
Evaluation of Tannin Extracts, Leonardite and Tributyrin Supplementation on Diarrhoea Incidence and Gut Microbiota of Weaned Piglets

 ,
,  , , , ,
, , , , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Housing, Experimental Design and Treatment
2.2. Chemical Evaluation of Experimental Diets
2.3. Zootechnical Performance, Diarrhoea Incidence and Sampling Procedures
2.4. Blood Serum Analysis
2.5. Microbiological Analysis of Faecal Samples
2.6. Nitrogen Content, Apparent Nitrogen Digestibility, Volatile Fatty Acids and pH of Faecal Samples
2.7. Bacterial DNA Extraction, V3-V4 Region Amplification and Sequencing
2.8. Statistical Analysis
3. Results and Discussion
3.1. Chemical Evaluation of the Experimental Diets
3.2. Zootechnical Performance
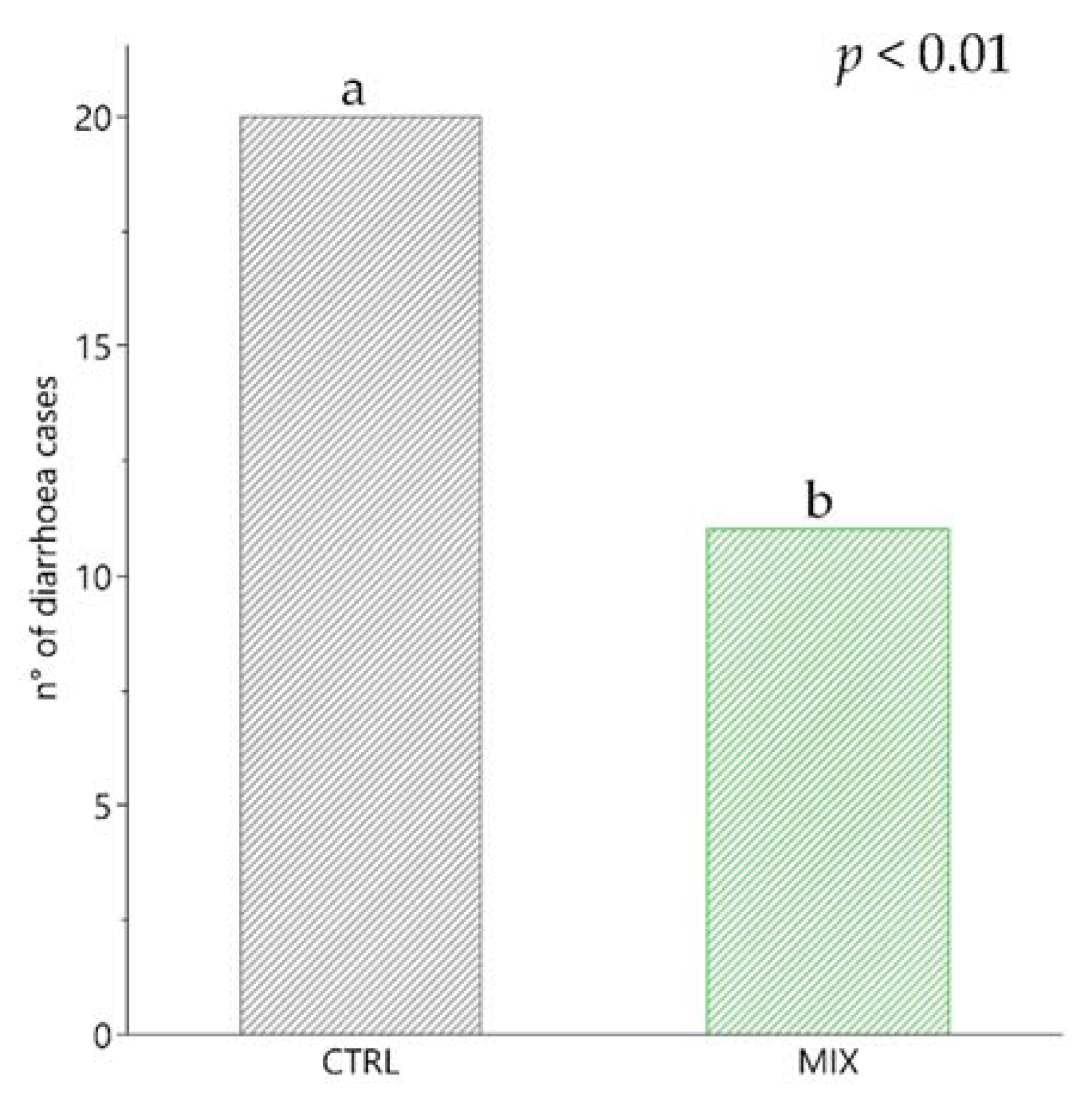
3.3. Diarrhoea Incidence
3.4. Serum Metabolic Profile
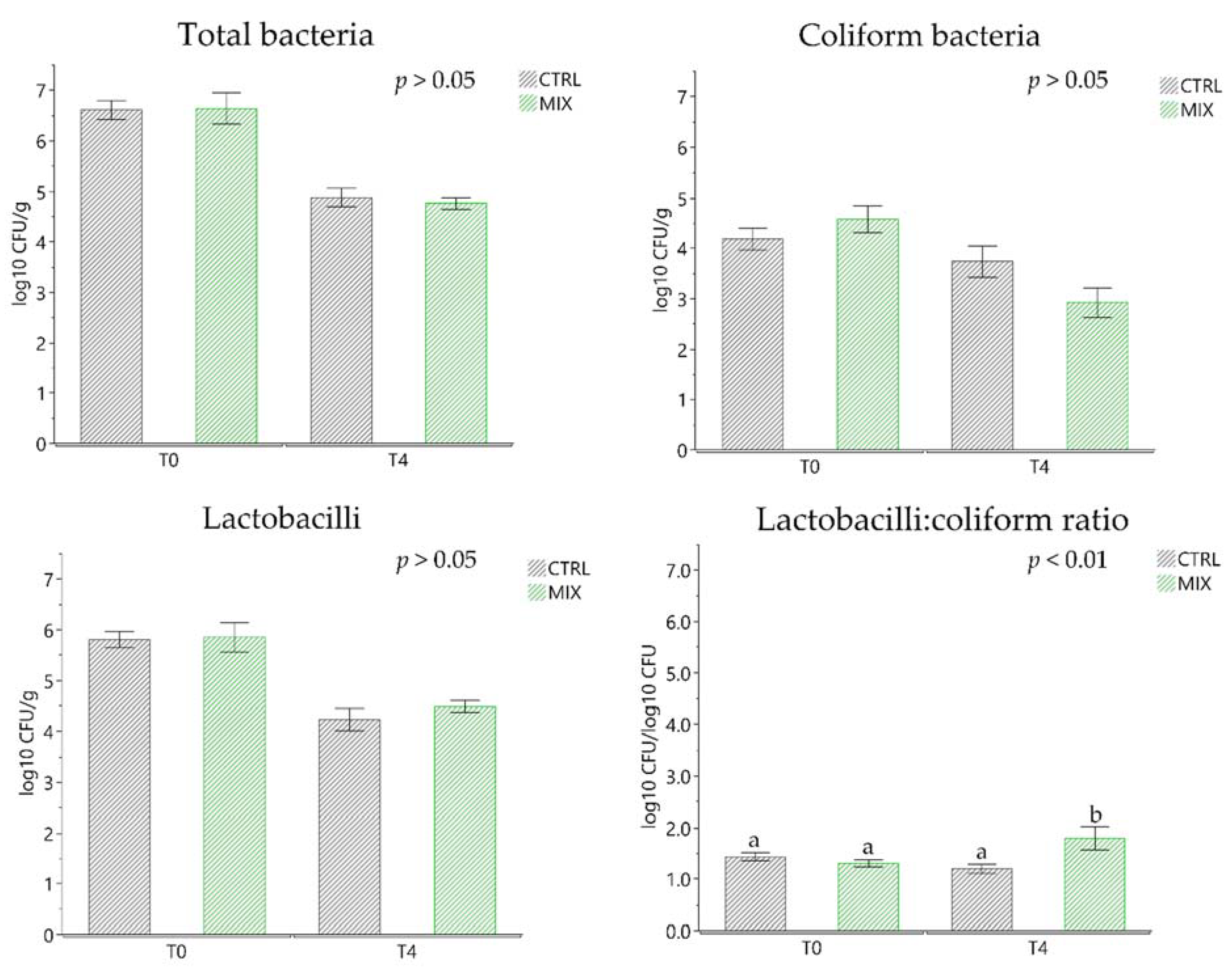
3.5. Microbiological Analysis of Faecal Samples
3.6. Nitrogen Content, Apparent Nitrogen Digestibility, Volatile Fatty Acids and pH of Faecal Samples
3.7. Microbiota Composition and Community Diversity Associated with Mix Supplementation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arsène, M.M.; Davares, A.K.; Andreevna, S.L.; Vladimirovich, E.A.; Carime, B.Z.; Marouf, R.; Khelifi, I. The use of probiotics in animal feeding for safe production and as potential alternatives to antibiotics. Vet. World 2021, 14, 319. [Google Scholar] [CrossRef]
- Ng, W.-J.; Shit, C.-S.; Ee, K.-Y.; Chai, T.-T. Plant Natural Products for Mitigation of Antibiotic Resistance. In Sustainable Agriculture Reviews 49; Springer: Berlin/Heidelberg, Germany, 2021; pp. 57–91. [Google Scholar]
- Tang, K.L.; Caffrey, N.P.; Nóbrega, D.B.; Cork, S.C.; Ronksley, P.E.; Barkema, H.W.; Polachek, A.J.; Ganshorn, H.; Sharma, N.; Kellner, J.D. Restricting the use of antibiotics in food-producing animals and its associations with antibiotic resistance in food-producing animals and human beings: A systematic review and meta-analysis. Lancet Planet. Health 2017, 1, e316–e327. [Google Scholar] [CrossRef]
- Cormican, M.; Hopkins, S.; Jarlier, V.; Reilly, J.; Simonsen, G.; Strauss, R.; Vandenberg, O.; Zabicka, D.; Zarb, P.; Catchpole, M.; et al. ECDC, EFSA and EMA Joint Scientific Opinion on a list of outcome indicators as regards surveillance of antimicrobial resistance and antimicrobial consumption in humans and food-producing animals. EFSA J. 2017, 15. [Google Scholar] [CrossRef]
- European Medicines Agency (EMA). Questions and Answers on Veterinary Medicinal Products Containing Zinc Oxide to be Administered Orally to Food-Producing Species. In Outcome of a Referral Procedure under Article 35 of Directive 2001/82/EC (EMEA/V/A/118); EMA/394961/2017; European Medicines Agency: London, UK, 2017. [Google Scholar]
- Hejna, M.; Onelli, E.; Moscatelli, A.; Bellotto, M.; Cristiani, C.; Stroppa, N.; Rossi, L. Heavy-Metal Phytoremediation from Livestock Wastewater and Exploitation of Exhausted Biomass. Int. J. Environ. Res. Public Health 2021, 18, 2239. [Google Scholar] [CrossRef]
- European Parliament. Regulation No. 1831/2003 of the European Parliament and of the Council of 22 September 2003 on Additives for Use in Animal Nutrition; Official Journal of the European Union: Brussels, Belgium, 2003. [Google Scholar]
- Bonetti, A.; Tugnoli, B.; Piva, A.; Grilli, E. Towards Zero Zinc Oxide: Feeding Strategies to Manage Post-Weaning Diarrhea in Piglets. Animals 2021, 11, 642. [Google Scholar] [CrossRef] [PubMed]
- Hejna, M.; Moscatelli, A.; Onelli, E.; Baldi, A.; Pilu, S.; Rossi, L. Evaluation of concentration of heavy metals in animal rearing system. Ital. J. Anim. Sci. 2019, 18, 1372–1384. [Google Scholar] [CrossRef] [Green Version]
- Lu, C.W.; Wang, S.E.; Wu, W.J.; Su, L.Y.; Wang, C.H.; Wang, P.H.; Wu, C.H. Alternative antibiotic feed additives alleviate pneumonia with inhibiting ACE-2 expression in the respiratory system of piglets. Food Sci. Nutr. 2021, 9, 1112–1120. [Google Scholar] [CrossRef] [PubMed]
- Rossi, L.; Dell’Orto, V.; Vagni, S.; Sala, V.; Reggi, S.; Baldi, A. Protective effect of oral administration of transgenic tobacco seeds against verocytotoxic Escherichia coli strain in piglets. Vet. Res. Commun. 2014, 38, 39–49. [Google Scholar] [CrossRef]
- Chelakkot, C.; Ghim, J.; Ryu, S.H. Mechanisms regulating intestinal barrier integrity and its pathological implications. Exp. Mol. Med. 2018, 50, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Domínguez Díaz, L.; Fernández-Ruiz, V.; Cámara, M. The frontier between nutrition and pharma: The international regulatory framework of functional foods, food supplements and nutraceuticals. Crit. Rev. Food Sci. Nutr. 2020, 60, 1738–1746. [Google Scholar] [CrossRef]
- Lallès, J.-P.; Montoya, C.A. Dietary alternatives to in-feed antibiotics, gut barrier function and inflammation in piglets post-weaning: Where are we now? Anim. Feed Sci. Technol. 2021, 114836. [Google Scholar] [CrossRef]
- Rossi, C.; Compiani, R.; Baldi, G.; Muraro, M.; Marden, J.; Rossi, R.; Pastorelli, G.; Corino, C.; Dell’Orto, V. Organic selenium supplementation improves growth parameters, immune and antioxidant status of newly received beef cattle. J. Anim. Feed Sci. 2017, 26, 100–108. [Google Scholar] [CrossRef] [Green Version]
- Tan, B.; Lim, T.; Boontiam, W. Effect of dietary supplementation with essential oils and a Bacillus probiotic on growth performance, diarrhoea and blood metabolites in weaned pigs. Anim. Prod. Sci. 2020, 61, 64–71. [Google Scholar] [CrossRef]
- Alemayehu, T.A.; Geremew, A.; Getahun, A. The Role of Functional Feed Additives in Tilapia Nutrition. Fish Aquac. J. 2018, 9, g1. [Google Scholar] [CrossRef]
- Bi, Y.; Yang, C.; Diao, Q.; Tu, Y. Effects of dietary supplementation with two alternatives to antibiotics on intestinal microbiota of preweaned calves challenged with Escherichia coli K99. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Dell’Anno, M.; Sotira, S.; Rebucci, R.; Reggi, S.; Castiglioni, B.; Rossi, L. In vitro evaluation of antimicrobial and antioxidant activities of algal extracts. Ital. J. Anim. Sci. 2020, 19, 103–113. [Google Scholar] [CrossRef] [Green Version]
- Caprarulo, V.; Hejna, M.; Giromini, C.; Liu, Y.; Dell’Anno, M.; Sotira, S.; Reggi, S.; Sgoifo-Rossi, C.A.; Callegari, M.L.; Rossi, L. Evaluation of Dietary Administration of Chestnut and Quebracho Tannins on Growth, Serum Metabolites and Fecal Parameters of Weaned Piglets. Animals 2020, 10, 1945. [Google Scholar] [CrossRef] [PubMed]
- Reggi, S.; Giromini, C.; Dell’Anno, M.; Baldi, A.; Rebucci, R.; Rossi, L. In Vitro Digestion of Chestnut and Quebracho Tannin Extracts: Antimicrobial Effect, Antioxidant Capacity and Cytomodulatory Activity in Swine Intestinal IPEC-J2 Cells. Animals 2020, 10, 195. [Google Scholar] [CrossRef] [Green Version]
- Caprarulo, V.; Giromini, C.; Rossi, L. Chestnut and quebracho tannins in pig nutrition: The effects on performance and intestinal health. Animal 2020, 100064. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, Z.; Bach, S.; McAllister, T. Sensitivity of Escherichia coli to seaweed (Ascophyllum nodosum) phlorotannins and terrestrial tannins. Asian Australas. J. Anim. Sci. 2009, 22, 238–245. [Google Scholar] [CrossRef]
- Liu, H.W.; Dong, X.F.; Tong, J.M.; Zhang, Q. A comparative study of growth performance and antioxidant status of rabbits when fed with or without chestnut tannins under high ambient temperature. Anim. Feed Sci. Technol. 2011, 164, 89–95. [Google Scholar] [CrossRef]
- Gancarcikova, S.; Nemcova, R.; Popper, M.; Hrckova, G.; Scirankova, L.; Madar, M.; Mudronova, D.; Vilcek, S.; Zitnan, R. The influence of feed-supplementation with probiotic strain Lactobacillus reuteri CCM 8617 and alginite on intestinal microenvironment of SPF mice infected with Salmonella Typhimurium CCM 7205. Probiotics Antimicrob. Proteins 2019, 11, 493–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Expert Group for Technical Advice on Organic Production (EGTOP). Final Report on Feed; EGTOP/1/2011; Directorate-General for Agriculture and Rural Development: Brussels, Belgium, 2011. [Google Scholar]
- Dell’Anno, M.; Hejna, M.; Sotira, S.; Caprarulo, V.; Reggi, S.; Pilu, R.; Miragoli, F.; Callegari, M.L.; Panseri, S.; Rossi, L. Evaluation of leonardite as a feed additive on lipid metabolism and growth of weaned piglets. Anim. Feed Sci. Technol. 2020, 266, 114519. [Google Scholar] [CrossRef]
- Trckova, M.; Lorencova, A.; Babak, V.; Neca, J.; Ciganek, M. The effect of leonardite and lignite on the health of weaned piglets. Res. Vet. Sci. 2018, 119, 134–142. [Google Scholar] [CrossRef]
- Miragoli, F.; Patrone, V.; Prandini, A.; Sigolo, S.; Dell’Anno, M.; Rossi, L.; Senizza, A.; Morelli, L.; Callegari, M.L. Implications of Tributyrin on Gut Microbiota Shifts Related to Performances of Weaning Piglets. Microorganisms 2021, 9, 584. [Google Scholar] [CrossRef]
- Sotira, S.; Dell’Anno, M.; Caprarulo, V.; Hejna, M.; Pirrone, F.; Callegari, M.L.; Tucci, T.V.; Rossi, L. Effects of Tributyrin Supplementation on Growth Performance, Insulin, Blood Metabolites and Gut Microbiota in Weaned Piglets. Animals 2020, 10, 726. [Google Scholar] [CrossRef]
- Murray, R.L.; Zhang, W.; Iwaniuk, M.; Grilli, E.; Stahl, C.H. Dietary tributyrin, an HDAC inhibitor, promotes muscle growth through enhanced terminal differentiation of satellite cells. Physiol. Rep. 2018, 6, e13706. [Google Scholar] [CrossRef]
- Zhang, W.-X.; Zhang, Y.; Zhang, X.-W.; Deng, Z.-X.; Liu, J.-X.; He, M.-L.; Wang, H.-F. Effects of Dietary Supplementation with Combination of Tributyrin and Essential Oil on Gut Health and Microbiota of Weaned Piglets. Animals 2020, 10, 180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panda, A.; Rao, S.; Raju, M.; Sunder, G.S. Effect of butyric acid on performance, gastrointestinal tract health and carcass characteristics in broiler chickens. Asian Australas. J. Anim. Sci. 2009, 22, 1026–1031. [Google Scholar] [CrossRef]
- European Commission. Amending Annex I to Directive 2002/32/EC of the European Parliament and of the Council as Regards Mercury, Free Gossypol, Nitrites and Mowrah, Bassia, Madhuca; Commission Directive EU 6/2010 (Text with EEA Relevance); Official Journal of the European Union: Brussels, Belgium, 2010. [Google Scholar]
- National Research Council (NRC). Nutrient Requirements of Swine, 11th Revised ed.; Animal Nutrition Series; National Research Council: Washington, DC, USA; The National Academies Press: Washington, DC, USA, 2012.
- American Oil Chemistry Society (AOCS). Crude Fiber Analysis in Feeds by Filter Bag Technique. In Official Methods and Recommended Practices, 4th ed.; AOCS: Champaign, IL, USA, 2009. [Google Scholar]
- Warke, R.; Kamat, A.; Kamat, M.; Thomas, P. Incidence of pathogenic psychrotrophs in ice creams sold in some retail outlets in Mumbai, India. Food Control 2000, 11, 77–83. [Google Scholar] [CrossRef]
- Henning, C.; Vijayakumar, P.; Adhikari, R.; Jagannathan, B.; Gautam, D.; Muriana, P.M. Isolation and taxonomic identity of bacteriocin-producing lactic acid bacteria from retail foods and animal sources. Microorganisms 2015, 3, 80–93. [Google Scholar] [CrossRef] [Green Version]
- Dowell, V.R., Jr.; Hawkins, T.M. Laboratory Methods in Anaerobic Bacteriology—CDC Laboratory Manual; Center for Disease Control: Atlanta, GA, USA, 1974.
- Sales, J.; Janssens, G. Acid-insoluble ash as a marker in digestibility studies: A review. J. Anim. Feed Sci. 2003, 12, 383–401. [Google Scholar] [CrossRef]
- Ahmed, S.; Minuti, A.; Bani, P. In vitro rumen fermentation characteristics of some naturally occurring and synthetic sugars. Ital. J. Anim. Sci. 2013, 12, e57. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Chong, J.; Liu, P.; Zhou, G.; Xia, J. Using MicrobiomeAnalyst for comprehensive statistical, functional, and meta-analysis of microbiome data. Nat. Protoc. 2020, 15, 799–821. [Google Scholar] [CrossRef] [PubMed]
- Dhariwal, A.; Chong, J.; Habib, S.; King, I.L.; Agellon, L.B.; Xia, J. MicrobiomeAnalyst: A web-based tool for comprehensive statistical, visual and meta-analysis of microbiome data. Nucleic Acids Res. 2017, 45, W180–W188. [Google Scholar] [CrossRef] [PubMed]
- European Commission. Commission Regulation (EC) No 429/2008, on Detailed Rules for the Implementation of Regulation (EC) No 1831/2003 of the European Parliament and of the Council as Regards the Preparation and the Presentation of Applications and the Assessment and the Authorisation of Feed Additives; Regulation EC 429/2008 (Text with EEA Relevance); Official Journal of the European Union: Brussels, Belgium, 2008. [Google Scholar]
- Rychen, G.; Aquilina, G.; Azimonti, G.; Bampidis, V.; Bastos, M.D.L.; Bories, G.; Chesson, A.; Cocconcelli, P.S.; Flachowsky, G. EFSA Panel on Additives and Products or Substances used in Animal Feed: Guidance on the assessment of the efficacy of feed additives. EFSA J. 2018, 16, e05274. [Google Scholar] [PubMed] [Green Version]
- Girard, M.; Thanner, S.; Pradervand, N.; Hu, D.; Ollagnier, C.; Bee, G. Hydrolysable chestnut tannins for reduction of postweaning diarrhea: Efficacy on an experimental ETEC F4 model. PLoS ONE 2018, 13, e0197878. [Google Scholar] [CrossRef]
- Hou, Y.; Wang, L.; Yi, D.; Ding, B.; Chen, X.; Wang, Q.; Zhu, H.; Liu, Y.; Yin, Y.; Gong, J.; et al. Dietary supplementation with tributyrin alleviates intestinal injury in piglets challenged with intrarectal administration of acetic acid. Br. J. Nutr. 2014, 111, 1748–1758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynegaard, J.; Kjeldsen, N.; Bache, J.; Weber, N.; Hansen, C.; Nielsen, J.; Amdi, C. Low protein diets without medicinal zinc oxide for weaned pigs reduced diarrhoea treatments and average daily gain. Animal 2021, 15, 100075. [Google Scholar] [CrossRef]
- Laine, T.M.; Lyytikäinen, T.; Yliaho, M.; Anttila, M. Risk factors for post-weaning diarrhoea on piglet producing farms in Finland. Acta Vet. Scand. 2008, 50, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Girard, M.; Hu, D.; Pradervand, N.; Neuenschwander, S.; Bee, G. Chestnut extract but not sodium salicylate decreases the severity of diarrhea and enterotoxigenic Escherichia coli F4 shedding in artificially infected piglets. PLoS ONE 2020, 15, e0214267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Girard, M.; Bee, G. Invited review: Tannins as a potential alternative to antibiotics to prevent coliform diarrhea in weaned pigs. Animal 2020, 14, 95–107. [Google Scholar] [CrossRef] [Green Version]
- Islam, K.M.S.; Schuhmacher, A.; Gropp, J.M. Humic Acid Substances in Animal Agriculture. Pak. J. Nutr. 2005, 4, 126–134. [Google Scholar]
- Dong, L.; Zhong, X.; He, J.; Zhang, L.; Bai, K.; Xu, W.; Wang, T.; Huang, X. Supplementation of tributyrin improves the growth and intestinal digestive and barrier functions in intrauterine growth-restricted piglets. Clin. Nutr. 2016, 35, 399–407. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Hu, J.; Mahfuz, S.; Piao, X. Effects of hydrolysable tannins as zinc oxide substitutes on antioxidant status, immune function, intestinal morphology, and digestive enzyme activities in weaned piglets. Animals 2020, 10, 757. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Song, Y.; Yu, B.; He, J.; Zheng, P.; Mao, X.; Huang, Z.; Luo, Y.; Luo, J.; Yan, H. Tannic acid prevents post-weaning diarrhea by improving intestinal barrier integrity and function in weaned piglets. J. Anim. Sci. Biotechnol. 2020, 11, 1–11. [Google Scholar] [CrossRef]
- Wang, C.; Shen, Z.; Cao, S.; Zhang, Q.; Peng, Y.; Hong, Q.; Feng, J.; Hu, C. Effects of tributyrin on growth performance, intestinal microflora and barrier function of weaned pigs. Anim. Feed Sci. Technol. 2019, 258, 114311. [Google Scholar] [CrossRef]
- Liu, Y.; Espinosa, C.; Abelilla, J.; Casas, G.; Lagos, L.; Lee, S.; Kwon, W.; Mathai, J.; Navarro, D.; Jaworski, N.; et al. Non-antibiotic feed additives in diets for pigs: A review. Anim. Nutr. 2018, 4, 113–125. [Google Scholar] [CrossRef]
- Klem, T.B.; Bleken, E.; Morberg, H.; Thoresen, S.I.; Framstad, T. Hematologic and biochemical reference intervals for Norwegian crossbreed grower pigs. Vet. Clin. Pathol. 2010, 39, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Friendship, R.M.; Lumsden, J.H.; McMillan, I.; Wilson, M.R. Hematology and Biochemistry Reference Values for Ontario Swine. Can. J. Comp. Med. 1984, 48, 390–393. [Google Scholar]
- Izsler. Parametri di Chimica Clinica: Valori Osservati in Suini di Diversa Età. Available online: https://www.izsler.it/pls/izs_bs/v3_s2ew_consultazione.mostra_pagina?id_pagina=1494 (accessed on 18 March 2021).
- Ference, B.A.; Graham, I.; Tokgozoglu, L.; Catapano, A.L. Impact of lipids on cardiovascular health: JACC health promotion series. J. Am. Coll. Cardiol. 2018, 72, 1141–1156. [Google Scholar] [CrossRef] [PubMed]
- Buscemi, S.; Corleo, D.; Buscemi, C.; Randazzo, C.; Borzì, A.M.; Barile, A.M.; Rosafio, G.; Ciaccio, M.; Caldarella, R.; Meli, F. Influence of Habitual Dairy Food Intake on LDL Cholesterol in a Population-Based Cohort. Nutrients 2021, 13, 593. [Google Scholar] [CrossRef] [PubMed]
- Fechner, A.; Kiehntopf, M.; Jahreis, G. The Formation of Short-Chain Fatty Acids Is Positively Associated with the Blood Lipid–Lowering Effect of Lupin Kernel Fiber in Moderately Hypercholesterolemic Adults. J. Nutr. 2014, 144, 599–607. [Google Scholar] [CrossRef]
- He, J.; Dong, L.; Xu, W.; Bai, K.; Lu, C.; Wu, Y.; Huang, Q.; Zhang, L.; Wang, T. Dietary tributyrin supplementation attenuates insulin resistance and abnormal lipid metabolism in suckling piglets with intrauterine growth retardation. PLoS ONE 2015, 10, e0136848. [Google Scholar]
- Galassi, G.; Mason, F.; Rapetti, L.; Crovetto, G.M.; Spanghero, M. Digestibility and metabolic utilisation of diets containing chestnut tannins and their effects on growth and slaughter traits of heavy pigs. Ital. J. Anim. Sci. 2019, 18, 746–753. [Google Scholar] [CrossRef] [Green Version]
- Sahakyan, N.; Bartoszek, A.; Jacob, C.; Petrosyan, M.; Trchounian, A. Bioavailability of Tannins and Other Oligomeric Polyphenols: A Still to Be Studied Phenomenon. Curr. Pharmacol. Rep. 2020, 6, 131–136. [Google Scholar] [CrossRef]
- Kim, Y.; Lee, J.H.; Kim, T.H.; Song, M.H.; Yun, W.; Oh, H.J.; Lee, J.S.; Kim, H.B.; Cho, J.H. Effects of low protein diets added with protease on growth performance, nutrient digestibility of weaned piglets and growing-finishing pigs. J. Anim. Sci. Technol. 2021. [Google Scholar] [CrossRef]
- Pieper, R.; Janczyk, P.; Schumann, R.; Souffrant, W. The intestinal microflora of piglets around weaning with emphasis on lactobacilli. Arch. Zootech. 2006, 9, 28–40. [Google Scholar]
- Castillo, M.; Martín-Orúe, S.M.; Manzanilla, E.G.; Badiola, I.; Martín, M.; Gasa, J. Quantification of total bacteria, enterobacteria and lactobacilli populations in pig digesta by real-time PCR. Vet. Microbiol. 2006, 114, 165–170. [Google Scholar] [CrossRef]
- Wang, M.; Huang, H.; Hu, Y.; Huang, J.; Yang, H.; Wang, L.; Chen, S.; Chen, C.; He, S. Effects of dietary microencapsulated tannic acid supplementation on the growth performance, intestinal morphology, and intestinal microbiota in weaning piglets. J. Anim. Sci. 2020, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maes, D.; Chantziaras, I.; Vallaey, E.; Demeyere, K.; Meyer, E.; Janssens, G.P. Faecal pH throughout the reproductive cycle of sows in commercial pig herds. J. Anim. Physiol. Anim. Nutr. 2020. [Google Scholar] [CrossRef] [PubMed]
- Dehority, B.A. Microbial ecology of cell wall fermentation. Forage Cell Wall Struct. Dig. 1993, 425–453. [Google Scholar] [CrossRef]
- Schautteet, K.; Vanrompay, D. Chlamydiaceae infections in pig. Vet. Res. 2011, 42, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Rogers, D.G.; Andersen, A.A. Intestinal lesions caused by a strain of Chlamydia suis in weanling pigs infected at 21 days of age. J. Vet. Diagn. Investig. 2000, 12, 233–239. [Google Scholar] [CrossRef] [Green Version]
- Nietfeld, J.C.; Janke, B.H.; Leslie-Steen, P.; Robison, D.J.; Zeman, D.H. Small intestinal Chlamydia infection in piglets. J. Vet. Diagn. Investig. 1993, 5, 114–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clavel, T.; Lepage, P.; Charrier, C. The family coriobacteriaceae. Prokaryotes 2014, 11, 201–238. [Google Scholar]
- Gill, S.R.; Pop, M.; Deboy, R.T.; Eckburg, P.B.; Turnbaugh, P.J.; Samuel, B.S.; Gordon, J.I.; Relman, D.A.; Fraser-Liggett, C.M.; Nelson, K.E. Metagenomic analysis of the human distal gut microbiome. Science 2006, 312, 1355–1359. [Google Scholar] [CrossRef] [Green Version]
- Ran, S.; Mu, C.; Zhu, W. Diversity and community pattern of sulfate-reducing bacteria in piglet gut. J. Anim. Sci. Biotechnol. 2019, 10, 1–11. [Google Scholar] [CrossRef]
- Unno, T.; Choi, J.-H.; Hur, H.-G.; Sadowsky, M.J.; Ahn, Y.-T.; Huh, C.-S.; Kim, G.-B.; Cha, C.-J. Changes in human gut microbiota influenced by probiotic fermented milk ingestion. J. Dairy Sci. 2015, 98, 3568–3576. [Google Scholar] [CrossRef] [Green Version]
- Yang, Q.; Huang, X.; Zhao, S.; Sun, W.; Yan, Z.; Wang, P.; Li, S.; Huang, W.; Zhang, S.; Liu, L. Structure and function of the fecal microbiota in diarrheic neonatal piglets. Front. Microbiol. 2017, 8, 502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amat, S.; Lantz, H.; Munyaka, P.M.; Willing, B.P. Prevotella in Pigs: The Positive and Negative Associations with Production and Health. Microorganisms 2020, 8, 1584. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Tsai, T.; Deng, F.; Wei, X.; Chai, J.; Knapp, J.; Apple, J.; Maxwell, C.V.; Lee, J.A.; Li, Y. Longitudinal investigation of the swine gut microbiome from birth to market reveals stage and growth performance associated bacteria. Microbiome 2019, 7, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Du, L.; Li, X.; Zhong, H.; Ding, Y.; Liu, Z.; Ge, L. Identification of the core bacteria in rectums of diarrheic and non-diarrheic piglets. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients, % as Fed Basis | CTRL | MIX |
|---|---|---|
| Barley, meal | 26.84 | 26.84 |
| Wheat, meal | 12.45 | 10.45 |
| Corn, flakes | 11.63 | 11.63 |
| Corn, meal | 10.00 | 10.00 |
| Barley, flakes | 7.50 | 7.50 |
| Soy protein concentrates | 5.00 | 5.00 |
| Biscuits, meal | 4.00 | 4.00 |
| Soybean, meal (44%) | 4.00 | 4.00 |
| Dextrose monohydrate | 3.50 | 3.50 |
| Sweet milk whey | 2.50 | 2.50 |
| Herring, meal | 2.00 | 2.00 |
| Plasma, meal | 2.00 | 2.00 |
| Beet pulp | 1.40 | 1.40 |
| Acidifiers 1 | 1.70 | 1.70 |
| Coconut oil | 1.00 | 1.00 |
| Soy oil | 1.00 | 1.00 |
| Dicalcium phosphate | 0.60 | 0.60 |
| L-Lysine | 0.60 | 0.60 |
| Benzoic acid | 0.50 | 0.50 |
| Vitamins and mineral premix 2 | 0.50 | 0.50 |
| DL-Methionine | 0.39 | 0.39 |
| L-Threonine | 0.35 | 0.35 |
| Sodium Chloride | 0.27 | 0.27 |
| L-Valine (96.5%) | 0.12 | 0.12 |
| Enzyme mix 3 | 0.10 | 0.10 |
| L-Tryptophan | 0.05 | 0.05 |
| Experimental mix 4 | - | 2.00 |
| Calculated Chemical Composition 5 | ||
| Crude protein (%) | 18.65 | 18.33 |
| Fat (%) | 4.78 | 4.75 |
| Crude fibre (%) | 3.00 | 2.98 |
| Ashes (%) | 5.52 | 5.48 |
| DE 6 (Mc/Kg) | 3.92 | 3.83 |
| Serum Metabolite | CTRL | MIX | SE CTRL | SE MIX | p-Value |
|---|---|---|---|---|---|
| Total protein content, g/L | 54.92 a | 47.75 b | 1.46 | 1.67 | 0.0258 |
| Albumin, g/L | 27.89 | 25.91 | 1.24 | 1.42 | 0.3808 |
| Globulin, g/L | 25.85 | 23.31 | 1.44 | 1.62 | 0.2938 |
| Albumin/Globulin (A/G) | 1.06 | 1.18 | 0.06 | 0.07 | 0.2465 |
| Urea, mmol/L | 0.99 | 0.78 | 0.06 | 0.07 | 0.0746 |
| Alanine aminotransferase (ALT-GPT), IU/L | 54.75 | 56.81 | 4.68 | 5.27 | 0.7867 |
| Total bilirubin, µmol/L | 1.90 | 1.80 | 0.11 | 0.13 | 0.5750 |
| Glucose, mmol/L | 6.19 | 6.91 | 0.48 | 0.55 | 0.4169 |
| Phosphorus, mmol/L | 3.26 | 3.10 | 0.13 | 0.15 | 0.4547 |
| Magnesium, mmol/L | 0.92 | 0.88 | 0.04 | 0.05 | 0.6328 |
| Creatinine, µmol/L | 70.18 | 65.27 | 3.81 | 4.47 | 0.5098 |
| Total cholesterol, mmol/L | 2.62 | 2.36 | 0.12 | 0.13 | 0.1901 |
| High density lipoprotein (HDL), mmol/L | 1.01 | 1.04 | 0.07 | 0.08 | 0.7955 |
| Low density lipoprotein (LDL), mmol/L | 1.48 a | 1.21 b | 0.07 | 0.08 | 0.0435 |
| Triglycerides, mmol/L | 0.60 | 0.69 | 0.09 | 0.10 | 0.4982 |
| Insulin, mU/L | 10.78 | 9.47 | 2.57 | 2.87 | 0.7437 |
| Glucagon, pg/mL | 294.10 | 286.51 | 13.99 | 22.38 | 0.8103 |
| Phylum | log2FC | Log CPM | p-Values | FDR |
|---|---|---|---|---|
| Bacteroidetes | 1.973 | 17.166 | <0.0001 | <0.0001 |
| Fibrobacteres | 2.736 | 11.433 | <0.0001 | 0.0002 |
| Chlamydiae | −2.860 | 10.731 | 0.0058 | 0.0192 |
| Actinobacteria | −1.449 | 6.976 | 0.0101 | 0.0238 |
| Firmicutes | 1.119 | 20.203 | 0.0119 | 0.0238 |
| Cyanobacteria | −1.566 | 7.227 | 0.0274 | 0.0456 |
| Verrucomicrobia | −2.050 | 11.582 | 0.0356 | 0.0509 |
| Family | log2FC | Log CPM | p-Values | FDR |
|---|---|---|---|---|
| Prevotellaceae | 2.010 | 16.721 | <0.0001 | <0.0001 |
| Fibrobacteraceae | 2.359 | 11.288 | 0.0004 | 0.0053 |
| Chlamydiaceae | −3.149 | 10.902 | 0.0035 | 0.0326 |
| Genus | ||||
| Prevotella | 1.963 | 16.848 | <0.0001 | 0.0003 |
| RFN20 | −2.295 | 7.274 | <0.0001 | 0.0003 |
| Eubacterium | −4.652 | 14.923 | <0.0001 | 0.0003 |
| Fibrobacter | 2.396 | 11.328 | 0.0006 | 0.0047 |
| Lachnospira | −2.050 | 12.780 | 0.0032 | 0.0214 |
| Desulfovibrio | −2.301 | 8.221 | 0.0081 | 0.0443 |
| Chlamydia | −2.701 | 10.827 | 0.0107 | 0.0503 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dell’Anno, M.; Reggi, S.; Caprarulo, V.; Hejna, M.; Sgoifo Rossi, C.A.; Callegari, M.L.; Baldi, A.; Rossi, L. Evaluation of Tannin Extracts, Leonardite and Tributyrin Supplementation on Diarrhoea Incidence and Gut Microbiota of Weaned Piglets. Animals 2021, 11, 1693. https://doi.org/10.3390/ani11061693
Dell’Anno M, Reggi S, Caprarulo V, Hejna M, Sgoifo Rossi CA, Callegari ML, Baldi A, Rossi L. Evaluation of Tannin Extracts, Leonardite and Tributyrin Supplementation on Diarrhoea Incidence and Gut Microbiota of Weaned Piglets. Animals. 2021; 11(6):1693. https://doi.org/10.3390/ani11061693
Chicago/Turabian StyleDell’Anno, Matteo, Serena Reggi, Valentina Caprarulo, Monika Hejna, Carlo Angelo Sgoifo Rossi, Maria Luisa Callegari, Antonella Baldi, and Luciana Rossi. 2021. "Evaluation of Tannin Extracts, Leonardite and Tributyrin Supplementation on Diarrhoea Incidence and Gut Microbiota of Weaned Piglets" Animals 11, no. 6: 1693. https://doi.org/10.3390/ani11061693