
Prevalence and Antibiotic Resistance Characteristics of Extraintestinal Pathogenic Escherichia coli among Healthy Chickens from Farms and Live Poultry Markets in China

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling and Bacterial Isolation
2.2. Antimicrobial Susceptibility Tests
2.3. Detection of Antibiotic Resistance Genes
2.4. Detection and Serotyping of ExPEC Isolates
2.5. Pulse-Field Gel Electrophoresis (PFGE) and Multilocus Sequence Typing (MLST)
2.6. Statistical Analysis
3. Results
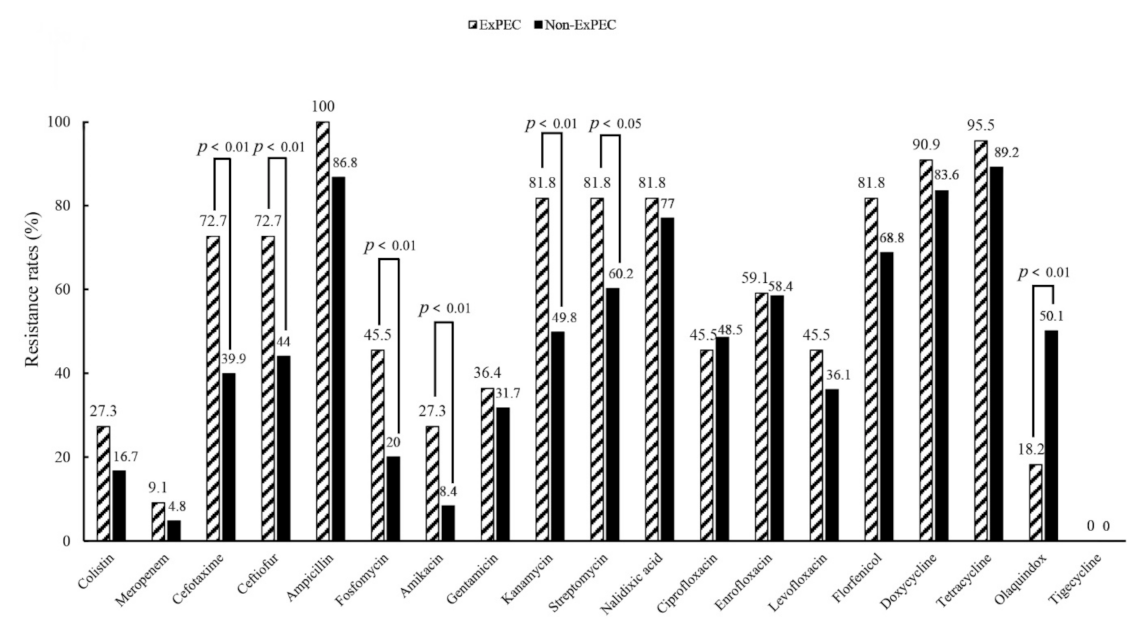
3.1. Antimicrobial Susceptibilities
3.2. Detection of Resistance Genes
3.3. Prevalence of ExPEC Isolates and Their Serogroups
3.4. Antimicrobial Resistance Phenotypes and Genotypes of the ExPEC Isolates
3.5. Genetic Relationships of the ExPEC Isolates
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Mellata, M. Human and Avian Extraintestinal Pathogenic Escherichia coli: Infections, Zoonotic Risks, and Antibiotic Resistance Trends. Foodborne Pathog. Dis. 2013, 10, 916–932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, J.R.; Russo, T.A. Extraintestinal pathogenic Escherichia coli: “The other bad E coli”. J. Lab. Clin. Med. 2002, 139, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Flores-Mireles, A.L.; Walker, J.N.; Caparon, M.; Hultgren, S.J. Urinary tract infections: Epidemiology, mechanisms of infection and treatment options. Nat. Rev. Microbiol. 2015, 13, 269–284. [Google Scholar] [CrossRef] [PubMed]
- Laupland, K.B. Incidence of bloodstream infection: A review of population-based studies. Clin. Microbiol. Infect. 2013, 19, 492–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russo, T.A.; Johnson, J.R. Medical and economic impact of extraintestinal infections due to Escherichia coli: Focus on an increasingly important endemic problem. Microbes Infect. 2003, 5, 449–456. [Google Scholar] [CrossRef]
- Johnson, J.R.; Murray, A.C.; Gajewski, A.; Sullivan, M.; Snippes, P.; Kuskowski, M.A.; Smith, K.E. Isolation and molecular characterization of nalidixic acid-resistant extraintestinal pathogenic Escherichia coli from retail chicken products. Antimicrob. Agents Chemother. 2003, 47, 2161–2168. [Google Scholar] [CrossRef] [Green Version]
- Gomi, R.; Matsuda, T.; Matsumura, Y.; Yamamoto, M.; Tanaka, M.; Ichiyama, S.; Yoneda, M. Whole-Genome Analysis of Antimicrobial-Resistant and Extraintestinal Pathogenic Escherichia coli in River Water. Appl. Environ. Microbiol. 2016, 83, e02703-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhi, S.; Stothard, P.; Banting, G.; Scott, C.; Huntley, K.; Ryu, K.; Otto, S.; Ashbolt, N.; Checkley, S.; Dong, T.; et al. Characterization of water treatment-resistant and multidrug-resistant urinary pathogenic Escherichia coli in treated wastewater. Water Res. 2020, 182, 115827. [Google Scholar] [CrossRef]
- Rayasam, S.D.G.; Ray, I.; Smith, K.R.; Riley, L.W. Extraintestinal Pathogenic Escherichia coli and Antimicrobial Drug Resistance in a Maharashtrian Drinking Water System. Am. J. Trop. Med. Hyg. 2019, 100, 1101–1104. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.R.; Porter, S.B.; Johnston, B.; Thuras, P.; Clock, S.; Crupain, M.; Rangan, U. Extraintestinal Pathogenic and Antimicrobial-Resistant Escherichia coli, Including Sequence Type 131 (ST131), from Retail Chicken Breasts in the United States in 2013. Appl. Environ. Microbiol. 2017, 83, e02956-16. [Google Scholar] [CrossRef] [Green Version]
- Díaz-Jiménez, D.; García-Meniño, I.; Fernández, J.; García, V.; Mora, A. Chicken and turkey meat: Consumer exposure to multidrug-resistant Enterobacteriaceae including mcr-carriers, uropathogenic E. coli and high-risk lineages such as ST131. Int. J. Food Microbiol. 2020, 331, 108750. [Google Scholar] [CrossRef]
- Jakobsen, L.; Spangholm, D.J.; Pedersen, K.; Jensen, L.B.; Emborg, H.-D.; Agersø, Y.; Aarestrup, F.M.; Hammerum, A.M.; Frimodt-Møller, N. Broiler chickens, broiler chicken meat, pigs and pork as sources of ExPEC related virulence genes and resistance in Escherichia coli isolates from community-dwelling humans and UTI patients. Int. J. Food Microbiol. 2010, 142, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Jakobsen, L.; Garneau, P.; Kurbasic, A.; Bruant, G.; Stegger, M.; Harel, J.; Jensen, K.S.; Brousseau, R.; Hammerum, A.M.; Frimodt-Møller, N. Microarray-based detection of extended virulence and antimicrobial resistance gene profiles in phylogroup B2 Escherichia coli of human, meat and animal origin. J. Med Microbiol. 2011, 60, 1502–1511. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, J.C.; Filho, R.A.C.P.; Kuaye, A.P.Y.; Andrade, L.N.; Chang, Y.-F.; Darini, A.L.C. Virulence potential of commensal multidrug resistant Escherichia coli isolated from poultry in Brazil. Infect. Genet. Evol. 2018, 65, 251–256. [Google Scholar] [CrossRef] [Green Version]
- Bin Kim, Y.; Yoon, M.Y.; Ha, J.S.; Seo, K.W.; Noh, E.B.; Son, S.H.; Lee, Y.J. Molecular characterization of avian pathogenic Escherichia coli from broiler chickens with colibacillosis. Poult. Sci. 2020, 99, 1088–1095. [Google Scholar] [CrossRef]
- Subedi, M.; Luitel, H.; Devkota, B.; Bhattarai, R.K.; Phuyal, S.; Panthi, P.; Shrestha, A.; Chaudhary, D.K. Antibiotic resistance pattern and virulence genes content in avian pathogenic Escherichia coli (APEC) from broiler chickens in Chitwan, Nepal. BMC Veter. Res. 2018, 14, 113. [Google Scholar] [CrossRef]
- Johnson, T.J.; Wannemuehler, Y.; Doetkott, C.; Rosenberger, S.C.; Nolan, L.K. Identification of minimal predictors of avian pathogenic Escherichia coli virulence for use as a rapid diagnostic tool. J. Clin. Microbiol. 2008, 46, 3987–3996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saidenberg, A.B.S.; Stegger, M.; Price, L.B.; Johannesen, T.B.; Aziz, M.; Cunha, M.P.; Moreno, A.M.; Knöbl, T. mcr-Positive Escherichia coli ST131-H22 from Poultry in Brazil. Emerg. Infect. Dis. 2020, 26, 1951–1954. [Google Scholar] [CrossRef]
- Mohamed, M.A.; Shehata, M.A.; Rafeek, E. Virulence Genes Content and Antimicrobial Resistance in Escherichia coli from Broiler Chickens. Veter. Med. Int. 2014, 2014, 195189. [Google Scholar] [CrossRef] [Green Version]
- Stromberg, Z.R.; Johnson, J.R.; Fairbrother, J.M.; Kilbourne, J.; Van Goor, A.; Curtiss, R.; Mellata, M. Evaluation of Escherichia coli isolates from healthy chickens to determine their potential risk to poultry and human health. PLoS ONE 2017, 12, e0180599. [Google Scholar] [CrossRef] [Green Version]
- Bélanger, L.; Garenaux, A.; Harel, J.; Boulianne, M.; Nadeau, E.; Dozois, C.M. Escherichia coli from animal reservoirs as a potential source of human extraintestinal pathogenic E. coli. FEMS Immunol. Med Microbiol. 2011, 62, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Pitout, J.D. Extraintestinal Pathogenic Escherichia coli: A Combination of Virulence with Antibiotic Resistance. Front. Microbiol. 2012, 3, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aslam, M.; Toufeer, M.; Narvaez Bravo, C.; Lai, V.; Rempel, H.; Manges, A.; Diarra, M.S. Characterization of Extraintestinal Pathogenic Escherichia coli isolated from retail poultry meats from Alberta, Canada. Int. J. Food Microbiol. 2014, 177, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Tumbarello, M.; Sanguinetti, M.; Montuori, E.; Trecarichi, E.M.; Posteraro, B.; Fiori, B.; Citton, R.; D’Inzeo, T.; Fadda, G.; Cauda, R.; et al. Predictors of mortality in patients with bloodstream infections caused by extended-spectrum-beta-lactamase-producing Enterobacteriaceae: Importance of inadequate initial antimicrobial treatment. Antimicrob. Agents Chemother. 2007, 51, 1987–1994. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.R.; Kuskowski, M.A.; Gajewski, A.; Sahm, D.F.; Karlowsky, J.A. Virulence characteristics and phylogenetic background of multidrug-resistant and antimicrobial-susceptible clinical isolates of Escherichia coli from across the United States, 2000–2001. J. Infect. Dis. 2004, 190, 1739–1744. [Google Scholar] [CrossRef] [Green Version]
- Collignon, P.; Voss, A. China, what antibiotics and what volumes are used in food production animals? Antimicrob. Resist. Infect. Control. 2015, 4, 16. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Gao, S.; Jiao, X.; Liu, X.F. Prevalence of serogroups and virulence factors of Escherichia coli strains isolated from pigs with postweaning diarrhoea in eastern China. Vet. Microbiol. 2004, 103, 13–20. [Google Scholar] [CrossRef]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing; Twenty-Fifth Informational Supplement. CLSI Document M100-S25; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2015. [Google Scholar]
- CLSI. Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacterial Isolated from Animals; Approved Standard-Fourth Edition and Supplement, VET01A4E and VET01S3E; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2015. [Google Scholar]
- Rebelo, A.R.; Bortolaia, V.; Kjeldgaard, J.S.; Pedersen, S.K.; Leekitcharoenphon, P.; Hansen, I.M.; Guerra, B.; Malorny, B.; Borowiak, M.; Hammerl, J.A.; et al. Multiplex PCR for detection of plasmid-mediated colistin resistance determinants, mcr-1, mcr-2, mcr-3, mcr-4 and mcr-5 for surveillance purposes. Eurosurveillance 2018, 23, 17-00672. [Google Scholar] [CrossRef]
- Borowiak, M.; Baumann, B.; Fischer, J.; Thomas, K.; Deneke, C.; Hammerl, J.A.; Szabo, I.; Malorny, B. Development of a Novel mcr-6 to mcr-9 Multiplex PCR and Assessment of mcr-1 to mcr-9 Occurrence in Colistin-Resistant Salmonella enterica Isolates From Environment, Feed, Animals and Food (2011–2018) in Germany. Front. Microbiol. 2020, 11, 80. [Google Scholar] [CrossRef] [Green Version]
- Poirel, L.; Walsh, T.R.; Cuvillier, V.; Nordmann, P. Multiplex PCR for detection of acquired carbapenemase genes. Diagn. Microbiol. Infect. Dis. 2011, 70, 119–123. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Chen, C.; Cui, C.-Y.; Zhang, Y.; Liu, X.; Cui, Z.-H.; Ma, X.-Y.; Feng, Y.-J.; Fang, L.-X.; Lian, X.-L.; et al. Plasmid-encoded tet(X) genes that confer high-level tigecycline resistance in Escherichia coli. Nat. Microbiol. 2019, 4, 1457–1464. [Google Scholar] [CrossRef] [PubMed]
- Cavaco, L.M.; Hasman, H.; Xia, S.; Aarestrup, F.M. qnrD, a novel gene conferring transferable quinolone resistance in Salmonella enterica serovar Kentucky and Bovismorbificans strains of human origin. Antimicrob. Agents Chemother. 2008, 53, 603–608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Guo, Q.; Xu, X.; Wang, X.; Ye, X.; Wu, S.; Hooper, D.C.; Wang, M. New plasmid-mediated quinolone resistance gene, qnrC, found in a clinical isolate of Proteus mirabilis. Antimicrob. Agents Chemother. 2009, 53, 1892–1897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yue, L.; Jiang, H.-X.; Liao, X.-P.; Liu, J.-H.; Li, S.-J.; Chen, X.-Y.; Chen, C.-X.; Lü, D.-H.; Liu, Y.-H. Prevalence of plasmid-mediated quinolone resistance qnr genes in poultry and swine clinical isolates of Escherichia coli. Vet. Microbiol. 2008, 132, 414–420. [Google Scholar] [CrossRef] [PubMed]
- Yamane, K.; Wachino, J.-I.; Suzuki, S.; Arakawa, Y. Plasmid-mediated qepA gene among Escherichia coli clinical isolates from Japan. Antimicrob. Agents Chemother. 2008, 52, 1564–1566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.-T.; Song, F.-J.; Zou, M. Characterization of Highly Prevalent Plasmids Coharboring mcr-1, oqxAB, and blaCTX-M and Plasmids Harboring oqxAB and blaCTX-M in Escherichia coli Isolates from Food-Producing Animals in China. Microb. Drug Resist. 2019, 25, 108–119. [Google Scholar] [CrossRef]
- Brinas, L.; Moreno, M.A.; Zarazaga, M.; Porrero, C.; Saenz, Y.; García, M.; Dominguez, L.; Torres, C. Detection of CMY-2, CTX-M-14, and SHV-12 beta-lactamases in Escherichia coli fecal-sample isolates from healthy chickens. Antimicrob. Agents Chemother. 2003, 47, 2056–2058. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.H.; Wei, S.Y.; Ma, J.Y.; Zeng, Z.L.; Lu, D.H.; Yang, G.-X.; Chen, Z.-L. Detection and characterisation of CTX-M and CMY-2 beta-lactamases among Escherichia coli isolates from farm animals in Guangdong Province of China. Int. J. Antimicrob. Agents 2007, 29, 576–581. [Google Scholar] [CrossRef]
- Pai, H.; Kang, C.-I.; Byeon, J.-H.; Lee, K.-D.; Park, W.B.; Kim, H.-B.; Kim, E.-C.; Oh, M.-D.; Choe, K.-W. Epidemiology and clinical features of bloodstream infections caused by AmpC-type-beta-lactamase-producing Klebsiella pneumoniae. Antimicrob. Agents Chemother. 2004, 48, 3720–3728. [Google Scholar] [CrossRef] [Green Version]
- Kojima, A.; Ishii, Y.; Ishihara, K.; Esaki, H.; Asai, T.; Oda, C.; Tamura, Y.; Takahashi, T.; Yamaguchi, K. Extended-spectrum-beta-lactamase-producing Escherichia coli strains isolated from farm animals from 1999 to 2002: Report from the Japanese Veterinary Antimicrobial Resistance Monitoring Program. Antimicrob. Agents Chemother. 2005, 49, 3533–3537. [Google Scholar] [CrossRef] [Green Version]
- Hou, J.; Huang, X.; Deng, Y.; He, L.; Yang, T.; Zeng, Z.; Chen, Z.; Liu, J.-H. Dissemination of the fosfomycin resistance gene fosA3 with CTX-M beta-lactamase genes and rmtB carried on IncFII plasmids among Escherichia coli isolates from pets in China. Antimicrob. Agents Chemother. 2012, 56, 2135–2138. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Chen, Z.-L.; Liu, J.-H.; Zeng, Z.-L.; Ma, J.-Y.; Jiang, H.-X. Emergence of RmtB methylase-producing Escherichia coli and Enterobacter cloacae isolates from pigs in China. J. Antimicrob. Chemother. 2007, 59, 880–885. [Google Scholar] [CrossRef] [Green Version]
- Debroy, C.; Roberts, E.; Fratamico, P.M. Detection of O antigens in Escherichia coli. Anim. Heal. Res. Rev. 2011, 12, 169–185. [Google Scholar] [CrossRef]
- Gautom, R.K. Rapid pulsed-field gel electrophoresis protocol for typing of Escherichia coli O157:H7 and other gram-negative organisms in 1 day. J. Clin. Microbiol. 1997, 35, 2977–2980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wirth, T.; Falush, D.; Lan, R.; Colles, F.; Mensa, P.; Wieler, L.H.; Karch, H.; Reeves, P.R.; Maiden, M.C.J.; Ochman, H.; et al. Sex and virulence in Escherichia coli: An evolutionary perspective. Mol. Microbiol. 2006, 60, 1136–1151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, C.; Wang, Y.; Shi, X.; Wang, S.; Ren, H.; Shen, Z.; Wang, Y.; Lin, J.; Wang, S. Rapid rise of the ESBL and mcr-1 genes in Escherichia coli of chicken origin in China, 2008–2014. Emerg. Microbes Infect. 2018, 7, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FDA. Food and Drug Administration (2010) CVM Updates-CVM Reports on Antimicrobials Sold or Distributed for Food-Producing Animals; Food Drug Admin: Silver Spring, MD, USA, 2010.
- Ferjani, S.; Saidani, M.; Amine, F.S.; Boubaker, I.B.-B. Prevalence and characterization of plasmid-mediated quinolone resistance genes in extended-spectrum beta-lactamase-producing Enterobacteriaceae in a Tunisian hospital. Microb. Drug Resist. 2015, 21, 158–166. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.-T.; Yang, Q.-E.; Li, L.; Sun, J.; Liao, X.-P.; Fang, L.-X.; Yang, S.-S.; Deng, H.; Liu, Y.-H. Dissemination and characterization of plasmids carrying oqxAB-blaCTX-M genes in Escherichia coli isolates from food-producing animals. PLoS ONE 2013, 8, e73947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chmielarczyk, A.; Pobiega, M.; De Champs, C.; Wojkowska-Mach, J.; Różańska, A.; Heczko, P.B.; Guillard, T.; Bulanda, M. The High Prevalence of Plasmid-Mediated Quinolone Resistance Among Very Low Birth-Weight Infants in Poland. Microb. Drug Resist. 2015, 21, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Brower, C.H.; Mandal, S.; Hayer, S.; Sran, M.; Zehra, A.; Patel, S.J.; Kaur, R.; Chatterjee, L.; Mishra, S.; Das, B.; et al. The Prevalence of Extended-Spectrum Beta-Lactamase-Producing Multidrug-Resistant Escherichia Coli in Poultry Chickens and Variation According to Farming Practices in Punjab, India. Environ. Health Perspect. 2017, 125, 077015. [Google Scholar] [CrossRef] [Green Version]
- Clermont, O.; Olier, M.; Hoede, C.; Diancourt, L.; Brisse, S.; Keroudean, M.; Glodt, J.; Picard, B.; Oswald, E.; Denamur, E. Animal and human pathogenic Escherichia coli strains share common genetic backgrounds. Infect. Genet. Evol. 2011, 11, 654–662. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, N.M.; Johnson, J.R.; Johnston, B.; Curtiss, R., 3rd; Mellata, M. Zoonotic potential of Escherichia coli isolates from retail chicken meat products and eggs. Appl. Environ. Microbiol. 2015, 81, 1177–1187. [Google Scholar] [CrossRef] [Green Version]
- Mora, A.; López, C.; Herrera, A.; Viso, S.; Mamani, R.; Dhabi, G.; Alonso, M.P.; Blanco, M.; Blanco, J.E.; Blanco, J. Emerging avian pathogenic Escherichia coli strains belonging to clonal groups O111:H4-D-ST2085 and O111:H4-D-ST117 with high virulence-gene content and zoonotic potential. Vet. Microbiol. 2012, 156, 347–352. [Google Scholar] [CrossRef]
- Maluta, R.P.; Logue, C.M.; Casas, M.R.T.; Meng, T.; Guastalli, E.A.L.; Rojas, T.C.G.; Montelli, A.C.; Sadatsune, T.; de Carvalho Ramos, M.; Nolan, L.K.; et al. Overlapped sequence types (STs) and serogroups of avian pathogenic (APEC) and human extra-intestinal pathogenic (ExPEC) Escherichia coli isolated in Brazil. PLoS ONE 2014, 9, e105016. [Google Scholar] [CrossRef] [PubMed]
- Moulin-Schouleur, M.; Répérant, M.; Laurent, S.; Brée, A.; Mignon-Grasteau, S.; Germon, P.; Rasschaert, D.; Schouler, C. Extraintestinal pathogenic Escherichia coli strains of avian and human origin: Link between phylogenetic relationships and common virulence patterns. J. Clin. Microbiol. 2007, 45, 3366–3376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.-M.; Liao, X.-P.; Zhang, W.-J.; Jiang, H.-X.; Sun, J.; Zhang, M.-J.; He, X.-F.; Lao, D.-X.; Liu, Y.-H. Prevalence of serogroups, virulence genotypes, antimicrobial resistance, and phylogenetic background of avian pathogenic Escherichia coli in south of China. Foodborne Pathog. Dis. 2010, 7, 1099–1106. [Google Scholar] [CrossRef]
- Johnson, T.J.; Wannemuehler, Y.; Johnson, S.J.; Stell, A.L.; Doetkott, C.; Johnson, J.R.; Kim, K.S.; Spanjaard, L.; Nolan, L.K. Comparison of extraintestinal pathogenic Escherichia coli strains from human and avian sources reveals a mixed subset representing potential zoonotic pathogens. Appl. Environ. Microbiol. 2008, 74, 7043–7050. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.R.; Oswald, E.; O’Bryan, T.T.; Kuskowski, M.A.; Spanjaard, L. Phylogenetic distribution of virulence-associated genes among Escherichia coli isolates associated with neonatal bacterial meningitis in the Netherlands. J. Infect. Dis. 2002, 185, 774–784. [Google Scholar] [CrossRef] [Green Version]
- Bonacorsi, S.; Clermont, O.; Houdouin, V.; Cordevant, C.; Brahimi, N.; Marecat, A.; Tinsley, C.; Nassif, X.; Lange, M.; Bingen, E. Molecular analysis and experimental virulence of French and North American Escherichia coli neonatal meningitis isolates: Identification of a new virulent clone. J. Infect. Dis. 2003, 187, 1895–1906. [Google Scholar] [CrossRef]
- Denisuik, A.J.; Lagacé-Wiens, P.R.S.; Pitout, J.D.; Mulvey, M.R.; Simner, P.J.; Tailor, F.; Karlowsky, J.A.; Hoban, D.J.; Adam, H.J.; Zhanel, G.G.; et al. Molecular epidemiology of extended-spectrum beta-lactamase-, AmpC beta-lactamase- and carbapenemase-producing Escherichia coli and Klebsiella pneumoniae isolated from Canadian hospitals over a 5 year period: CANWARD 2007-11. J. Antimicrob. Chemother. 2013, 68 (Suppl. 1), i57–i65. [Google Scholar] [CrossRef] [Green Version]
- Karlowsky, J.A.; Denisuik, A.J.; Lagacé-Wiens, P.R.S.; Adam, H.J.; Baxter, M.R.; Hoban, D.J.; Zhanel, G.G. In Vitro activity of fosfomycin against Escherichia coli isolated from patients with urinary tract infections in Canada as part of the CANWARD surveillance study. Antimicrob. Agents Chemother. 2013, 58, 1252–1256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Baño, J.; Gutiérrez-Gutiérrez, B.; Machuca, I.; Pascual, A. Treatment of Infections Caused by Extended-Spectrum-Beta-Lactamase-, AmpC-, and Carbapenemase-Producing Enterobacteriaceae. Clin. Microbiol. Rev. 2018, 31, e00079-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, F.; Zhang, R.; Yang, Y.; Li, H.; Wang, J.; Lan, J.; Li, P.; Zhu, Y.; Xie, Z.; Jiang, S. Occurrence and Molecular Characteristics of Mcr-1-Positive Escherichia coli from Healthy Meat Ducks in Shandong Province of China. Animals 2020, 10, 1299. [Google Scholar] [CrossRef]
- Ranjan, A.; Shaik, S.; Mondal, A.; Nandanwar, N.; Hussain, A.; Semmler, T.; Kumar, N.; Tiwari, S.K.; Jadhav, S.; Wieler, L.H.; et al. Molecular Epidemiology and Genome Dynamics of New Delhi Metallo-beta-Lactamase-Producing Extraintestinal Pathogenic Escherichia coli Strains from India. Antimicrob. Agents Chemother. 2016, 60, 6795–6805. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Antimicrobials | Resistance Rates of Isolates (%) * | |||
|---|---|---|---|---|
| Total (n = 926) | Layer Farms (n = 371) | White-Feather Broiler Farms (n = 78) | Live Poultry Markets (n = 477) | |
| β-lactams | - | - | - | - |
| AMP | 87.1 | 83.0 a | 100 b | 88.3 c |
| CTF | 44.7 | 39.4 a | 98.7 b | 40.0 a |
| CTX | 41.8 | 26.1 a | 100 b | 41.5 c |
| MEM | 4.9 | 1.9 a | 48.7 b | 0.0 c |
| Quinolones | - | - | - | - |
| NAL | 77.1 | 77.6 a | 96.2 b | 73.6 a |
| ENR | 58.4 | 59.3 a | 89.7 b | 52.6 a |
| CIP | 48.4 | 45.8 a | 83.3 b | 44.7 a |
| LEV | 36.3 | 40.2 a | 51.3 a | 30.8 b |
| Tetracyclines | - | - | - | - |
| TET | 89.3 | 87.3 a | 97.4 b | 89.5 a |
| DOX | 83.8 | 81.1 a | 97.4 b | 83.6 a |
| TIG | 0 | 0 | 0 | 0 |
| Aminoglycosides | - | - | - | - |
| STR | 60.7 | 63.9 a | 83.3 b | 54.5 c |
| KAN | 50.5 | 46.6 a | 96.2 b | 46.1 a |
| GEN | 31.9 | 26.1 a | 73.1 b | 29.6 a |
| AMK | 8.9 | 5.1 a | 35.9 b | 7.3 a |
| Polypeptides | - | - | - | - |
| COL | 17.0 | 4.9 a | 73.1 b | 17.2 c |
| Others | - | - | - | - |
| FFC | 69.1 | 68.7 a | 94.9 b | 65.2 a |
| FOS | 20.6 | 10.5 a | 78.2 b | 19.1 c |
| Resistance Genes | No. of Positive Isolates (%) * | |||
|---|---|---|---|---|
| Total (n = 926) | Layer Farms (n = 371) | White-Feather Broiler Farms 9 (n = 78) | Live Poultry Markets (n = 477) | |
| Carbapenemases | - | - | - | - |
| blaNDM | 45 (4.9) | 11 (3.0) a | 34 (43.6) b | 0 (0) c |
| ESBLs | - | - | - | - |
| blaCTX-M-9G | 222 (24.0) | 52 (14.0) a | 54 (69.2) b | 116 (24.3) c |
| blaCTX-M-1G | 130 (14.0) | 43 (11.6) a | 32 (41.0) b | 55 (11.5) a |
| pAmpC | - | - | - | - |
| blaCMY-2 | 53 (5.7) | 11 (3.0) a | 3 (3.8) ab | 39 (8.2) b |
| blaDHA-1 | 3 (0.3) | 1 (0.3) a | 0 (0.0) a | 2 (0.4) a |
| MCR | - | - | - | - |
| mcr-1 | 157 (17.0) | 22 (5.9) a | 53 (67.9) b | 82 (17.2) c |
| PMQR | - | - | - | - |
| oqxAB | 181 (19.5) | 41 (11.1) a | 22 (28.2) b | 118 (24.7) b |
| qnrB * | 34 (3.7) | 7 (1.9) a | 1 (1.3) ab | 26 (5.5) b |
| qnrS | 311 (33.6) | 140 (37.7) a | 12 (15.4) b | 159 (33.3) a |
| qnrD * | 21 (2.3) | 3 (0.8) a | 0 (0.0) ab | 18 (3.8) b |
| PFR | - | - | - | - |
| fosA3 | 189 (20.4) | 39 (10.5) a | 61 (78.2) b | 89 (18.7) c |
| fosA | 2 (0.2) | 0 (0.0) a | 0 (0.0) a | 2 (0.4) a |
| Others | - | - | - | - |
| rmtB | 35 (3.8) | 2 (0.5) a | 19 (24.4) b | 14 (2.9) c |
| Location | Origins | No. of Farms/Markets | Year | ExPEC Isolates (%)/ Total Isolates |
|---|---|---|---|---|
| Weifang in Shandong | White-feather broiler farm | 1 | 2015 | 6 (7.7%)/78 |
| Hefei in Anhui | Layer farm | 1 | 2015 | 1 (6.3%)/16 |
| Liaocheng in Shandong | Layer farm | 1 | 2015 | 2 (4.9%)/41 |
| Binzhou in Shandong | Layer farms | 2 | 2016 | 1 (0.7%)/135 |
| Xi’an in Shanxi | Layer farm | 1 | 2015 | 2 (2.1%)/97 |
| Qingdao in Shandong | Layer farms | 2 | 2017 | 0/82 |
| Linyi in Shandong | Live poultry market | 1 | 2015 | 3 (4.4%)/68 |
| Qingdao in Shandong | Live poultry markets | 11 | 2015 | 6 (3.0%)/199 |
| Yantai in Shandong | Live poultry markets | 2 | 2015 | 1 (1.8%)/55 |
| Zaozhuang in Shandong | Live poultry market | 1 | 2015 | 0/57 |
| Zibo in Shandong | Live poultry market | 1 | 2015 | 0/67 |
| Weifang in Shandong | Live poultry market | 1 | 2015 | 0/31 |
| - | White-feather broiler farm | 1 | - | 6 (7.7%)/78 |
| - | Layer farms | 7 | - | 6 (1.6%)/371 |
| - | Live poultry markets | 17 | - | 10 (2.1%)/477 |
| Strain (ST Types) | Source | City | Serotype # | ExPEC-Defining Markers | Resistance Phenotype | Resistance Genes |
|---|---|---|---|---|---|---|
| WF1-3-24 | White-feather broiler farm A | Weifang | ND | KpsM II, iutA | mcr-1, blaCTX-M-1G, fosA3, rmtB, oqxAB, floR | COL, CTX, CTF, AMP, FOS, AMK, GEN, KAN, STR, NAL, CIP, ENR, LEV, FFL, DOX, TET, OLA, SXT |
| WF1-5-10 | White-feather broiler farm A | Weifang | O86 | KpsM II, iutA | blaCTX-M-9G, fosA3, rmtB, floR | CTX, CTF, AMP, AMK, GEN, KAN, STR, NAL, CIP, ENR, LEV, FFL, DOX, TET, SXT |
| WF1-5-13 | White-feather broiler farm A | Weifang | ND | papA, iutA | mcr-1, blaNDM-5, blaCTX-M-9G, blaCTX-M-1G, fosA3, floR | COL, MEM, CTX, CTF, AMP, FOS, KAN, STR, NAL, CIP, ENR, LEV, FFL, DOX, TET, SXT |
| WF1-5-21 | White-feather broiler farm A | Weifang | O78 | papA, iutA | mcr-1, blaCTX-M-9G, fosA3, rmtB, floR | COL, CTX, CTF, AMP, FOS, AMK, GEN, KAN, STR, NAL, FFL, DOX, TET, SXT |
| WF1-5-40 | White-feather broiler farm A | Weifang | ND | KpsM II, iutA | mcr-1, blaNDM-1, blaCTX-M-9G, fosA3, rmtB, floR | COL, MEM, CTX, CTF, AMP, FOS, AMK, GEN, KAN, STR, FFL, DOX, TET, SXT |
| WF1-5-43 | White-feather broiler farm A | Weifang | O86 | KpsM II, iutA | blaCTX-M-9G, fosA3 | CTX, CTF, AMP, FOS, KAN, STR, NAL, CIP, ENR, LEV, DOX, TET, SXT |
| AH-234 | Layer farm B | Hefei | O78 | KpsM II, iutA | blaCTX-M-9G, blaCMY-2, floR | CTX, CTF, AMP, AMK, GEN, KAN, STR, NAL, CIP, ENR, LEV *, FFL, TET, SXT |
| G-1-5 | Layer farm C | Liaocheng | ND | KpsM II, iutA | mcr-1 | COL, AMP, KAN, STR, NAL, DOX, TET, SXT |
| GX-J-1-29 | Layer farm C | Liaocheng | O78 | KpsM II, iutA | qnrS, floR | AMP, FFL, DOX, TET, SXT |
| BZ-J-1-4 | Layer farm D | Binzhou | O78 | KpsM II, iutA | blaCTX-M-1G, fosA3 | CTX, CTF, AMP, FOS, STR, NAL, FFL, DOX, TET, SXT |
| XA-8N-7 | Layer farm E | Xi’an | O83 | KpsM II, iutA | blaCTX-M-1G, floR | CTX, CTF, AMP, KAN, NAL, ENR, LEV, FFL, DOX, TET, SXT |
| XA-3N-1 | Layer farm E | Xi’an | O18 | KpsM II, iutA | blaCTX-M-1G, floR | CTX, CTF, AMP, KAN, STR, NAL, CIP, ENR, LEV, FFL, DOX, TET, SXT |
| LS-A-3 | Live poultry market 1 | Linyi | O78 | KpsM II, iutA | blaCTX-M-9G, fosA3, floR | CTX, CTF, AMP, FOS, KAN, STR, NAL, CIP, ENR, LEV, FFL, DOX, TET, SXT |
| LS-A-7 | Live poultry market 1 | Linyi | O78 | papA, iutA | blaCTX-M-1G, fosA3, oqxAB, floR | CTX, CTF, AMP, FOS, KAN, STR, NAL, CIP, ENR, LEV, FFL, DOX, TET, OLA, SXT |
| LS-A-27 | Live poultry market 1 | Linyi | O78 | KpsM II, iutA | blaCTX-M-9G, floR | CTX, CTF, AMP, KAN, STR, NAL, ENR, FFL, DOX, TET, SXT |
| YJ-JC-8 | Live poultry market 2 | Qingdao | O78 | papA, iutA | oqxAB | AMP, AMK, GEN, KAN, STR, NAL, CIP, ENR, LEV, FFL, DOX, TET, OLA, SXT |
| JS-JC-2 | Live poultry market 3 | Qingdao | O26 | KpsM II, iutA | floR | CTX, CTF, AMP, FOS, GEN, KAN, STR, NAL, CIP, ENR, LEV, FFL, DOX, TET, SXT |
| JS-JC-4 | Live poultry market 3 | Qingdao | ND | KpsM II, iutA | qnrS | AMP, KAN, STR, NAL, DOX, TET, SXT |
| JS-JC-7 | Live poultry market 3 | Qingdao | O26 | KpsM II, iutA | qnrS | AMP, STR, DOX, TET, SXT |
| JS-JC-9 | Live poultry market 3 | Qingdao | ND | KpsM II, iutA | blaCTX-M-9G, qnrS | CTX, CTF, AMP, KAN, STR, FFL, DOX, TET, SXT |
| L2-JC-34 | Live poultry market 4 | Qingdao | O45 | KpsM II, iutA | mcr-1, blaCTX-M-9G, fosA3, floR | COL, CTX, CTF, AMP, FOS, GEN, KAN, NAL, FFL, SXT |
| HY-1-4 | Live poultry market 5 | Yantai | O78 | KpsM II, iutA | - | AMP, NAL, ENR, FFL, DOX, TET, SXT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zou, M.; Ma, P.-P.; Liu, W.-S.; Liang, X.; Li, X.-Y.; Li, Y.-Z.; Liu, B.-T. Prevalence and Antibiotic Resistance Characteristics of Extraintestinal Pathogenic Escherichia coli among Healthy Chickens from Farms and Live Poultry Markets in China. Animals 2021, 11, 1112. https://doi.org/10.3390/ani11041112
Zou M, Ma P-P, Liu W-S, Liang X, Li X-Y, Li Y-Z, Liu B-T. Prevalence and Antibiotic Resistance Characteristics of Extraintestinal Pathogenic Escherichia coli among Healthy Chickens from Farms and Live Poultry Markets in China. Animals. 2021; 11(4):1112. https://doi.org/10.3390/ani11041112
Chicago/Turabian StyleZou, Ming, Ping-Ping Ma, Wen-Shuang Liu, Xiao Liang, Xu-Yong Li, You-Zhi Li, and Bao-Tao Liu. 2021. "Prevalence and Antibiotic Resistance Characteristics of Extraintestinal Pathogenic Escherichia coli among Healthy Chickens from Farms and Live Poultry Markets in China" Animals 11, no. 4: 1112. https://doi.org/10.3390/ani11041112