brink, P.J. Use, fate and ecological risks of antibiotics applied in tilapia cage farming in Thailand. Environ. Pollut. 2014, 191, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Kraemer, S.A.; Ramachandran, A.; Perron, G.G. Antibiotic Pollution in the Environment: From Microbial Ecology to Public Policy. Microorganisms 2019, 7, 180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolivar, R.B.; Cruz, E.V.; Jimenez, E.B.T.; Sayco, R.; Argueza, R.; Ferket, P.; Stark, C.; Malheiros, R.; Ayoola, A.; Johnstone, W. Feeding Reduction Strategies and Alternative Feeds to Reduce Production Costs of Tilapia Culture; Technical Reports: Investigations 2007–2009; AquaFish Collaborative Research; North Carolina State University: Raleigh, CA, USA, 2010. [Google Scholar]
- Kaleem, O.; Bio Singou Sabi, A.-F. Overview of aquaculture systems in Egypt and Nigeria, prospects, potentials, and constraints. Aquac. Fish 2020. [Google Scholar] [CrossRef]
- da Silva Cardoso, A.J.; dos Santos, W.V.; Gomes, J.R.; Martins, M.T.S.; Coura, R.R.; de Almeida Oliveira, M.G.; Salaro, A.L.; de Moraes FrançaFerreira, P.; Carneiro, A.P.S.; Zuanon, J.A.S. Ginger oil, Zingiber officinale, improve palatability, growth and nutrient utilisation efficiency in Nile tilapia fed with excess of starch. Anim. Feed Sci. Technol. 2021, 272, 114756. [Google Scholar] [CrossRef]
- Welengane, E.; Sado, R.Y.; de Almeida Bicudo, Á.J. Protein—sparing effect by dietary lipid increase in juveniles of the hybrid fish tambatinga (♀ Colossoma macropomum×♂ Piaractus brachypomus). Aquac. Nutr. 2019, 25, 1272–1280. [Google Scholar] [CrossRef]
- Phan, L.T.T.; Masagounder, K.; Mas-Muñoz, J.; Schrama, J.W. Differences in energy utilization efficiency of digested protein, fat and carbohydrates in snakehead (Channa striata). Aquaculture 2021, 532, 736066. [Google Scholar] [CrossRef]
- Mohan, K.; Ravichandran, S.; Muralisankar, T.; Uthayakumar, V.; Chandirasekar, R.; Seedevi, P.; Abirami, R.G.; Rajan, D.K. Application of marine-derived polysaccharides as immunostimulants in aquaculture: A review of current knowledge and further perspectives. Fish Shellfish Immunol. 2019, 86, 1177–1193. [Google Scholar] [CrossRef]
- Encarnação, P. 5—Functional feed additives in aquaculture feeds. In Aquafeed Formulation; Nates, S.F., Ed.; Academic Press: San Diego, CA, USA, 2016; pp. 217–237. [Google Scholar] [CrossRef]
- Buruiana, C.-T.; Gómez, B.; Vizireanu, C.; Garrote, G. Manufacture and evaluation of xylooligosaccharides from corn stover as emerging prebiotic candidates for human health. LWT Food Sci. Technol. 2017, 77, 449–459. [Google Scholar] [CrossRef]
- Zhang, W.; You, Y.; Lei, F.; Li, P.; Jiang, J. Acetyl-assisted autohydrolysis of sugarcane bagasse for the production of xylo-oligosaccharides without additional chemicals. Bioresour. Technol. 2018, 265, 387–393. [Google Scholar] [CrossRef]
- Heniegal, A.M.; Ramadan, M.A.; Naguib, A.; Agwa, I.S. Study on properties of clay brick incorporating sludge of water treatment plant and agriculture waste. Case Stud. Constr. Mater. 2020, 13, e00397. [Google Scholar] [CrossRef]
- Biswas, R.; Uellendahl, H.; Ahring, B.K. Wet explosion pretreatment of sugarcane bagasse for enhanced enzymatic hydrolysis. Biomass Bioenergy 2014, 61, 104–113. [Google Scholar] [CrossRef]
- Lachos-Perez, D.; Martinez-Jimenez, F.; Rezende, C.; Tompsett, G.; Timko, M.; Forster-Carneiro, T. Subcritical water hydrolysis of sugarcane bagasse: An approach on solid residues characterization. J. Supercrit. Fluids 2016, 108, 69–78. [Google Scholar] [CrossRef]
- Nunes, L.J.; Loureiro, L.M.; Sá, L.C.; Silva, H.F. Sugarcane industry waste recovery: A case study using thermochemical conversion technologies to increase sustainability. Appl. Sci. 2020, 10, 6481. [Google Scholar] [CrossRef]
- da Silva Braga, R.; Poletto, M. Preparation and Characterization of Hemicellulose Films from Sugarcane Bagasse. Materials 2020, 13, 941. [Google Scholar] [CrossRef] [Green Version]
- Alokika; Anu; Kumar, A.; Kumar, V.; Singh, B. Cellulosic and hemicellulosic fractions of sugarcane bagasse: Potential, challenges and future perspective. Int. J. Biol. Macromol. 2021, 169, 564–582. [Google Scholar] [CrossRef]
- Laluce, C.; Roldan, I.U.; Pecoraro, E.; Igbojionu, L.I.; Ribeiro, C.A. Effects of pretreatment applied to sugarcane bagasse on composition and morphology of cellulosic fractions. Biomass Bioenergy 2019, 126, 231–238. [Google Scholar] [CrossRef]
- Candido, R.G.; Gonçalves, A.R. Evaluation of two different applications for cellulose isolated from sugarcane bagasse in a biorefinery concept. Ind. Crop. Prod. 2019, 142, 111616. [Google Scholar] [CrossRef]
- Kaur, R.; Uppal, S.; Sharma, P. Production of xylooligosaccharides from sugarcane bagasse and evaluation of their prebiotic potency in vitro. Waste Biomass Valorization 2019, 10, 2627–2635. [Google Scholar] [CrossRef]
- Zhao, S.; Zhang, G.-L.; Chen, C.; Yang, Q.; Luo, X.-M.; Wang, Z.-B.; Wu, A.-M.; Feng, J.-X. A combination of mild chemical pre-treatment and enzymatic hydrolysis efficiently produces xylooligosaccharides from sugarcane bagasse. J. Clean. Prod. 2021, 291, 125972. [Google Scholar] [CrossRef]
- Marcondes, W.F.; Milagres, A.M.F.; Arantes, V. Co-production of xylo-oligosaccharides, xylose and cellulose nanofibrils from sugarcane bagasse. J. Biotechnol. 2020, 321, 35–47. [Google Scholar] [CrossRef]
- Gómez-García, R.; Campos, D.A.; Aguilar, C.N.; Madureira, A.R.; Pintado, M. Valorization of melon fruit (Cucumis melo L.) by-products: Phytochemical and Biofunctional properties with Emphasis on Recent Trends and Advances. Trends Food Sci. Technol. 2020, 99, 507–519. [Google Scholar] [CrossRef]
- Hoseinifar, S.H.; Mirvaghefi, A.; Amoozegar, M.A.; Sharifian, M.; Esteban, M.Á. Modulation of innate immune response, mucosal parameters and disease resistance in rainbow trout (Oncorhynchus mykiss) upon synbiotic feeding. Fish Shellfish Immunol. 2015, 45, 27–32. [Google Scholar] [CrossRef]
- Esteban, M.A. An Overview of the Immunological Defenses in Fish Skin. ISRN Immunol. 2012, 2012, 29. [Google Scholar] [CrossRef] [Green Version]
- Hosseini, M.; Kolangi Miandare, H.; Hoseinifar, S.H.; Yarahmadi, P. Dietary Lactobacillus acidophilus modulated skin mucus protein profile, immune and appetite genes expression in gold fish (Carassius auratus gibelio). Fish Shellfish Immunol. 2016, 59, 149–154. [Google Scholar] [CrossRef]
- Rashmeei, M.; Hosseini Shekarabi, S.P.; Shamsaie Mehrgan, M.; Paknejad, H. Stimulatory effect of dietary chasteberry (Vitex agnus-castus) extract on immunity, some immune-related gene expression, and resistance against Aeromonas hydrophila infection in goldfish (Carassius auratus). Fish Shellfish Immunol. 2020, 107, 129–136. [Google Scholar] [CrossRef]
- Fu, G.H.; Liu, F.; Xia, J.H.; Yue, G.H. The LBP gene and its association with resistance to Aeromonas hydrophila in tilapia. Int. J. Mol. Sci. 2014, 15, 22028–22041. [Google Scholar] [CrossRef] [Green Version]
- Hidalgo, M.C.; Exposito, A.; Palma, J.M.; de la Higuera, M. Oxidative stress generated by dietary Zn-deficiency: Studies in rainbow trout (Oncorhynchus mykiss). Int. J. Biochem. Cell Biol. 2002, 34, 183–193. [Google Scholar] [CrossRef]
- Li, Z.-H.; Li, P.; Shi, Z.-C. Responses of the hepatic glutathione antioxidant defense system and related gene expression in juvenile common carp after chronic treatment with tributyltin. Ecotoxicology 2015, 24, 700–705. [Google Scholar] [CrossRef]
- Liu, H.; Li, H.; Wei, H.; Zhu, X.; Han, D.; Jin, J.; Yang, Y.; Xie, S. Biofloc formation improves water quality and fish yield in a freshwater pond aquaculture system. Aquaculture 2019, 506, 256–269. [Google Scholar] [CrossRef]
- Khanjani, M.H.; Sharifinia, M. Biofloc technology as a promising tool to improve aquaculture production. Rev. Aquac. 2020. [Google Scholar] [CrossRef]
- Bossier, P.; Ekasari, J. Biofloc technology application in aquaculture to support sustainable development goals. Microb. Biotechnol. 2017, 10, 1012–1016. [Google Scholar] [CrossRef]
- Jung, J.-Y.; Hur, J.W.; Kim, K.; Han, H.-S. Evaluation of floc-harvesting technologies in biofloc technology (BFT) system for aquaculture. Bioresour. Technol. 2020, 314, 123719. [Google Scholar] [CrossRef]
- Avnimelech, Y. Biofloc Technology: A Practical Guide Book; World Aquaculture Society: Baton Rouge, LA, USA, 2015. [Google Scholar]
- Serradell, A.; Torrecillas, S.; Makol, A.; Valdenegro, V.; Fernández-Montero, A.; Acosta, F.; Izquierdo, M.S.; Montero, D. Prebiotics and phytogenics functional additives in low fish meal and fish oil based diets for European sea bass (Dicentrarchus labrax): Effects on stress and immune responses. Fish Shellfish Immunol. 2020, 100, 219–229. [Google Scholar] [CrossRef]
- Li, Z.; Tran, N.T.; Ji, P.; Sun, Z.; Wen, X.; Li, S. Effects of prebiotic mixtures on growth performance, intestinal microbiota and immune response in juvenile chu’s croaker, Nibea coibor . Fish Shellfish Immunol. 2019, 89, 564–573. [Google Scholar] [CrossRef]
- Mandal, A.; Das, S.K. Comparative efficacy of neem (Azadirachta indica) and non-neem supplemented biofloc media in controlling the harmful luminescent bacteria in natural pond culture of Litopenaeus vannaemei. Aquaculture 2018, 492, 157–163. [Google Scholar] [CrossRef]
- Qiao, G.; Chen, P.; Sun, Q.; Zhang, M.; Zhang, J.; Li, Z.; Li, Q. Poly-β-hydroxybutyrate (PHB) in bioflocs alters intestinal microbial community structure, immune-related gene expression and early Cyprinid herpesvirus 2 replication in gibel carp (Carassius auratus gibelio). Fish Shellfish Immunol. 2020, 97, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Avnimelech, Y. Carbon/nitrogen ratio as a control element in aquaculture systems. Aquaculture 1999, 176, 227–235. [Google Scholar] [CrossRef]
- Cardona, E.; Lorgeoux, B.; Chim, L.; Goguenheim, J.; Le Delliou, H.; Cahu, C. Biofloc contribution to antioxidant defence status, lipid nutrition and reproductive performance of broodstock of the shrimp Litopenaeus stylirostris: Consequences for the quality of eggs and larvae. Aquaculture 2016, 452, 252–262. [Google Scholar] [CrossRef] [Green Version]
- Khodadadian Zou, H.; Hoseinifar, S.H.; Kolangi Miandare, H.; Hajimoradloo, A. Agaricus bisporus powder improved cutaneous mucosal and serum immune parameters and up-regulated intestinal cytokines gene expression in common carp (Cyprinus carpio) fingerlings. Fish Shellfish Immunol. 2016, 58, 380–386. [Google Scholar] [CrossRef]
- Van Doan, H.; Hoseinifar, S.H.; Tapingkae, W.; Tongsiri, S.; Khamtavee, P. Combined administration of low molecular weight sodium alginate boosted immunomodulatory, disease resistance and growth enhancing effects of Lactobacillus plantarum in Nile tilapia (Oreochromis niloticus). Fish Shellfish Immunol. 2016, 58, 678–685. [Google Scholar] [CrossRef]
- Van Doan, H.; Tapingkae, W.; Moonmanee, T.; Seepai, A. Effects of low molecular weight sodium alginate on growth performance, immunity, and disease resistance of tilapia, Oreochromis niloticus . Fish Shellfish Immunol. 2016, 55, 186–194. [Google Scholar] [CrossRef]
- Parry, R.M., Jr.; Chandan, R.C.; Shahani, K.M. A rapid and sensitive assay of muramidase. Proc. Soc. Exp. Biol. Medicine. Soc. Exp. Biol. Med. (N. Y.) 1965, 119, 384–386. [Google Scholar] [CrossRef]
- Van Doan, H.; Hoseinifar, S.H.; Dawood, M.A.O.; Chitmanat, C.; Tayyamath, K. Effects of Cordyceps militaris spent mushroom substrate and Lactobacillus plantarum on mucosal, serum immunology and growth performance of Nile tilapia (Oreochromis niloticus). Fish Shellfish Immunol. 2017, 70, 87–94. [Google Scholar] [CrossRef]
- Secomebs, C.J. Isolation of salmonid macrophage and analysis of their killing ability. In Techniques in Fish Immunology; Stolen, J.S., Fletcher, T.C., Anderson, D.P., Roberson, B.S., Van Muiswinkel, W.B., Eds.; SOS Publication: Fair Haven, NJ, USA, 1990; pp. 137–152. [Google Scholar]
- Doan, H.V.; Hoseinifar, S.H.; Jaturasitha, S.; Dawood, M.A.O.; Harikrishnan, R. The effects of berberine powder supplementation on growth performance, skin mucus immune response, serum immunity, and disease resistance of Nile tilapia (Oreochromis niloticus) fingerlings. Aquaculture 2020, 520, 734927. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- SAS. Statistical Analysis; SAS Institute Inc.: Cary, NC, USA, 2003. [Google Scholar]
- Brinchmann, M.F. Immune relevant molecules identified in the skin mucus of fish using-omics technologies. Mol. BioSystems 2016, 12, 2056–2063. [Google Scholar] [CrossRef] [Green Version]
- Reverter, M.; Tapissier-Bontemps, N.; Lecchini, D.; Banaigs, B.; Sasal, P. Biological and ecological roles of external fish mucus: A review. Fishes 2018, 3, 41. [Google Scholar] [CrossRef] [Green Version]
- Kulczykowska, E. Stress Response System in the Fish Skin—Welfare Measures Revisited. Front. Physiol. 2019, 10, 72. [Google Scholar] [CrossRef] [Green Version]
- Dawood, M.A.O. Effect of Various Feed Additives on the Performance of Aquatic Animals; Kagoshima University: Kagoshima, Japan, 2016. [Google Scholar]
- Pietrzak, E.; Mazurkiewicz, J.; Slawinska, A. Innate Immune Responses of Skin Mucosa in Common Carp (Cyprinus Carpio) Fed a Diet Supplemented with Galactooligosaccharides. Animals 2020, 10, 438. [Google Scholar] [CrossRef] [Green Version]
- Dash, S.; Das, S.K.; Samal, J.; Thatoi, H.N. Epidermal mucus, a major determinant in fish health: A review. Iran. J. Vet. Res. 2018, 19, 72–81. [Google Scholar]
- Hoseinifar, S.H.; Jahazi, M.A.; Nikdehghan, N.; Van Doan, H.; Volpe, M.G.; Paolucci, M. Effects of dietary polyphenols from agricultural by-products on mucosal and humoral immune and antioxidant responses of convict cichlid (Amatitlania nigrofasciata). Aquaculture 2020, 517, 734790. [Google Scholar] [CrossRef]
- García Beltrán, J.M.; Silvera, D.G.; Ruiz, C.E.; Campo, V.; Chupani, L.; Faggio, C.; Esteban, M.Á. Effects of dietary Origanum vulgare on gilthead seabream (Sparus aurata L.) immune and antioxidant status. Fish Shellfish Immunol. 2020, 99, 452–461. [Google Scholar] [CrossRef]
- Ceballos-Francisco, D.; Castillo, Y.; De La Rosa, F.; Vásquez, W.; Reyes-Santiago, R.; Cuello, A.; Cuesta, A.; Esteban, M.Á. Bactericidal effect on skin mucosa of dietary guava (Psidium guajava L.) leaves in hybrid tilapia (Oreochromis niloticus × O. mossambicus). J. Ethnopharmacol. 2020, 259, 112838. [Google Scholar] [CrossRef]
- Hosseini, S.M.; Hoseinifar, S.H.; Mazandarani, M.; Paknejad, H.; Van Doan, H.; El-Haroun, E.R. The potential benefits of orange peels derived pectin on serum and skin mucus immune parameters, antioxidant defence and growth performance in common carp (Cyprinus carpio). Fish Shellfish Immunol. 2020, 103, 17–22. [Google Scholar] [CrossRef]
- Yousefi, S.; Monsef Shokri, M.; Allaf Noveirian, H.; Hoseinifar, S.H. Effects of dietary yeast cell wall on biochemical indices, serum and skin mucus immune responses, oxidative status and resistance against Aeromonas hydrophila in juvenile Persian sturgeon (Acipenser persicus). Fish Shellfish Immunol. 2020, 106, 464–472. [Google Scholar] [CrossRef] [PubMed]
- Van Doan, H.; Lumsangkul, C.; Hoseinifar, S.H.; Hung, T.Q.; Stejskal, V.; Ringø, E.; Dawood, M.A.O.; Esteban, M.Á. Administration of watermelon rind powder to Nile tilapia (Oreochromis niloticus) culture under biofloc system: Effect on growth performance, innate immune response, and disease resistance. Aquaculture 2020. [Google Scholar] [CrossRef]
- Van Doan, H.; Hoseinifar, S.H.; Harikrishnan, R.; Khamlor, T.; Punyatong, M.; Tapingkae, W.; Yousefi, M.; Palma, J.; El-Haroun, E. Impacts of pineapple peel powder on growth performance, innate immunity, disease resistance, and relative immune gene expression of Nile tilapia, Oreochromis niloticus. Fish Shellfish Immunol. 2021. [Google Scholar] [CrossRef]
- Ghodrati, M.; Rajabi Islami, H.; Hosseini Shekarabi, S.P.; Shenavar Masouleh, A.; Shamsaie Mehrgan, M. Combined effects of enzymes and probiotics on hemato-biochemical parameters and immunological responses of juvenile Siberian sturgeon (Acipenser baerii). Fish Shellfish Immunol. 2021, 112, 116–124. [Google Scholar] [CrossRef]
- Magnadottir, B. Immunological control of fish diseases. Mar. Biotechnol. 2010, 12, 361–379. [Google Scholar] [CrossRef]
- Neumann, N.F.; Stafford, J.L.; Barreda, D.; Ainsworth, A.J.; Belosevic, M. Antimicrobial mechanisms of fish phagocytes and their role in host defense. Dev. Comp. Immunol. 2001, 25, 807–825. [Google Scholar] [CrossRef]
- Thomas, D.C. The phagocyte respiratory burst: Historical perspectives and recent advances. Immunol Lett. 2017, 192, 88–96. [Google Scholar] [CrossRef]
- Zhang, X.; Sun, Z.; Cai, J.; Wang, J.; Wang, G.; Zhu, Z.; Cao, F. Effects of dietary fish meal replacement by fermented moringa (Moringa oleifera Lam.) leaves on growth performance, nonspecific immunity and disease resistance against Aeromonas hydrophila in juvenile gibel carp (Carassius auratus gibelio var. CAS III). Fish Shellfish Immunol. 2020, 102, 430–439. [Google Scholar] [CrossRef]
- Ye, G.; Dong, X.; Yang, Q.; Chi, S.; Liu, H.; Zhang, H.; Tan, B.; Zhang, S. Low-gossypol cottonseed protein concentrate used as a replacement of fish meal for juvenile hybrid grouper (Epinephelus fuscoguttatus ♀ × Epinephelus lanceolatus ♂): Effects on growth performance, immune responses and intestinal microbiota. Aquaculture 2020, 524, 735309. [Google Scholar] [CrossRef]
- Abdel-Latif, H.M.R.; Abdel-Tawwab, M.; Khalil, R.H.; Metwally, A.A.; Shakweer, M.S.; Ghetas, H.A.; Khallaf, M.A. Black soldier fly (Hermetia illucens) larvae meal in diets of European seabass: Effects on antioxidative capacity, non-specific immunity, transcriptomic responses, and resistance to the challenge with Vibrio alginolyticus. Fish Shellfish Immunol. 2021, 111, 111–118. [Google Scholar] [CrossRef]
- Colombo, R.; Lanças, F.M.; Yariwake, J.H. Determination of flavonoids in cultivated sugarcane leaves, bagasse, juice and in transgenic sugarcane by liquid chromatography-UV detection. J. Chromatogr. A 2006, 1103, 118–124. [Google Scholar] [CrossRef]
- Zheng, R.; Su, S.; Li, J.; Zhao, Z.; Wei, J.; Fu, X.; Liu, R.H. Recovery of phenolics from the ethanolic extract of sugarcane (Saccharum officinarum L.) baggase and evaluation of the antioxidant and antiproliferative activities. Ind. Crop. Prod. 2017, 107, 360–369. [Google Scholar] [CrossRef]
- Shakoor, H.; Feehan, J.; Apostolopoulos, V.; Platat, C.; Al Dhaheri, A.S.; Ali, H.I.; Ismail, L.C.; Bosevski, M.; Stojanovska, L. Immunomodulatory Effects of Dietary Polyphenols. Nutrients 2021, 13, 728. [Google Scholar] [CrossRef]
- Wang, T.; Secombes, C.J. The cytokine networks of adaptive immunity in fish. Fish Shellfish Immunol. 2013, 35, 1703–1718. [Google Scholar] [CrossRef]
- Sakai, M.; Hikima, J.-i.; Kono, T. Fish cytokines: Current research and applications. Fish. Sci. 2021, 87, 1–9. [Google Scholar] [CrossRef]
- Rebl, A.; Goldammer, T. Under control: The innate immunity of fish from the inhibitors’ perspective. Fish Shellfish Immunol. 2018, 77, 328–349. [Google Scholar] [CrossRef]
- Siddik, M.A.B.; Chaklader, M.R.; Foysal, M.J.; Howieson, J.; Fotedar, R.; Gupta, S.K. Influence of fish protein hydrolysate produced from industrial residues on antioxidant activity, cytokine expression and gut microbial communities in juvenile barramundi Lates calcarifer. Fish Shellfish Immunol. 2020, 97, 465–473. [Google Scholar] [CrossRef]
- Khoa, T.N.D.; Waqalevu, V.; Honda, A.; Matsui, H.; Truong, N.X.; Sakaguchi, K.; Kawaji, H.; Ishikawa, M.; Shiozaki, K.; Kotani, T. Enrichment effects of fermented by-product of Shochu distillery on Brachionus plicatilis sp. rotifer and larviculture performance in Japanese flounder (Paralichthys olivaceus). Aquaculture 2021, 535, 736352. [Google Scholar] [CrossRef]
- Harikrishnan, R.; Devi, G.; Van Doan, H.; Balasundaram, C.; Esteban, M.Á.; Abdel-Tawwab, M. Impact of grape pomace flour (GPF) on immunity and immune-antioxidant-anti-inflammatory genes expression in Labeo rohita against Flavobacterium columnaris. Fish Shellfish Immunol. 2021, 111, 69–82. [Google Scholar] [CrossRef]
- Bi, D.; Wang, Y.; Gao, Y.; Li, X.; Chu, Q.; Cui, J.; Xu, T. Recognition of Lipopolysaccharide and Activation of NF-κB by Cytosolic Sensor NOD1 in Teleost Fish. Front. Immunol. 2018, 9, 1413. [Google Scholar] [CrossRef] [Green Version]
- Lu, X.-J.; Chu, C.-Q.; Chen, Q.; Chen, J. A novel lipopolysaccharide-binding protein (LBP) gene from sweetfish Plecoglossus altivelis: Molecular characterization and its role in the immune response of monocytes/macrophages. Fish Shellfish Immunol. 2014, 38, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Raetz, C.R.; Whitfield, C. Lipopolysaccharide endotoxins. Annu. Rev. Biochem. 2002, 71, 635–700. [Google Scholar] [CrossRef] [Green Version]
- Guttvik, A.; Paulsen, B.; Dalmo, R.A.; Espelid, S.; Lund, V.; Bøgwald, J. Oral administration of lipopolysaccharide to Atlantic salmon (Salmo salar L.) fry. Uptake, distribution, influence on growth and immune stimulation. Aquaculture 2002, 214, 35–53. [Google Scholar] [CrossRef]
- Imai, H.; Nakagawa, Y. Biological significance of phospholipid hydroperoxide glutathione peroxidase (PHGPx, GPx4) in mammalian cells. Free Radic. Biol. Med. 2003, 34, 145–169. [Google Scholar] [CrossRef]
- Diamond, J. Sediment Toxicity Assessment; JSTOR: New York, NY, USA, 1993. [Google Scholar]
- Manal, I. Detoxification and antioxidant effects of garlic and curcumin in Oreochromis niloticus injected with aflatoxin B 1 with reference to gene expression of glutathione peroxidase (GPx) by RT-PCR. Fish Physiol. Biochem. 2016, 42, 617–629. [Google Scholar]
- Caxico Vieira, C.A.S.; Vieira, J.S.; Bastos, M.S.; Zancanela, V.; Barbosa, L.T.; Gasparino, E.; Del Vesco, A.P. Expression of genes related to antioxidant activity in Nile tilapia kept under salinity stress and fed diets containing different levels of vitamin C. J. Toxicol. Environ. Health A 2018, 81, 20–30. [Google Scholar] [CrossRef]
- Abdelazim, A.M.; Saadeldin, I.M.; Swelum, A.A.-A.; Afifi, M.M.; Alkaladi, A. Oxidative stress in the muscles of the fish Nile tilapia caused by zinc oxide nanoparticles and its modulation by vitamins C and E. Oxidative Med. Cell. Longev. 2018, 2018, 6926712. [Google Scholar] [CrossRef]
- Tan, X.; Sun, Z.; Ye, C.; Lin, H. The effects of dietary Lycium barbarum extract on growth performance, liver health and immune related genes expression in hybrid grouper (Epinephelus lanceolatus♂ × E. fuscoguttatus♀) fed high lipid diets. Fish Shellfish Immunol. 2019, 87, 847–852. [Google Scholar] [CrossRef]
- Safari, R.; Hoseinifar, S.H.; Dadar, M.; Nejadmoghaddam, S.; Van Doan, H. Effect of dietary sodium acetate on skin mucus immune parameters and expression of gene related to growth, immunity and antioxidant system in common carp (Cyprinus carpio) intestine. Ann. Anim. Sci. 2020, 20, 1441–1452. [Google Scholar] [CrossRef]
- Hoseinifar, S.H.; Khodadadian Zou, H.; Paknejad, H.; Hajimoradloo, A.; Van Doan, H. Effects of dietary white-button mushroom powder on mucosal immunity, antioxidant defence, and growth of common carp (Cyprinus carpio). Aquaculture 2019, 501, 448–454. [Google Scholar] [CrossRef]
- Hoseini, S.M.; Khalili, M.; Rajabiesterabadi, H.; Hoseinifar, S.H.; Doan, H.V. Effects of dietary monoterpene, myrcene, administration on immune- and health-related genes expression in common carp gill following exposure to copper sulfate. Fish Shellfish. Immunol. 2020, 98, 438–445. [Google Scholar] [CrossRef]
- Peixoto, M.J.; Ferraz, R.; Magnoni, L.J.; Pereira, R.; Gonçalves, J.F.; Calduch-Giner, J.; Pérez-Sánchez, J.; Ozório, R.O.A. Protective effects of seaweed supplemented diet on antioxidant and immune responses in European seabass (Dicentrarchus labrax) subjected to bacterial infection. Sci. Rep. 2019, 9, 16134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sporck, D.; Reinoso, F.A.M.; Rencoret, J.; Gutiérrez, A.; del Rio, J.C.; Ferraz, A.; Milagres, A.M.F. Xylan extraction from pretreated sugarcane bagasse using alkaline and enzymatic approaches. Biotechnol. Biofuels 2017, 10, 296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, S.-K.; Jung, C.-D.; Yu, J.-H.; Kim, H. Environmentally Friendly Approach for the Production of Glucose and High-Purity Xylooligosaccharides from Edible Biomass Byproducts. Appl. Sci. 2020, 10, 8119. [Google Scholar] [CrossRef]
- Reddy, S.S.; Krishnan, C. Production of high-pure xylooligosaccharides from sugarcane bagasse using crude β-xylosidase-free xylanase of Bacillus subtilis KCX006 and their bifidogenic function. LWT-Food Sci. Technol. 2016, 65, 237–245. [Google Scholar] [CrossRef]
- Chen, H.H.; Chen, Y.K.; Chang, H.C.; Lin, S.Y. Immunomodulatory effects of xylooligosaccharides. Food Sci. Technol. Res. 2012, 18, 195–199. [Google Scholar] [CrossRef] [Green Version]
- Palaniappan, A.; Antony, U.; Emmambux, M.N. Current status of xylooligosaccharides: Production, characterization, health benefits and food application. Trends Food Sci. Technol. 2021, 111, 506–519. [Google Scholar] [CrossRef]
- Van Doan, H.; Hoseinifar, S.H.; Faggio, C.; Chitmanat, C.; Mai, N.T.; Jaturasitha, S.; Ringø, E. Effects of corncob derived xylooligosaccharide on innate immune response, disease resistance, and growth performance in Nile tilapia (Oreochromis niloticus) fingerlings. Aquaculture 2018, 495, 786–793. [Google Scholar] [CrossRef]
- Zhang, Z.-H.; Chen, M.; Xie, S.-W.; Chen, X.-Q.; Liu, Y.-J.; Tian, L.-X.; Niu, J. Effects of dietary xylooligosaccharide on growth performance, enzyme activity and immunity of juvenile grass carp, Ctenopharyngodon idellus. Aquac. Rep. 2020, 18, 100519. [Google Scholar] [CrossRef]
- Afanasjeva, N. Obtaining phenolic compounds by sugar cane bagasse pyrolysis and its antioxidant capacity measured through chemical and electrochemical methods. J. Phys. Conf. Ser. 2018, 1119, 012013. [Google Scholar] [CrossRef]
- Velazquez-Martinez, V.; Valles-Rosales, D.; Rodriguez-Uribe, L.; Holguin, O.; Quintero-Quiroz, J.; Reyes-Jaquez, D.; Rodriguez-Borbon, M.I.; Villagrán-Villegas, L.Y.; Delgado, E. Antimicrobial, Shelf-Life Stability, and Effect of Maltodextrin and Gum Arabic on the Encapsulation Efficiency of Sugarcane Bagasse Bioactive Compounds. Foods 2021, 10, 116. [Google Scholar] [CrossRef]
- Zheng, R.; Su, S.; Zhou, H.; Yan, H.; Ye, J.; Zhao, Z.; You, L.; Fu, X. Antioxidant/antihyperglycemic activity of phenolics from sugarcane (Saccharum officinarum L.) bagasse and identification by UHPLC-HR-TOFMS. Ind. Crop. Prod. 2017, 101, 104–114. [Google Scholar] [CrossRef]
- Giri, S.S.; Jun, J.W.; Sukumaran, V.; Park, S.C. Dietary Administration of Banana (Musa acuminata) Peel Flour Affects the Growth, Antioxidant Status, Cytokine Responses, and Disease Susceptibility of Rohu, Labeo rohita. J. Immunol. Res. 2016, 2016, 4086591. [Google Scholar] [CrossRef] [Green Version]
- Wu, N.; Song, Y.-L.; Wang, B.; Zhang, X.-Y.; Zhang, X.-J.; Wang, Y.-L.; Cheng, Y.-Y.; Chen, D.-D.; Xia, X.-Q.; Lu, Y.-S.; et al. Fish gut-liver immunity during homeostasis or inflammation revealed by integrative transcriptome and proteome studies. Sci. Rep. 2016, 6, 36048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hermesz, E.; Ferencz, Á. Identification of two phospholipid hydroperoxide glutathione peroxidase (gpx4) genes in common carp. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2009, 150, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Mengistu, S.B.; Mulder, H.A.; Benzie, J.A.H.; Komen, H. A systematic literature review of the major factors causing yield gap by affecting growth, feed conversion ratio and survival in Nile tilapia (Oreochromis niloticus). Rev. Aquac. 2020, 12, 524–541. [Google Scholar] [CrossRef] [Green Version]
- Dawood, M.A.O.; Koshio, S. Recent advances in the role of probiotics and prebiotics in carp aquaculture: A review. Aquaculture 2016, 454, 243–251. [Google Scholar] [CrossRef]
- Reverter, M.; Tapissier-Bontemps, N.; Sasal, P.; Saulnier, D. Use of Medicinal Plants in Aquaculture. In Diagnosis and Control of Diseases of Fish and Shellfish; Austin, B., Newaj-Fyzul, A., Eds.; Wiley Online Library: Hoboken, NJ, USA, 2017. [Google Scholar] [CrossRef]
- Dharmaraj, M.; Manissery, J.K.; Keshavanath, P. Effects of a biodegradable substrate, sugarcane bagasse and supplemental feed on growth and production of fringe-lipped peninsula carp, Labeo fimbriatus [Bloch]. Acta Ichthyol. Piscat. 2002, 2. [Google Scholar] [CrossRef] [Green Version]
- de Almeida, G.A.P.; de Andrade Ferreira, M.; de Lima Silva, J.; Chagas, J.C.C.; Véras, A.S.C.; de Barros, L.J.A.; de Almeida, G.L.P. Sugarcane bagasse as exclusive roughage for dairy cows in smallholder livestock system. Asian-Australas. J. Anim. Sci. 2018, 31, 379. [Google Scholar] [CrossRef] [Green Version]
- Kheravii, S.K.; Swick, R.A.; Choct, M.; Wu, S.-B. Dietary sugarcane bagasse and coarse particle size of corn are beneficial to performance and gizzard development in broilers fed normal and high sodium diets. Poult. Sci. 2017, 96, 4006–4016. [Google Scholar] [CrossRef]
- Valladares-Diestra, K.K.; Porto de Souza Vandenberghe, L.; Soccol, C.R. A biorefinery approach for enzymatic complex production for the synthesis of xylooligosaccharides from sugarcane bagasse. Bioresour. Technol. 2021, 333, 125174. [Google Scholar] [CrossRef]
- Sun, C.-Y.; Liu, Y.; Feng, L.; Jiang, W.-D.; Wu, P.; Jiang, J.; Kuang, S.-Y.; Tang, L.; Zhou, X.-Q. Xylooligosaccharide supplementation improved growth performance and prevented intestinal apoptosis in grass carp. Aquaculture 2021, 535, 736360. [Google Scholar] [CrossRef]
- García-Ríos, L.; Miranda-Baeza, A.; Coelho-Emerenciano, M.G.; Huerta-Rábago, J.A.; Osuna-Amarillas, P. Biofloc technology (BFT) applied to tilapia fingerlings production using different carbon sources: Emphasis on commercial applications. Aquaculture 2019, 502, 26–31. [Google Scholar] [CrossRef]
- Laice, L.M.; Corrêa Filho, R.A.C.; Ventura, A.S.; Farias, K.N.N.; Silva, A.L.d.N.; Fernandes, C.E.; Silva, A.C.F.; Barbosa, P.T.L.; de Souza, A.I.; Emerenciano, M.G.C.; et al. Use of symbiotics in biofloc (BFT)-based Nile tilapia culture: Production performance, intestinal morphometry and hematological parameters. Aquaculture 2021, 530, 735715. [Google Scholar] [CrossRef]
- de Sousa, A.A.; Pinho, S.M.; Rombenso, A.N.; de Mello, G.L.; Emerenciano, M.G.C. Pizzeria by-product: A complementary feed source for Nile tilapia (Oreochromis niloticus) raised in biofloc technology? Aquaculture 2019, 501, 359–367. [Google Scholar] [CrossRef]
- Kishawy, A.T.; Sewid, A.H.; Nada, H.S.; Kamel, M.A.; El-Mandrawy, S.A.; Abdelhakim, T.; El-Murr, A.E.I.; Nahhas, N.E.; Hozzein, W.N.; Ibrahim, D. Mannanoligosaccharides as a carbon source in Biofloc boost dietary plant protein and water quality, growth, immunity and Aeromonas hydrophila resistance in Nile tilapia (Oreochromis niloticus). Animals 2020, 10, 1724. [Google Scholar] [CrossRef]
- Ameram, N.; Muhammad, S.; Yusof, N.; Ishak, S.; Ali, A.; Shoparwe, N.F.; Ter, T.P. Chemical composition in sugarcane bagasse: Delignification with sodium hydroxide. Malays. J. Fundam. Appl. Sci 2019, 15, 232–236. [Google Scholar] [CrossRef]
- Manzoor, M.; Jabeen, F.; Thomas-Hall, S.R.; Altaf, J.; Younis, T.; Schenk, P.M.; Qazi, J.I. Sugarcane bagasse as a novel low/no cost organic carbon source for growth of Chlorella sp. BR2. Biofuels 2019, 1–7. [Google Scholar] [CrossRef]
- Di Marco, E.; Soraire, P.M.; Romero, C.M.; Villegas, L.B.; Martínez, M.A. Raw sugarcane bagasse as carbon source for xylanase production by Paenibacillus species: A potential degrader of agricultural wastes. Environ. Sci. Pollut. Res. 2017, 24, 19057–19067. [Google Scholar] [CrossRef]
- Nguyen, H.C.; Su, C.-H.; Yu, Y.-K.; Huong, D.T.M. Sugarcane bagasse as a novel carbon source for heterotrophic cultivation of oleaginous microalga Schizochytrium sp. Ind. Crop. Prod. 2018, 121, 99–105. [Google Scholar] [CrossRef]
- Emerenciano, M.; Cuzon, G.; Paredes, A.; Gaxiola, G. Evaluation of biofloc technology in pink shrimp Farfantepenaeus duorarum culture: Growth performance, water quality, microorganisms profile and proximate analysis of biofloc. Aquacult. Int. 2013, 21, 1381–1394. [Google Scholar] [CrossRef]
- Emerenciano, M.G.C.; Martínez-Córdova, L.R.; Martínez-Porchas, M.; Miranda-Baeza, A. Biofloc technology (BFT): A tool for water quality management in aquaculture. In Water Quality; IntechOpen: London, UK, 2017; pp. 91–109. [Google Scholar]
- Torrecillas, S.; Montero, D.; Izquierdo, M. Improved health and growth of fish fed mannan oligosaccharides: Potential mode of action. Fish Shellfish Immunol. 2014, 36, 525–544. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Fu, G.; Dong, H.; Li, J.; Du, Y.; Zhang, D. High-efficiency secretion of β-mannanase in Bacillus subtilis through protein synthesis and secretion optimization. J. Agric. Food Chem. 2017, 65, 2540–2548. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Ingredients | SB0 | SB10 | SB20 | SB40 | SB80 |
|---|---|---|---|---|---|
| Fish meal | 150 | 150 | 150 | 150 | 150 |
| Corn meal | 200 | 200 | 200 | 200 | 200 |
| Soybean meal | 390 | 390 | 390 | 394 | 400 |
| Wheat flour | 70 | 70 | 70 | 70 | 70 |
| Rice bran | 150 | 150 | 145 | 126 | 80 |
| SB 1 | 0 | 10 | 20 | 40 | 80 |
| Cellulose | 20 | 10 | 5 | 0 | 0 |
| Soybean oil | 5 | 5 | 5 | 5 | 5 |
| Premix 2 | 10 | 10 | 10 | 10 | 10 |
| Vitamin C 3 | 5 | 5 | 5 | 5 | 5 |
| Proximate composition of the experimental diets (%) | |||||
| Crude protein | 32.6 | 32.4 | 32 | 31.4 | 31.8 |
| Crude lipid | 2.34 | 2.57 | 2.69 | 3.58 | 2.68 |
| Fiber | 3.75 | 3.83 | 4.35 | 4.76 | 5.06 |
| Ash | 7.75 | 7.72 | 7.67 | 7.50 | 7.34 |
| Dry matter | 96.63 | 96.80 | 94.02 | 93.97 | 96.8 |
| GE (cal/g) 4 | 4239 | 4255 | 4200 | 4214 | 4219 |
| Primer Name | Primer Sequence (5′-3′) | Target Gene | Tm (°C) | Product Size (bp) | Accession No. |
|---|---|---|---|---|---|
| 18S rRNA -F | GTGCATGGCCGTTCTTAGTT | 18S rRNA | 60 | 150 | XR_003216134 |
| 18S rRNA -R | CTCAATCTCGTGTGGCTGAA | 60 | |||
| IL1-F | GTCTGTCAAGGATAAGCGCTG | IL-1 | 59 | 200 | XM_019365844 |
| IL1-R | ACTCTGGAGCTGGATGTTGA | 58 | |||
| IL8-F | CTGTGAAGGCATGGGTGTG | IL-8 | 59 | 196 | NM_001279704 |
| IL8-R | ATCACTTTCTTCACCCAGGG | 58 | |||
| LBP-F | ACCAGAAACTGCGAGAAGGA | LBP | 59 | 200 | XM_013271147 |
| LBP-R | GATTGGTGGTCGGAGGTTTG | 59 | |||
| GSTa-F | ACTGCACACTCATGGGAACA | GSTa | 60 | 190 | NM_001279635 |
| GSTa-R | TTAAAAGCCAGCGGATTGAC | 60 | |||
| GPX-F | GGTGGATGTGAATGGAAAGG | GPX | 60 | 190 | NM_001279711 |
| GPX-R | CTTGTAAGGTTCCCCGTCAG | 59 | |||
| GSR-F | CTGCACCAAAGAACTGCAAAC | GSR | 60 | 172 | XM_005467348 |
| GSR-R | CAGAGAAGGCAGTCCACTC | 60 |
| Ingredients | SB0 | SB10 | SB20 | SB40 | SB80 |
|---|---|---|---|---|---|
| IW (g) | 15.12 ± 0.007 | 15.12 ± 0.01 | 15.17 ± 0.01 | 15.10 ± 0.01 | 15.07 ± 0.004 |
| FW (g) | |||||
| 4 weeks | 36.65 ± 0.21 | 36.23 ± 0.13 | 37.25 ± 0.11 | 37.43 ± 0.20 | 39.00 ± 0.25 |
| 8 weeks | 71.48 ± 0.20 b | 71.35 ± 0.13 b | 74.78 ± 0.04 a | 75.60 ± 0.10 a | 71.90 ± 0.01 b |
| WG (g) | |||||
| 4 weeks | 21.53 ± 0.20 | 21.12 ± 0.43 | 22.08 ± 0.12 | 22.33 ± 0.20 | 23.93 ± 0.25 |
| 8 weeks | 56.83 ± 0.20 b | 56.23 ± 0.13 b | 59.62 ± 0.05 a | 60.50 ± 0.10 a | 56.83 ± 0.02 b |
| FCR | |||||
| 4 weeks | 1.05 ± 0.006 | 1.08 ± 0.02 | 1.08 ± 0.002 | 1.04 ± 0.009 | 0.97 ± 0.009 |
| 8 weeks | 1.23 ± 0.003 b | 1.23 ± 0.006 b | 1.22 ± 0.003 b | 1.18 ± 0.001 b | 1.29 ± 0.003 a |
| SGR | |||||
| 4 weeks | 3.16 ± 0.02 | 3.11 ± 0.04 | 3.21 ± 0.01 | 3.24 ± 0.02 | 3.39 ± 0.02 |
| 8 weeks | 2.77 ± 0.004 b | 2.77 ± 0.003 b | 2.85 ± 0.002 a | 2.88 ± 0.003 a | 2.79 ± 0.00 b |
| SR (%) | |||||
| 4 weeks | 100 | 100 | 100 | 100 | 100 |
| 8 weeks | 100 | 100 | 100 | 100 | 100 |
| Ingredients | SB0 | SB10 | SB20 | SB40 | SB80 |
|---|---|---|---|---|---|
| 4 weeks | |||||
| SMLA | 0.73 ± 0.08 b | 1.22 ± 0.06 ab | 1.64 ± 0.41 ab | 1.74 ± 0.43 a | 1.73a ± 0.17 a |
| SMPA | 0.08 ± 0.006 b | 0.09 ± 0.005 ab | 0.10 ± 0.003 a | 0.09 ± 0.005 ab | 0.10 ± 0.007 a |
| 8 weeks | |||||
| SMLA | 1.50 ± 0.31 c | 2.89 ± 066 ab | 3.73 ± 0.95 a | 2.61 ± 0.90 b | 3.49 ± 1.19 ab |
| SMPA | 0.09 ± 0.006 b | 0.14 ± 0.006 a | 0.12 ± 0.001 a | 0.14 ± 0.003 a | 0.15 ± 0.02 a |
| Ingredients | SB0 | SB10 | SB20 | SB40 | SB80 |
|---|---|---|---|---|---|
| 4 weeks | |||||
| SL | 1.90 ± 0.29 c | 2.36 ± 0.02 bc | 3.20 ± 0.25 a | 2.87 ± 0.09 ab | 3.18 ± 0.35 a |
| SP | 0.22 ± 0.02 | 0.22 ± 0.07 | 0.28 ± 0.04 | 0.26 ± 0.04 | 0.21 ± 0.02 |
| RB | 0.12 ± 0.008 b | 0.21 ± 0.02 a | 0.10 ± 0.004 b | 0.14 ± 0.02 b | 0.12 ± 0.10 b |
| 8 weeks | |||||
| SL | 5.69 ± 0.41 b | 8.68 ± 1.09 a | 8.42 ± 1.04 a | 8.72 ± 0.32 a | 7.88 ± 0.38 ab |
| SP | 0.19 ± 0.01 | 0.16 ± 0.01 | 0.17 ± 0.02 | 0.20 ± 0.02 | 0.17 ± 0.02 |
| RB | 0.25 ± 0.02 | 0.27 ± 0.006 | 0.18 ± 0.02 | 0.20 ± 0.005 | 0.25 ± 0.07 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lumsangkul, C.; Tapingkae, W.; Sringarm, K.; Jaturasitha, S.; Le Xuan, C.; Wannavijit, S.; Outama, P.; Van Doan, H. Effect of Dietary Sugarcane Bagasse Supplementation on Growth Performance, Immune Response, and Immune and Antioxidant-Related Gene Expressions of Nile Tilapia (Oreochromis niloticus) Cultured under Biofloc System. Animals 2021, 11, 2035. https://doi.org/10.3390/ani11072035
Lumsangkul C, Tapingkae W, Sringarm K, Jaturasitha S, Le Xuan C, Wannavijit S, Outama P, Van Doan H. Effect of Dietary Sugarcane Bagasse Supplementation on Growth Performance, Immune Response, and Immune and Antioxidant-Related Gene Expressions of Nile Tilapia (Oreochromis niloticus) Cultured under Biofloc System. Animals. 2021; 11(7):2035. https://doi.org/10.3390/ani11072035
Chicago/Turabian StyleLumsangkul, Chompunut, Wanaporn Tapingkae, Korawan Sringarm, Sanchai Jaturasitha, Chinh Le Xuan, Supreya Wannavijit, Piyatida Outama, and Hien Van Doan. 2021. "Effect of Dietary Sugarcane Bagasse Supplementation on Growth Performance, Immune Response, and Immune and Antioxidant-Related Gene Expressions of Nile Tilapia (Oreochromis niloticus) Cultured under Biofloc System" Animals 11, no. 7: 2035. https://doi.org/10.3390/ani11072035