
S. aureus Biofilm Protein Expression Linked to Antimicrobial Resistance: A Proteomic Study

 , , , , ,
, , , , , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. Proteomic Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Oniciuc, E.-A.; Likotrafiti, E.; Alvarez-Molina, A.; Prieto, M.; López, M.; Alvarez-Ordóñez, A. Food processing as a risk factor for antimicrobial resistance spread along the food chain. Curr. Opin. Food Sci. 2019, 30, 21–26. [Google Scholar] [CrossRef]
- Guardabassi, L.; Larsen, J.; Weese, J.; Butaye, P.; Battisti, A.; Kluytmans, J.; Lloyd, D.; Skov, R. Public health impact and antimicrobial selection of meticillin-resistant staphylococci in animals. J. Glob. Antimicrob. Resist. 2013, 1, 55–62. [Google Scholar] [CrossRef]
- Weese, J.S.; van Duijkeren, E. Methicillin-resistant Staphylococcus aureus and Staphylococcus pseudintermedius in veterinary medicine. Vet. Microbiol. 2010, 140, 418–429. [Google Scholar] [CrossRef]
- Herrera, J.J.R.; Cabo, M.L.; González, A.; Pazos, I.; Pastoriza, L. Adhesion and detachment kinetics of several strains of Staphylococcus aureus subsp. aureus under three different experimental conditions. Food Microbiol. 2007, 24, 585–591. [Google Scholar] [CrossRef]
- Donlan, R.M.; Costerton, J.W. Biofilms: Survival mechanisms of clinically relevant microorganisms. Clin. Microbiol. Rev. 2002, 15, 167–193. [Google Scholar] [CrossRef] [Green Version]
- Sauer, K.; Camper, A.K.; Ehrlich, G.D.; Costerton, J.W.; Davies, D.G. Pseudomonas aeruginosa displays multiple phenotypes during development as a biofilm. J. Bacteriol. 2002, 184, 1140–1154. [Google Scholar] [CrossRef] [Green Version]
- Resch, A.; Rosenstein, R.; Nerz, C.; Götz, F. Differential gene expression profiling of Staphylococcus aureus cultivated under biofilm and planktonic conditions. Appl. Environ. Microbiol. 2005, 71, 2663–2676. [Google Scholar] [CrossRef] [Green Version]
- Costerton, J.W.; Stewart, P.S.; Greenberg, E.P. Bacterial biofilms: A common cause of persistent infections. Science 1999, 284, 1318–1322. [Google Scholar] [CrossRef] [Green Version]
- Brooks, J.D.; Flint, S.H. Biofilms in the food industry: Problems and potential solutions. Int. J. Food Sci. Technol. 2008, 43, 2163–2176. [Google Scholar] [CrossRef]
- Srey, S.; Jahid, I.K.; Ha, S.-D. Biofilm formation in food industries: A food safety concern. Food Control 2013, 31, 572–585. [Google Scholar] [CrossRef]
- Wassenaar, T.; Ussery, D.; Nielsen, L.; Ingmer, H. Review and phylogenetic analysis of qac genes that reduce susceptibility to quaternary ammonium compounds in Staphylococcus species. Eur. J. Microbiol. Immunol. 2015, 5, 44–61. [Google Scholar] [CrossRef] [Green Version]
- Glinel, K.; Thebault, P.; Humblot, V.; Pradier, C.-M.; Jouenne, T. Antibacterial surfaces developed from bio-inspired approaches. Acta Biomater. 2012, 8, 1670–1684. [Google Scholar] [CrossRef]
- Simoes, M.; Simões, L.C.; Vieira, M.J. A review of current and emergent biofilm control strategies. LWT Food Sci. Technol. 2010, 43, 573–583. [Google Scholar] [CrossRef] [Green Version]
- Mah, T.-F.C.; O’Toole, G.A. Mechanisms of biofilm resistance to antimicrobial agents. Trends Microbiol. 2001, 9, 34–39. [Google Scholar] [CrossRef]
- Branda, S.S.; Vik, Å.; Friedman, L.; Kolter, R. Biofilms: The matrix revisited. Trends Microbiol. 2005, 13, 20–26. [Google Scholar] [CrossRef]
- Flemming, H.-C.; Wingender, H.-C.F.J.; Szewzyk, U.; Steinberg, P.; Rice, S.A.; Kjelleberg, S.A.R.S. Biofilms: An emergent form of bacterial life. Nat. Rev. Microbiol. 2016, 14, 563. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Vishakha, K.; Banerjee, S.; Mondal, S.; Ganguli, A. Sodium alginate-based edible coating containing nanoemulsion of Citrus sinensis essential oil eradicates planktonic and sessile cells of food-borne pathogens and increased quality attributes of tomatoes. Int. J. Biol. Macromol. 2020, 162, 1770–1779. [Google Scholar] [CrossRef] [PubMed]
- Atshan, S.S.; Shamsudin, M.N.; Esekawi, Z.; Lung, L.T.T.; Talab, F.B.; Liew, Y.K.; Alreshidi, M.A.; Abduljaleel, S.A.; Hamat, R.A. Comparative proteomic analysis of extracellular proteins expressed by various clonal types of Staphylococcus aureus and during planktonic growth and biofilm development. Front. Microbiol. 2015, 6, 524. [Google Scholar] [CrossRef]
- Bénard, L.; Litzler, P.Y.; Cosette, P.; Lemeland, J.F.; Jouenne, T.; Junter, G.A. Proteomic analysis of Staphylococcus aureus biofilms grown in vitro on mechanical heart valve leaflets. J. Biomed. Mater. Res. Part A An Off. J. Soc. Biomater. Japanese Soc. Biomater. Aust. Soc. Biomater. Korean Soc. Biomater. 2009, 88, 1069–1078. [Google Scholar]
- Resch, A.; Leicht, S.; Saric, M.; Pásztor, L.; Jakob, A.; Götz, F.; Nordheim, A. Comparative proteome analysis of Staphylococcus aureus biofilm and planktonic cells and correlation with transcriptome profiling. Proteomics 2006, 6, 1867–1877. [Google Scholar] [CrossRef]
- Di Ciccio, P.; Vergara, A.; Festino, A.; Paludi, D.; Zanardi, E.; Ghidini, S.; Ianieri, A. Biofilm formation by Staphylococcus aureus on food contact surfaces: Relationship with temperature and cell surface hydrophobicity. Food Control 2015, 50, 930–936. [Google Scholar] [CrossRef]
- Soggiu, A.; Piras, C.; Greco, V.; Devoto, P.; Urbani, A.; Calzetta, L.; Bortolato, M.; Roncada, P. Exploring the neural mechanisms of finasteride: A proteomic analysis in the nucleus accumbens. Psychoneuroendocrinology 2016, 74, 387–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piras, C.; Soggiu, A.; Bonizzi, L.; Greco, V.; Ricchi, M.; Arrigoni, N.; Bassols, A.; Urbani, A.; Roncada, P. Identification of immunoreactive proteins of Mycobacterium avium subsp. paratuberculosis. Proteomics 2015, 15, 813–823. [Google Scholar] [CrossRef]
- Cramton, S.E.; Gerke, C.; Schnell, N.F.; Nichols, W.W.; Götz, F. The intercellular adhesion (ica) locus is present in Staphylococcus aureus and is required for biofilm formation. Infect. Immun. 1999, 67, 5427–5433. [Google Scholar] [CrossRef] [Green Version]
- Seidl, K.; Goerke, C.; Wolz, C.; Mack, D.; Berger-Bächi, B.; Bischoff, M. Staphylococcus aureus CcpA affects biofilm formation. Infect. Immun. 2008, 76, 2044–2050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziebuhr, W. Staphylococcus aureus and Staphylococcus epidermidis: Emerging pathogens in nosocomial infections. Contrib. Microbiol. 2001, 8, 102–107. [Google Scholar]
- Weber, M.; Liedtke, J.; Plattes, S.; Lipski, A. Bacterial community composition of biofilms in milking machines of two dairy farms assessed by a combination of culture-dependent and–independent methods. PLoS ONE 2019, 14, e0222238. [Google Scholar] [CrossRef]
- Enright, M.C. The evolution of a resistant pathogen–the case of MRSA. Curr. Opin. Pharmacol. 2003, 3, 474–479. [Google Scholar] [CrossRef]
- Nostro, A.; Blanco, A.R.; Cannatelli, M.A.; Enea, V.; Flamini, G.; Morelli, I.; Roccaro, A.S.; Alonzo, V. Susceptibility of methicillin-resistant staphylococci to oregano essential oil, carvacrol and thymol. FEMS Microbiol. Lett. 2004, 230, 191–195. [Google Scholar] [CrossRef] [Green Version]
- Braga, L.C.; Shupp, J.; Cummings, C.; Jett, M.; Takahashi, J.; Carmo, L.; Chartone-Souza, E.; Nascimento, A. Pomegranate extract inhibits Staphylococcus aureus growth and subsequent enterotoxin production. J. Ethnopharmacol. 2005, 96, 335–339. [Google Scholar] [CrossRef]
- Marques, S.C.; Rezende, J.D.G.O.S.; Alves, L.A.D.F.; Silva, B.C.; Alves, E.; De Abreu, L.R.; Píccoli, R.H. Formation of biofilms by Staphylococcus aureus on stainless steel and glass surfaces and its resistance to some selected chemical sanitizers. Braz. J. Microbiol. 2007, 38, 538–543. [Google Scholar] [CrossRef] [Green Version]
- Oulahal, N.; Brice, W.; Martial, A.; Degraeve, P. Quantitative analysis of survival of Staphylococcus aureus or Listeria innocua on two types of surfaces: Polypropylene and stainless steel in contact with three different dairy products. Food Control 2008, 19, 178–185. [Google Scholar] [CrossRef]
- Wolf, C.; Hochgräfe, F.; Kusch, H.; Albrecht, D.; Hecker, M.; Engelmann, S. Proteomic analysis of antioxidant strategies of Staphylococcus aureus: Diverse responses to different oxidants. Proteomics 2008, 8, 3139–3153. [Google Scholar] [CrossRef]
- Engelmann, S.; Hecker, M. Proteomic analysis to investigate regulatory networks in Staphylococcus aureus. In Bacterial Pathogenesis; Springer: Berlin/Heidelberg, Germany, 2008; pp. 25–45. [Google Scholar]
- Sałamaszyńska-Guz, A.; Rose, S.; Lykkebo, C.A.; Taciak, B.; Bącal, P.; Uśpieński, T.; Douthwaite, S. Biofilm formation and motility are promoted by Cj0588-directed methylation of rRNA in Campylobacter jejuni. Front. Cell. Infect. Microbiol. 2018, 7, 533. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Wu, Y.; Lin, Z.; Wang, G.; Jiang, S.; Sun, X.; Tu, H.; Yu, Z.; Qu, D. ClpP participates in stress tolerance, biofilm formation, antimicrobial tolerance, and virulence of Enterococcus faecalis. BMC Microbiol. 2020, 20, 30. [Google Scholar] [CrossRef] [Green Version]
- Lilleorg, S.; Reier, K.; Volõnkin, P.; Remme, J.; Liiv, A. Phenotypic effects of paralogous ribosomal proteins bL31A and bL31B in E. coli. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef]
- Shams, F.; Oldfield, N.J.; Wooldridge, K.G.; Turner, D.P.J. Fructose-1, 6-bisphosphate aldolase (FBA)—A conserved glycolytic enzyme with virulence functions in bacteria: ‘Ill met by moonlight’. Biochem. Soc. Trans. 2014, 42, 1792–1795. [Google Scholar] [CrossRef] [PubMed]
- Oldfield, N.J.; Shams, F.; Wooldridge, K.G.; Turner, D.P.J. Moonlighting Functions of Bacterial Fructose 1, 6-Bisphosphate Aldolases. In Moonlighting Proteins: Novel Virulence Factors in Bacterial Infections; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2017; pp. 321–331. [Google Scholar]
- Ziveri, J.; Tros, F.; Guerrera, I.C.; Chhuon, C.; Audry, M.; Dupuis, M.; Barel, M.; Korniotis, S.; Fillatreau, S.; Gales, L.; et al. The metabolic enzyme fructose-1, 6-bisphosphate aldolase acts as a transcriptional regulator in pathogenic Francisella. Nat. Commun. 2017, 8, 1–15. [Google Scholar] [CrossRef]
- Sadykov, M.R.; Hartmann, T.; Mattes, T.A.; Hiatt, M.; Jann, N.J.; Zhu, Y.; Ledala, N.; Landmann, R.; Herrmann, M.; Rohde, H.; et al. CcpA coordinates central metabolism and biofilm formation in Staphylococcus epidermidis. Microbiology 2011, 157, 3458. [Google Scholar] [CrossRef] [Green Version]
- Rudra, P.; Boyd, J.M. Metabolic control of virulence factor production in Staphylococcus aureus. Curr. Opin. Microbiol. 2020, 55, 81–87. [Google Scholar] [CrossRef]
- Kim, J.N.; Burne, R.A. CcpA and CodY coordinate acetate metabolism in Streptococcus mutans. Appl. Environ. Microbiol. 2017, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuchs, S.; Pané-Farré, J.; Kohler, C.; Hecker, M.; Engelmann, S. Anaerobic gene expression in Staphylococcus aureus. J. Bacteriol. 2007, 189, 4275–4289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becker, P.; Hufnagle, W.; Peters, G.; Herrmann, M. Detection of differential gene expression in biofilm-forming versus planktonic populations of Staphylococcus aureus using micro-representational-difference analysis. Appl. Environ. Microbiol. 2001, 67, 2958–2965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, K.; Yang, X.; Yang, J.; Qiao, X.; Li, F.; Liu, X.; Wei, J.; Wang, L. Alcohol dehydrogenase modulates quorum sensing in biofilm formations of Acinetobacter baumannii. Microb. Pathog. 2020, 148, 104451. [Google Scholar] [CrossRef]
- Shen, F.; Ge, C.; Yuan, P. Metabolomics Study Reveals Inhibition and Metabolic Dysregulation in Staphylococcus aureus Planktonic Cells and Biofilms Induced by Carnosol. Front. Microbiol. 2020, 11, 2315. [Google Scholar] [CrossRef]
- Christensen, B.E.; Trønnes, H.N.; Vollan, K.; Smidsrød, O.; Bakke, R. Biofilm removal by low concentrations of hydrogen peroxide. Biofouling 1990, 2, 165–175. [Google Scholar] [CrossRef]
- Vázquez-Sánchez, D.; Rodríguez-López, P. Biofilm formation of Staphylococcus aureus. In Staphylococcus aureus; Elsevier: Amsterdam, The Netherlands, 2018; pp. 87–103. [Google Scholar]


{kind=link}
| Strains | Source | BPI |
|---|---|---|
| A—S. aureus ATCC n.35556 | ATCC n.35556 | 0.758 |
| B—S. aureus ATCC n.12600 | ATCC n.12600 | 0.405 |
| C—S. aureus n.281 | Food | 1.019 |
| D—S. aureus n.402 | Food-handler | 0.311 |
| E—S. aureus n.184 | Food-handler | 0.290 |
| F—S. aureus ATCC 29213 | ATCC n.29213 | 0.260 |
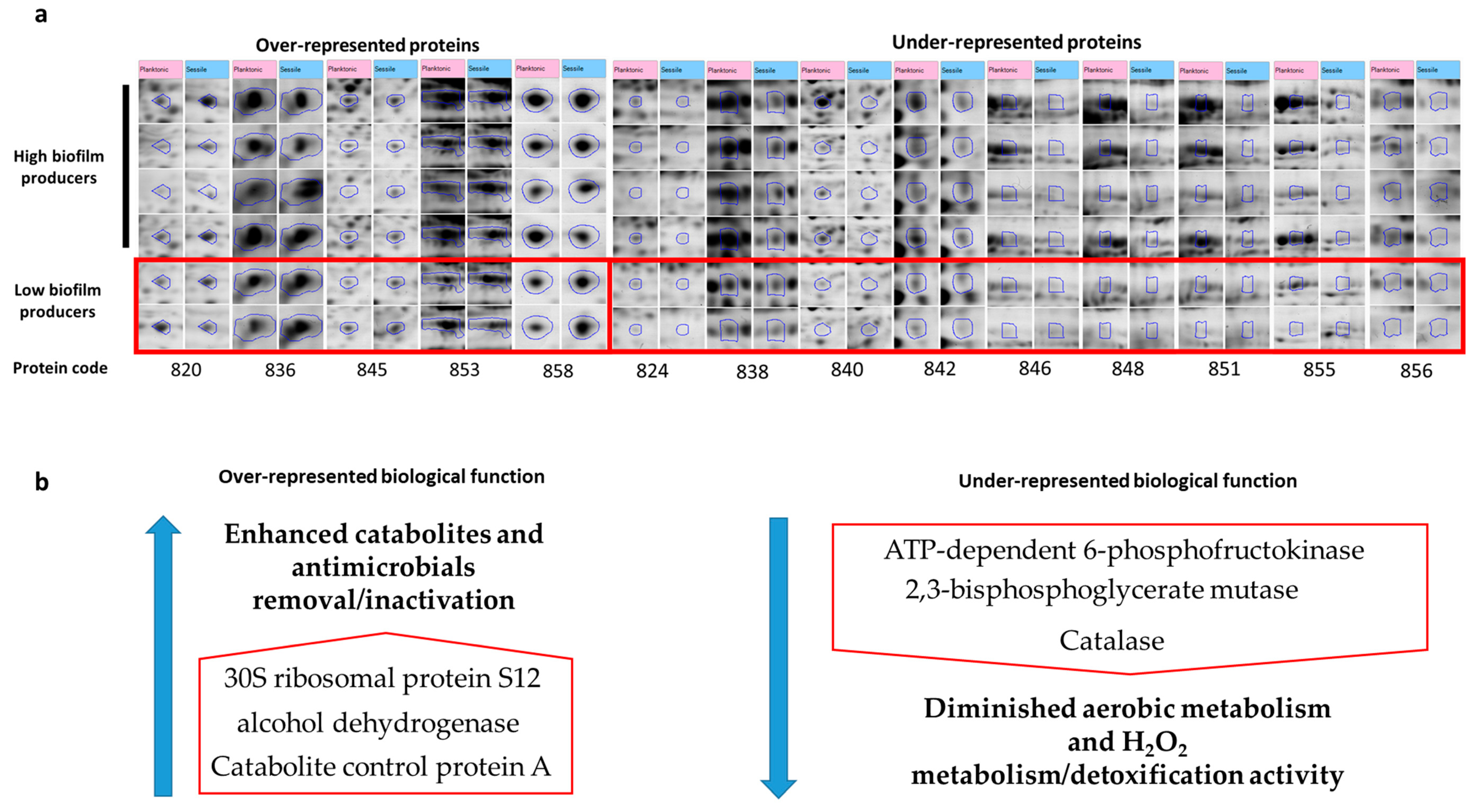
| Uniprot Name/Accession Number | Uniprot Name | SameSpots Coding Number | Protein Name | Mascot Score | Sequence Coverage (%) | N of Matched Peptides | Regulation in Sessile vs. Planktonic | High Biofilm Producers (Sessile vs. Planktonic) |
|---|---|---|---|---|---|---|---|---|
| Q2FJ31 | ADH_STAA3 | 820 | alcohol dehydrogenase | 66 | 35 | 9/59 | ↑0.070 | ↑0.018 |
| A7X656 | GPMA_STAA1 | 824 | 2,3-bisphosphoglycerate-dependent phosphoglycerate mutase OS = Staphylococcus aureus | 116 | 51 | 11/52 | ↓0.001 | ↓0.016 |
| P0A0H0 | RS12_STAA8 | 836 | 30S ribosomal protein S12 | 68 | 37 | 4/45 | ↑0.026 | 0.135 |
| Q2FJN4 | AHPC_STAA3 | 838 | Alkyl hydroperoxide reductase subunit C OS = Staphylococcus aureus | 67 | 39 | 6/31 | ↓0.028 | 0.137 |
| A6U2G5 | PFKA_STAA2 | 840 | ATP-dependent 6-phosphofructokinase OS = Staphylococcus aureus | 121 | 35 | 11/31 | ↓0.084 | ↓0.007 |
| A7WZR9 | CLPP_STAA1 | 842 | ATP-dependent Clp protease proteolytic subunit OS = Staphylococcus aureus | 76 | 30 | 9/23 | ↓0.008 | 0.084 |
| Q5HF38 | CCPA_STAAC | 845 | Catabolite control protein A OS = Staphylococcus aureus | 76 | 26 | 9/35 | ↑0.001 | ↑0.004 |
| Q9L4S1 | CATA_STAAU | 846 | Catalase OS = Staphylococcus aureus GN = katA PE = 3 SV = 1 | 40 | 16 | ↓0.041 | ↓0.041 | |
| Q9L4S1 | CATA_STAAU | 848 | Catalase OS = Staphylococcus aureus GN = katA PE = 3 SV = 1 | 68 | 30 | 6/40 | ↓0.021 | ↓0.027 |
| Q9L4S1 | CATA_STAAU | 851 | Catalase OS = Staphylococcus aureus GN = katA PE = 3 SV = 1 | 98 | 47 | 11/40 | ↓0.012 | ↓0.004 |
| Q2FDQ4 | ALF1_STAA3 | 853 | Fructose-bisphosphate aldolase class 1 OS = Staphylococcus aureus | 112 | 40 | 17/67 | ↑0.096 | ↑0.0002 |
| Q2YSZ4 | GCSPB_STAAB | 855 | Probable glycine dehydrogenase (decarboxylating) subunit 2 OS = Staphylococcus aureus | 68 | 26 | 9/50 | ↓0.037 | ↓0.013 |
| A7X395 | ENGB_STAA1 | 856 | Probable GTP-binding protein EngB OS = Staphylococcus aureus _12042016_ | 68 | 16 | 4/16 | ↓0.007 | 0.060 |
| Q7A551 | Y1532_STAAN | 858 | putative universal stress protein SA153 (782) OS = Staphylococcus aureus | 86 | 36 | 6/35 | ↑0.010 | 0.094 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piras, C.; Di Ciccio, P.A.; Soggiu, A.; Greco, V.; Tilocca, B.; Costanzo, N.; Ceniti, C.; Urbani, A.; Bonizzi, L.; Ianieri, A.; et al. S. aureus Biofilm Protein Expression Linked to Antimicrobial Resistance: A Proteomic Study. Animals 2021, 11, 966. https://doi.org/10.3390/ani11040966
Piras C, Di Ciccio PA, Soggiu A, Greco V, Tilocca B, Costanzo N, Ceniti C, Urbani A, Bonizzi L, Ianieri A, et al. S. aureus Biofilm Protein Expression Linked to Antimicrobial Resistance: A Proteomic Study. Animals. 2021; 11(4):966. https://doi.org/10.3390/ani11040966
Chicago/Turabian StylePiras, Cristian, Pierluigi Aldo Di Ciccio, Alessio Soggiu, Viviana Greco, Bruno Tilocca, Nicola Costanzo, Carlotta Ceniti, Andrea Urbani, Luigi Bonizzi, Adriana Ianieri, and et al. 2021. "S. aureus Biofilm Protein Expression Linked to Antimicrobial Resistance: A Proteomic Study" Animals 11, no. 4: 966. https://doi.org/10.3390/ani11040966