
Histological Study of Suprabranchial Chamber Membranes in Anabantoidei and Clariidae Fishes

 , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
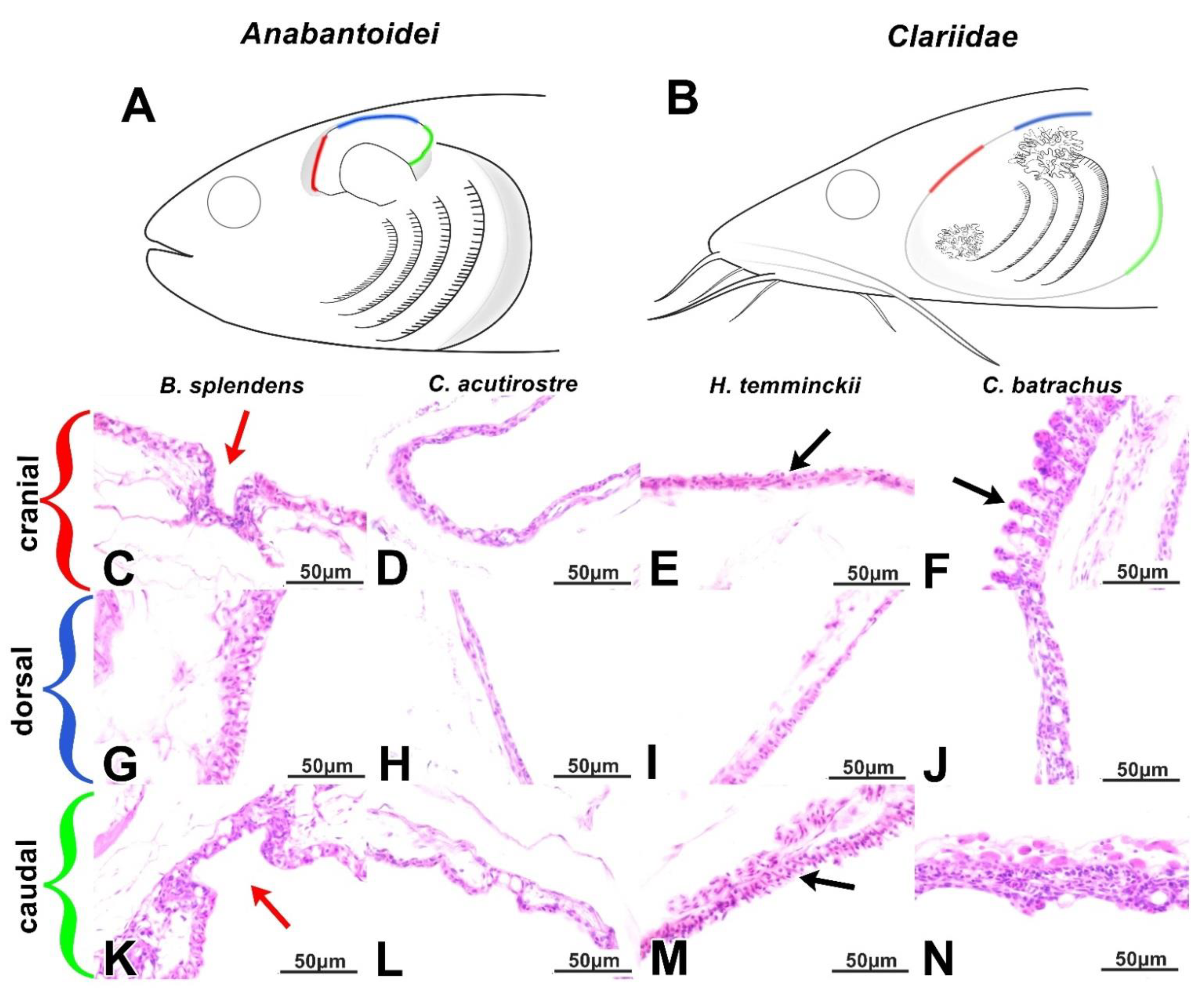
3.1. Gills and Gill Cavity
3.2. Labyrinth Organ (LO) and Suprabranchial Cavity of Anabantoidei Species
3.3. Dendritic Organ (DO) and Suprabranchial Cavity of Clariidae Species
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Graham, J.B. Air-Breathing Fishes: Evolution, Diversity, and Adaptation. Available online: https://books.google.pl/books?hl=pl&lr=&id=JQh3D3BFCZwC&oi=fnd&pg=PP1&dq=graham+anabantoidei+1997&ots=cQKY65Qk2A&sig=FxSoI_y0_yV2egwyrUC8V11a1-k&redir_esc=y#v=onepage&q=graham anabantoidei 1997&f=false (accessed on 8 February 2021).
- Zaccone, G.; Lauriano, E.R.; Capillo, G.; Kuciel, M. Air- breathing in fish: Air- breathing organs and control of respiration: Nerves and neurotransmitters in the air-breathing organs and the skin. Acta Histochem. 2018, 120, 630–641. [Google Scholar] [CrossRef]
- Gisbert, E.; Ruban, G.I. Ontogenetic behaviour of Siberian sturgeon, Acipenser baerii: A synthesis between laboratory tests and field data. Environ. Biol. Fishes 2003, 67, 311–319. [Google Scholar] [CrossRef]
- Satora, L.; Kozioł, K.; Waldman, W.; Mytych, J. Differential expression of epidermal growth factor receptor (EGFR) in stomach and diverticulum of Otocinclus affinis (Steindachner, 1877) as a potential element of the epithelium remodeling mechanism. Acta Histochem. 2019, 121, 151–155. [Google Scholar] [CrossRef] [PubMed]
- Rüber, L. Labyrinth fishes (Anabantoidei). Timetree Life 2009, 344–347. [Google Scholar]
- Rüber, L.; Britz, R.; Zardoya, R. Molecular phylogenetics and evolutionary diversification of labyrinth fishes (Perciformes: Anabantoidei). Syst. Biol. 2006, 55, 374–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, R.A.; Britz, R.; Rüber, L. Phylogenetic systematics of leaffishes (Teleostei: Polycentridae, Nandidae). J. Zool. Syst. Evol. Res. 2015, 53, 259–272. [Google Scholar] [CrossRef]
- Munshi, J.S. The accessory respiratory organs of Clarias batrachus (Linn.). J. Morphol. 1961, 109, 115–139. [Google Scholar] [CrossRef] [PubMed]
- Nelson, J.S.; Schultze, H.-P.; Wilson, M.V.H. Origin and Phylogenetic Interrelationships of Teleosts Honoring Gloria Arratia. 2010. Available online: file:///C:/Users/MDPI/AppData/Local/Temp/NelsonSchultzeWilson2010OriginPhylogRelatTeleostscover.pdf (accessed on 8 February 2021).
- Degani, G. Mitochondrial DNA sequence analysis in Anabantoidei fish. Adv. Biol. Chem. 2013, 03, 347–355. [Google Scholar] [CrossRef] [Green Version]
- Berra, T.M. Freshwater Fish Distribution. Available online: https://books.google.pl/books?hl=pl&lr=&id=EvP5x2xPx9wC&oi=fnd&pg=PP2&dq=berra+fish+breathing&ots=JwivI_X1wI&sig=Kp3VYLXCeIq_LzJBD_uVvPhEfBk&redir_esc=y#v=onepage&q=berra fish breathing&f=false (accessed on 8 February 2021).
- Huang, C.-Y.; Lin, C.-P.; Lin, H.-C. Morphological and Biochemical Variations in the Gills of 12 Aquatic Air-Breathing Anabantoid Fish. Sour. Physiol. Biochem. Zool. Ecol. Evol. Approaches 2011, 84, 125–134. [Google Scholar] [CrossRef]
- Blank, T.; Burggren, W. Hypoxia-induced developmental plasticity of the gills and air-breathing organ of Trichopodus trichopterus. J. Fish Biol. 2014, 84, 808–826. [Google Scholar] [CrossRef]
- Damsgaard, C.; Baliga, V.B.; Bates, E.; Burggren, W.; McKenzie, D.J.; Taylor, E.; Wright, P.A. Evolutionary and cardio-respiratory physiology of air-breathing and amphibious fishes. Acta Physiol. 2020, 228, e13406. [Google Scholar] [CrossRef]
- Mendez-Sanchez, J.F.; Burggren, W.W. Environmental modulation of the onset of air breathing and survival of Betta splendens and Trichopodus trichopterus. J. Fish Biol. 2014, 84, 794–807. [Google Scholar] [CrossRef]
- Wiley, E.O.; Johnson, G.D. A Teleost Classification Based on Monophyletic Groups. 2010. Available online: https://repository.si.edu/bitstream/handle/10088/9786/vz_10Wiley-Johnson-2010-Teleost_Classification.pdf (accessed on 8 February 2021).
- Hughes, G.M.; Munshi, J.S. Fine structure of the respiratory surfaces of an air-breathing fish, the climbing perch anabas testudineus (Bloch). Nature 1968, 219, 1382–1384. [Google Scholar] [CrossRef]
- Hughes, G.M.; Munshi, J.S. Nature of the air-breathing organs of the Indian fishes Channa, Amphipnous, Clarias and Saccobranchus as shown by electron microscopy. J. Zool. 1973, 170, 245–270. [Google Scholar] [CrossRef]
- Betancur, R.R.; Wiley, E.O.; Arratia, G.; Acero, A.; Bailly, N.; Miya, M.; Lecointre, G.; Ortí, G. Phylogenetic classification of bony fishes. BMC Evol. Biol. 2017, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, A.-E.; Mohamed, K.; Ahmed, S.-A.; Masoud, F. Anatomical, Light and Scanning Electron Microscopic Studies on the Air Breathing Dendretic Organ of the Sharp Tooth Catfish (Clarias gariepinus). J. Vet. Anat. 2008, 1, 29–37. [Google Scholar]
- Arratia, G. Catfish head skeleton—An overview. Catfishes 2003, 1, 3–46. [Google Scholar]
- Teugels, G.; Adriaens, D. Taxonomy and phylogeny of Clariidae—An overview. Catfishes 2003, 465–487. [Google Scholar]
- Mbanga, B.; van Dyk, C.; Maina, J.N. Morphometric and morphological study of the respiratory organs of the bimodally-breathing African sharptooth catfish (Clarias gariepinus): Burchell (1822). Zoology 2018, 130, 6–18. [Google Scholar] [CrossRef] [PubMed]
- Belão, T.C.; Zeraik, V.M.; Florindo, L.H.; Kalinin, A.L.; Leite, C.A.C.; Rantin, F.T. Control of cardiorespiratory function in response to hypoxia in an air-breathing fish, the African sharptooth catfish, Clarias gariepinus. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2015, 187, 130–140. [Google Scholar] [CrossRef]
- El-Hady, E.; El-Behery, E.I.; Ebraheim, L.L. Morpho-histological approach of African cat fish (Clarias gariepinus) respiratory system with mucocytes and arterial blood supply attribute. Int. J. Fish. Aquat. Stud. 2019, 7, 31–41. [Google Scholar]
- Chandra, S.; Banerjee, T.K. Histopathological analysis of the respiratory organs of Channa striata subjected to air exposure. Vet. Arh. 2004, 74, 37–52. [Google Scholar]
- Roy, B.C.; Ando, M.; Nakatani, M.; Okada, T.; Sawada, Y.; Itoh, T.; Tsukamasa, Y. Muscle fiber types, growth and development in the whole myotome of cultured Pacific bluefin tuna Thunnus orientalis. Fish. Sci. 2012, 78, 471–483. [Google Scholar] [CrossRef]
- Yan, H.Y. Auditory role of the suprabranchial chamber in gourami fish. J. Comp. Physiol. A Sens. Neural Behav. Physiol. 1998, 183, 325–333. [Google Scholar] [CrossRef]
- Bertmar, G.; Kapoor, B.G.; Miller, R.V. Epibranchial Organs in Lower Teleostean Fishes—An Example of Structural Adaptation. Int. R. Gen. Exp. Zool. 1969, 4, 1–48. [Google Scholar]
- Liem, K.F. Functional Morphology of the Integumentary, Respiratory, and Digestive Systems of the Synbranchoid Fish Monopterus albus. JSTOR 1967, 1967, 375–388. [Google Scholar] [CrossRef]
- Munshi, J.S.; Olson, K.R.; Ojha, J.; Ghosh, T.K. Morphology and vascular anatomy of the accessory respiratory organs of the air-breathing climbing perch, Anabas testudineus (Bloch). Am. J. Anat. 1986, 176, 321–331. [Google Scholar] [CrossRef]
- Olson, K.R.; Roy, P.K.; Ghosh, T.K.; Munshi, J.S. Microcirculation of gills and accessory respiratory organs from the air-breathing snakehead fish, Channa punctata, C. gachua, and C. marulius. Anat. Rec. 1994, 238, 92–107. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.; Wolf, J.; Braunbeck, T. OECD Guidance Document for the Diagnosis of Endocrine-Related Histopathology of Fish Gonads. 2009. Available online: https://www.webpages.uidaho.edu/fish511/Readings/Readings%202010/OECD%20guidance%20histology.pdf (accessed on 8 February 2021).
- Powers, D.A. Molecular ecology of teleost fish hemoglobins strategies for adapting to changing environments. Integr. Comp. Biol. 1980, 20, 139–162. [Google Scholar] [CrossRef]
- Wells, R.M.G.; Baldwin, J.; Seymour, R.S.; Baudinette, R.V.; Christian, K.; Bennett, M.B. Oxygen transport capacity in the air-breathing fish, Megalops cyprinoides: Compensations for strenuous exercise. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2003, 134, 45–53. [Google Scholar] [CrossRef]
- Ip, Y.K.; Chew, S.F. Air-breathing and excretory nitrogen metabolism in fishes. Acta Histochem. 2018, 120, 680–690. [Google Scholar] [CrossRef]
- Apriliani, N.S.; Supriyati, H.; Ja’far Luthfi, M.; Luthfi, M.J. Histological Study of Respiratory Organ of Betta sp. Proc. Int. Conf. Sci. Eng. 2019, 2, 181–184. [Google Scholar]
- Huang, L.; Yang, L.; Liu, J.; Cao, X. Comparative Histological Analysis of Intestines of Loach, Grass Carp and Catfish Provide Insights into Adaptive Characteristics in Air-Breathing Fish. Croat. J. Fish. 2020, 78, 91–98. [Google Scholar] [CrossRef]
- Maina, J.N. Functional morphology of the respiratory organs of the air-breathing fish with particular emphasis on the African catfishes, Clarias mossambicus and C. gariepinus. Acta Histochem. 2018, 120, 613–622. [Google Scholar] [CrossRef] [PubMed]
- Hsia, C.C.W.; Schmitz, A.; Lambertz, M.; Perry, S.F.; Maina, J.N. Evolution of air breathing: Oxygen homeostasis and the transitions from water to land and sky. Compr. Physiol. 2013, 3, 849–915. [Google Scholar] [CrossRef] [Green Version]
- Soliman, S.; Abd-Elhafeez, H.H.; Sa, S.; Hh, A.-E. Mesenchymal Cells in Cartilage Growth and Regeneration “An Immunohistochemical and Electron Microscopic Study. J. Cytol. Histol. 2016, 7, 437. [Google Scholar] [CrossRef] [Green Version]
- Peters, H.M. On the mechanism of air ventilaton in anabantoids (Pisces: Teleostei). Zoomorphologie 1978, 89, 93–123. [Google Scholar] [CrossRef]
- Maina, J.N.; Maloiy, G.M.O. The morphology of the respiratory organs of the African air-breathing catfish (Clarias mossambicus): A light, electron and scanning microscopic study, with morphometric observations. J. Zool. 1986, 209, 421–445. [Google Scholar] [CrossRef]
- Munshi, J.S.; Singh, B.N. On the respiratory organs of Amphipnous cuchia (Ham. Buch.). J. Morphol. 1968, 124, 423–443. [Google Scholar] [CrossRef]
- Bettex Galland, M.; Hughes, G.M. Contractile filamentous material in the pillar cells of fish gills. J. Cell Sci. 1973, 13, 359–370. [Google Scholar]
- Munshi, J.S. The accessory respiratory organs of Anabas testudineus (Bloch) (Anabantidae, Pisces). Proc. Linn. Soc. London 1968, 179, 107–126. [Google Scholar] [CrossRef]
- Daniels, C.B.; Orgeig, S.; Sullivan, L.C.; Ling, N.; Bennett, M.B.; Schürch, S.; Val, A.L.; Brauner, C.J.; Luis Val, A.; Brauner, C.J. The Origin and Evolution of the Surfactant System in Fish: Insights into the Evolution of Lungs and Swim Bladders. Source Physiol. Biochem. Zool. Ecol. Evol. Approaches 2004, 77, 732–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daniels, C.B.; Skinner, C.H. The Composition and Function of Surface-Active Lipids in the Goldfish Swim Bladder. Physiol. Zool. 1994, 67, 1230–1256. [Google Scholar] [CrossRef]
- Daniels, C.B.; Orgeig, S. The comparative biology of pulmonary surfactant: Past, present and future. In Proceedings of the Comparative Biochemistry and Physiology—A Molecular and Integrative Physiology; Elsevier Inc.: Amsterdam, The Netherlands, 2001; Volume 129, pp. 9–36. [Google Scholar]
- Moron, S.E.; de Andrade, C.A.; Fernandes, M.N. Response of mucous cells of the gills of traíra (Hoplias malabaricus) and jeju (Hoplerythrinus unitaeniatus) (Teleostei: Erythrinidae) to hypo- and hyper-osmotic ion stress. Neotrop. Ichthyol. 2009, 7, 491–498. [Google Scholar] [CrossRef] [Green Version]
- Alkan, Z.; Oğuz, A.R. Investigation of gill mucus cells of Lake Van fish (Alburnus tarichi) during reproductive migration. Fish Physiol. Biochem. 2021, 1–11. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Species | Body Weight (g) | Length (cm) | ||
|---|---|---|---|---|
| Average | SD | Average | SD | |
| Betta splendens | 0.99 | 0.23 | 4.32 | 0.20 |
| Ctenopoma acutirostre | 0.91 | 0.06 | 3.64 | 0.09 |
| Helostoma temminckii | 2.55 | 0.36 | 5.80 | 0.27 |
| Clarias angolensis | 8.31 | 1.09 | 11.47 | 0.80 |
| Clarias batrachus | 3.04 | 0.73 | 8.04 | 0.69 |
| Species | Gills | Gill Chamber | Labyrinth Organ | Dendritic Organ |
|---|---|---|---|---|
| B. splendens | acidic, mixed | mixed | acidic, mixed | N/A |
| C. acutirostre | acidic, mixed | acidic | acidic, mixed | N/A |
| H. temminckii | acidic | mixed | acidic | N/A |
| C. angolensis | acidic | acidic, mixed | N/A | acidic |
| C. batrachus | strongly acidic | acidic, mixed | N/A | acidic, neutral (few) |
| Epithelial Height (µm) | ||||||||
|---|---|---|---|---|---|---|---|---|
| Species | Cranial | Dorsal | Caudal | LO/DO | ||||
| Average | SD | Average | SD | Average | SD | Average | SD | |
| B. splendens | 11.13 * | 5.32 | 14.04 * | 5.25 | 10.85 * | 2.64 | 6.19 | 1.61 |
| C. acutirostre | 10.26 * | 3.08 | 4.02 * | 1.36 | 15.28 * | 4.07 | 6.65 | 1.81 |
| H. temminckii | 17.19 * | 6.38 | 7.46 * | 1.34 | 13.86 * | 5.68 | 7.80 | 1.87 |
| C. angolensis | 31.97 * | 8.65 | 22.06 * | 5.17 | 39.96 * | 11.87 | 28.36 | 5.88 |
| C. batrachus | 23.66 * | 3.40 | 21.00 * | 4.25 | 33.43 * | 8.03 | 22.72 | 4.78 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adamek-Urbańska, D.; Błażewicz, E.; Sobień, M.; Kasprzak, R.; Kamaszewski, M. Histological Study of Suprabranchial Chamber Membranes in Anabantoidei and Clariidae Fishes. Animals 2021, 11, 1158. https://doi.org/10.3390/ani11041158
Adamek-Urbańska D, Błażewicz E, Sobień M, Kasprzak R, Kamaszewski M. Histological Study of Suprabranchial Chamber Membranes in Anabantoidei and Clariidae Fishes. Animals. 2021; 11(4):1158. https://doi.org/10.3390/ani11041158
Chicago/Turabian StyleAdamek-Urbańska, Dobrochna, Ewelina Błażewicz, Magdalena Sobień, Robert Kasprzak, and Maciej Kamaszewski. 2021. "Histological Study of Suprabranchial Chamber Membranes in Anabantoidei and Clariidae Fishes" Animals 11, no. 4: 1158. https://doi.org/10.3390/ani11041158