
Effect of a Polyphenol-Based Additive in Pig Diets in the Early Stages of Growth

 ,
,  ,
,  , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experiment 1 (Post-Weaning Trial)
2.1.1. Animals, Experimental Design, and Diets
2.1.2. Clinical and Biochemical Parameters
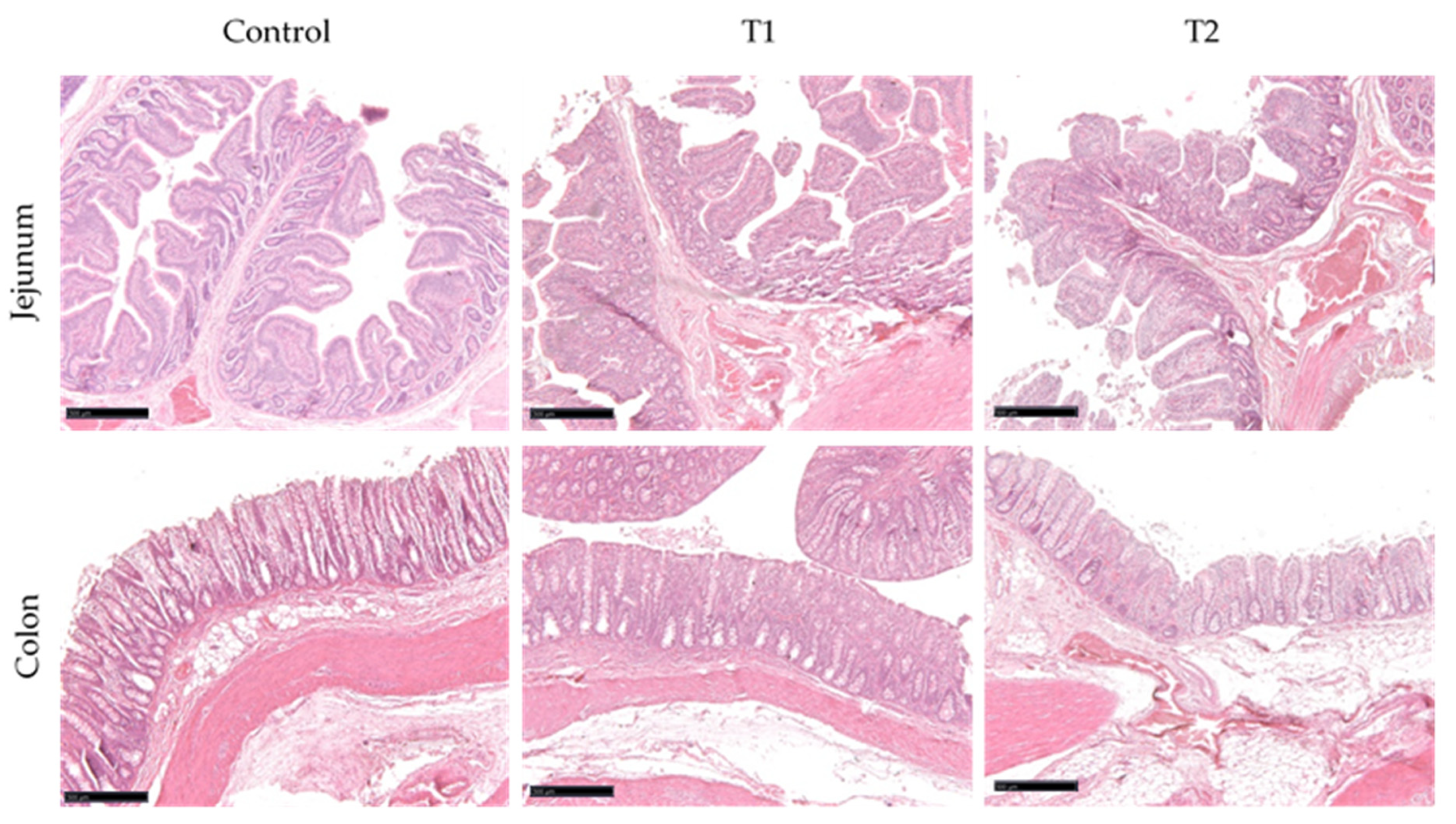
2.1.3. Histological Analysis of Jejunum and Colon
2.1.4. Microbiological Analysis of Fecal Microbiota
2.2. Experiment 2 (Digestibility Trial)
Animals, Experimental Design, and Diets
2.3. Chemical Analysis
2.4. Statistical Analysis
3. Results
3.1. Experiment 1 (Post-Weaning Trial)
3.1.1. Zootechnical Performance
3.1.2. Clinical and Biochemical Parameters
3.1.3. Histological Analysis of Jejunum and Colon
3.1.4. Fecal Microbiota Composition
3.2. Experiment 2 (Digestibility Trial)
4. Discussion
4.1. Post-Weaning Trial
4.2. Digestibility Trial
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Perricone, V.; Comi, M.; Bontempo, V.; Lecchi, C.; Ceciliani, F.; Crestani, M.; Ferrari, A.; Savoini, G.; Agazzi, A. Effects of nucleotides administration on growth performance and immune response of post-weaning piglets. Ital. J. Anim. Sci. 2020, 19, 295–301. [Google Scholar] [CrossRef]
- Heo, J.M.; Opapeju, F.O.; Pluske, J.R.; Kim, J.C.; Hampson, D.J.; Nyachoti, C.M. Gastrointestinal health and function in weaned pigs: A review of feeding strategies to control post-weaning diarrhoea without using in-feed antimicrobial compounds. J. Anim. Physiol. Anim. Nutr. 2013, 97, 207–237. [Google Scholar] [CrossRef]
- Trckova, M.; Lorencova, A.; Babak, V.; Neca, J.; Ciganek, M. The effect of leonardite and lignite on the health of weaned piglets. Res. Vet. Sci. 2018, 119, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Rossi, L.; Vagni, S.; Polidori, C.; Alborali, G.L.; Baldi, A.; Dell’Orto, V. Experimental Induction of Escherichia coli Diarrhoea in Weaned Piglets. Open J. Vet. Med. 2012, 2, 18112. [Google Scholar] [CrossRef] [Green Version]
- López-Gálvez, G.; López-Alonso, M.; Pechova, A.; Mayo, B.; Dierick, N.; Gropp, J. Alternatives to antibiotics and trace elements (copper and zinc) to improve gut health and zootechnical parameters in piglets: A review. Anim. Feed Sci. Technol. 2021, 271, 114727. [Google Scholar] [CrossRef]
- Barton, M.D. Antibiotic use in animal feed and its impact on human healt. Nutr. Res. Rev. 2000, 13, 279–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stein, H.H. Experience of feeding pigs without antibiotics: A European perspective. Anim. Biotechnol. 2002, 13, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Casewell, M.; Friis, C.; Marco, E.; McMullin, P.; Phillips, I. The European ban on growth-promoting antibiotics and emerging consequences for human and animal health. J. Antimicrob. Chemother. 2003, 52, 159–161. [Google Scholar] [CrossRef] [Green Version]
- Van Boeckel, T.P.; Brower, C.; Gilbert, M.; Grenfell, B.T.; Levin, S.A.; Robinson, T.P.; Teillant, A.; Laxminarayan, R. Global trends in antimicrobial use in food animals. Proc. Natl. Acad. Sci. USA 2015, 112, 5649–5654. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Espinosa, C.D.; Abelilla, J.J.; Casas, G.A.; Lagos, L.V.; Lee, S.A.; Kwon, W.B.; Mathai, J.K.; Navarro, D.M.D.L.; Jaworski, N.W.; et al. Non-antibiotic feed additives in diets for pigs: A review. Anim. Nutr. 2018, 4, 113–125. [Google Scholar] [CrossRef]
- Silva-Guillen, Y.V.; Arellano, C.; Boyd, R.D.; Martinez, G.; Van Heugten, E. Growth performance, oxidative stress and immune status of newly weaned pigs fed peroxidized lipids with or without supplemental vitamin e or polyphenols. J. Anim. Sci. Biotechnol. 2020, 11, 22. [Google Scholar] [CrossRef] [PubMed]
- Verhelst, R.; Schroyen, M.; Buys, N.; Niewold, T. Dietary polyphenols reduce diarrhea in enterotoxigenic Escherichia coli (ETEC) infected post-weaning piglets. Livest. Sci. 2014, 160, 138–140. [Google Scholar] [CrossRef]
- Xu, X.; Hua, H.; Wang, L.; He, P.; Zhang, L.; Qin, Q.; Yu, C.; Wang, X.; Zhang, G.; Liu, Y. Holly polyphenols alleviate intestinal inflammation and alter microbiota composition in lipopolysaccharide-challenged pigs. Br. J. Nutr. 2020, 123, 881–891. [Google Scholar] [CrossRef]
- Fiesel, A.; Gessner, D.K.; Most, E.; Eder, K. Effects of dietary polyphenol-rich plant products from grape or hop on pro-inflammatory gene expression in the intestine, nutrient digestibility and faecal microbiota of weaned pigs. BMC Vet. Res. 2014, 10, 196. [Google Scholar] [CrossRef] [Green Version]
- Visscher, C.; Hankel, J.; Nies, A.; Keller, B.; Galvez, E.; Strowig, T.; Keller, C.; Breves, G. Performance, fermentation characteristics and composition of the microbiome in the digest of piglets kept on a feed with humic acid-rich peat. Front. Vet. Sci. 2019, 6, 29. [Google Scholar] [CrossRef]
- Wang, Q.; Chen, Y.J.; Yoo, J.S.; Kim, H.J.; Cho, J.H.; Kim, I.H. Effects of supplemental humic substances on growth performance, blood characteristics and meat quality in finishing pigs. Livest. Sci. 2008, 117, 270–274. [Google Scholar] [CrossRef]
- Ji, F.; McGlone, J.J.; Kim, S.W. Effects of dietary humic substances on pig growth performance, carcass characteristics, and ammonia emission. J. Anim. Sci. 2006, 84, 2482–2490. [Google Scholar] [CrossRef] [Green Version]
- Guilloteau, P.; Martin, L.; Eeckhaut, V.; Ducatelle, R.; Zabielski, R.; Van Immerseel, F. From the gut to the peripheral tissues: The multiple effects of butyrate. Nutr. Res. Rev. 2010, 23, 366–384. [Google Scholar] [CrossRef] [Green Version]
- Lu, H.; Su, S.; Ajuwon, K.M. Butyrate supplementation to gestating sows and piglets induces muscle and adipose tissue oxidative genes and improves growth performance. J. Anim. Sci. 2012, 90, 430–432. [Google Scholar] [CrossRef] [Green Version]
- Miragoli, F.; Patrone, V.; Prandini, A.; Sigolo, S.; Dell’anno, M.; Rossi, L.; Senizza, A.; Morelli, L.; Callegari, M.L. Implications of tributyrin on gut microbiota shifts related to performances of weaning piglets. Microorganisms 2021, 9, 584. [Google Scholar] [CrossRef]
- Sotira, S.; Dell’Anno, M.; Caprarulo, V.; Hejna, M.; Pirrone, F.; Callegari, M.L.; Tucci, T.V.; Rossi, L. Effects of tributyrin supplementation on growth performance, insulin, blood metabolites and gut microbiota in weaned piglets. Animals 2020, 10, 726. [Google Scholar] [CrossRef]
- Nochta, I.; Tuboly, T.; Halas, V.; Babinszky, L. Effect of different levels of mannan-oligosaccharide supplementation on some immunological variables in weaned piglets. J. Anim. Physiol. Anim. Nutr. 2009, 93, 496–504. [Google Scholar] [CrossRef] [PubMed]
- Halas, V.; Nochta, I. Mannan oligosaccharides in nursery pig nutrition and their potential mode of action. Animals 2012, 2, 261–274. [Google Scholar] [CrossRef] [Green Version]
- Spring, P.; Wenk, C.; Connolly, A.; Kiers, A. A review of 733 published trials on Bio-Mos®, a mannan oligosaccharide, and Actigen®, a second generation mannose rich fraction, on farm and companion animals. J. Appl. Anim. Nutr. 2015, 3, E8. [Google Scholar] [CrossRef]
- Newman, K.E.; Newman, M.C. Evaluation of mannan oligosaccharide on the microflora and immunoglobulin status of sows and piglet performance. J. Anim. Sci. 2001, 79, 189. [Google Scholar]
- Spring, P.; Wenk, C.; Dawson, K.A.; Newman, K.E. The Effects of Dietary Mannanoligosaccharides on Cecal Parameters and the Concentrations of Enteric Bacteria in the Ceca of Salmonella-Challenged Broiler Chicks. Poult. Sci. 2000, 79, 205–211. [Google Scholar] [CrossRef]
- Shi, H.; Kim, I.H. Dietary yeast extract complex supplementation increases growth performance and nutrient digestibility of weaning pigs. Livest. Sci. 2019, 230, 103850. [Google Scholar] [CrossRef]
- Wu, Y.; Pan, L.; Tian, Q.; Piao, X. Comparative digestibility of energy and ileal amino acids in yeast extract and spray-dried porcine plasma fed to pigs. Arch. Anim. Nutr. 2018, 72, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.; Tian, Q.; Wu, Y.; Long, S.; Yin, J.; Piao, X. Yeast extract could be used as a partial substitute for spray-dried porcine plasma in diets for weaned pigs. Livest. Sci. 2019, 224, 20–25. [Google Scholar] [CrossRef]
- Patterson, R.; Heo, J.M.; Wickramasuriya, S.S.; Yi, Y.J.; Nyachoti, C.M. Dietary nucleotide rich yeast extract mitigated symptoms of colibacillosis in weaned pigs challenged with an enterotoxigenic strain of Escherichia coli. Anim. Feed Sci. Technol. 2019, 254, 114204. [Google Scholar] [CrossRef]
- Waititu, S.M.; Yin, F.; Patterson, R.; Yitbarek, A.; Rodriguez-Lecompte, J.C.; Nyachoti, C.M. Dietary supplementation with a nucleotide-rich yeast extract modulates gut immune response and microflora in weaned pigs in response to a sanitary challenge. Animal 2017, 11, 2156–2164. [Google Scholar] [CrossRef]
- National Research Council. Nutrient Requirements of Swine, 11th ed.; National Academies Press: Washington, DC, USA, 2012; ISBN 978-0-309-22423-9. [Google Scholar]
- Abeni, F.; Petrera, F.; Dal Prà, A.; Rapetti, L.; Malagutti, L.; Galassi, G. Blood parameters in fattening pigs fed whole-ear corn silage and housed in group pens or in metabolic cages. J. Anim. Sci. 2015, 93, 3901–3908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abeni, F.; Petrera, F.; Prà, A.D.; Rapetti, L.; Crovetto, G.M.; Galassi, G. Blood parameters in fattening pigs from two genetic types fed diet with three different protein concentrations. Transl. Anim. Sci. 2018, 2, 372–382. [Google Scholar] [CrossRef] [PubMed]
- Boone, L.; Meyer, D.; Cusick, P.; Ennulat, D.; Provencher Bolliger, A.; Everds, N.; Meador, V.; Elliott, G.; Honor, D.; Bounous, D.; et al. Selection and interpretation of clinical pathology indicators of hepatic injury in preclinical studies. Vet. Clin. Pathol. 2005, 34, 182–188. [Google Scholar] [CrossRef]
- Magistrelli, D.; Zanchi, R.; Malagutti, L.; Galassi, G.; Canzi, E.; Rosi, F. Effects of Cocoa Husk Feeding on the Composition of Swine Intestinal Microbiota. J. Agric. Food Chem. 2016, 64, 2046–2052. [Google Scholar] [CrossRef] [PubMed]
- Arlington, V. AOAC Official Methods of Analysis of Official Analytical Chemist; AOAC International: Gaithersburg, MD, USA, 1995. [Google Scholar]
- Mertens, D.R. Gravimetric determination of amylase-treated neutral detergent fiber in feeds with refluxing in beakers or crucibles: Collaborative study. J. AOAC Int. 2002, 85, 1217–1240. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for Dietary Fiber, Neutral Detergent Fiber, and Nonstarch Polysaccharides in Relation to Animal Nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- SAS Institute Inc. SAS® 9.4 Statements: Reference; SAS Institute Inc.: Cary, NC, USA, 2013. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.M.; Walker, S.C. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67. [Google Scholar] [CrossRef]
- Rychen, G.; Aquilina, G.; Azimonti, G.; Bampidis, V.; Bastos, M.; de Lourdes Bastos, M.; Bories, G.; Chesson, A.; Cocconcelli, P.S.; Flachowsky, G.; et al. Guidance on the assessment of the efficacy of feed additives. EFSA J. 2018, 16, e05274. [Google Scholar] [CrossRef] [Green Version]
- Stensland, I.; Kim, J.C.; Bowring, B.; Collins, A.M.; Mansfield, J.P.; Pluske, J.R. A comparison of diets supplemented with a feed additive containing organic acids, cinnamaldehyde and a permeabilizing complex, or zinc oxide, on post-weaning diarrhoea, selected bacterial populations, blood measures and performance in weaned pigs experime. Animals 2015, 5, 1147–1168. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.S.; Xiong, X.; Li, J.Z.; Yin, Y.L. Effects of chito-oligosaccharide on intestinal mucosal amino acid profiles and alkaline phosphatase activities, and serum biochemical variables in weaned piglets. Livest. Sci. 2016, 190, 141–146. [Google Scholar] [CrossRef]
- Shu, Y.; Yu, B.; He, J.; Yu, J.; Zheng, P.; Yuan, Z.; Chen, D.; Mao, X. Excess of dietary benzoic acid supplementation leads to growth retardation, hematological abnormality and organ injury of piglets. Livest. Sci. 2016, 190, 94–103. [Google Scholar] [CrossRef]
- Pluske, J.R.; Williams, I.H.; Aherne, F.X. Maintenance of villous height and crypt depth in piglets by providing continuous nutrition after weaning. Anim. Sci. 1996, 62, 131–144. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y. Fatty acids, inflammation and intestinal health in pigs. J. Anim. Sci. Biotechnol. 2015, 6, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verdile, N.; Mirmahmoudi, R.; Brevini, T.A.L.; Gandolfi, F. Evolution of pig intestinal stem cells from birth to weaning. Animal 2019, 13, 2830–2839. [Google Scholar] [CrossRef]
- Leitch, E.C.M.W.; Walker, A.W.; Duncan, S.H.; Holtrop, G.; Flint, H.J. Selective colonization of insoluble substrates by human faecal bacteria. Environ. Microbiol. 2007, 9, 667–679. [Google Scholar] [CrossRef]
- Ze, X.; Duncan, S.H.; Louis, P.; Flint, H.J. Ruminococcus bromii is a keystone species for the degradation of resistant starch in the human colon. ISME J. 2012, 6, 1535–1543. [Google Scholar] [CrossRef]
- Topping, D.L.; Clifton, P.M. Short-chain fatty acids and human colonic function: Roles of resistant starch and nonstarch polysaccharides. Physiol. Rev. 2001, 81, 1031–1064. [Google Scholar] [CrossRef]
- Nugent, A.P. Health properties of resistant starch. Nutr. Bull. 2005, 30, 27–54. [Google Scholar] [CrossRef]
- Ze, X.; Le Mougen, F.; Duncan, S.H.; Louis, P.; Flint, H.J. Some are more equal than others: The role of “keystone” species in the degradation of recalcitrant substrates. Gut Microbes 2013, 4, 236–240. [Google Scholar] [CrossRef] [Green Version]
- Pryde, S.E.; Duncan, S.H.; Hold, G.L.; Stewart, C.S.; Flint, H.J. The microbiology of butyrate formation in the human colon. FEMS Microbiol. Lett. 2002, 217, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Louis, P.; Scott, K.P.; Duncan, S.H.; Flint, H.J. Understanding the effects of diet on bacterial metabolism in the large intestine. J. Appl. Microbiol. 2007, 102, 1197–1208. [Google Scholar] [CrossRef] [PubMed]
- Schiavon, S.; Carraro, L.; Dalla Bona, M.; Cesaro, G.; Carnier, P.; Tagliapietra, F.; Sturaro, E.; Galassi, G.; Malagutti, L.; Trevisi, E.; et al. Growth performance, and carcass and raw ham quality of crossbred heavy pigs from four genetic groups fed low protein diets for dry-cured ham production. Anim. Feed Sci. Technol. 2015, 208, 170–181. [Google Scholar] [CrossRef]
- Il Consorzio | Consorzio del Prosciutto di Parma. Available online: https://www.prosciuttodiparma.com/il-consorzio/ (accessed on 20 July 2021).
- Documenti-Prosciutto di San Daniele. Available online: https://prosciuttosandaniele.it/il-consorzio/documenti/ (accessed on 20 July 2021).


{kind=link}
| Item | Experiment 1 | Experiment 2 |
|---|---|---|
| Ingredients (g/kg DM 1) | ||
| Corn meal | 295 | 320 |
| Barley meal | 156 | 225 |
| Wheat meal | 172 | 93.0 |
| Wheat bran | 74.0 | 59.0 |
| Beet pulp | 29.0 | 24.5 |
| Soybean hulls | 7.0 | 0 |
| Soybean meal CP 2 48% | 49.0 | 116 |
| Soybean seeds flaked | 38.0 | 9.0 |
| Soybean protein concentrate | 38.0 | 29.0 |
| Oils | 29.0 | 25.3 |
| Milk whey | 27.0 | 32.8 |
| Lactose | 16.5 | 0 |
| Fish meal | 14.3 | 10.0 |
| CaHPO4 | 4.8 | 6.3 |
| NaCl | 4.1 | 3.5 |
| CaSO4 | 2.9 | 5.1 |
| Lysine | 5.6 | 4.8 |
| Threonine | 2.4 | 1.8 |
| Methionine | 2.1 | 1.6 |
| Vitamin-mineral mix 3 | 33.3 | 33.3 |
| Chemical analysis (g/kg DM) | ||
| CP 2 | 183 | 182 |
| EE 4 | 65.8 | 58.0 |
| NDF 5 | 124 | 131 |
| ADF 6 | 50.5 | 56.3 |
| Ash | 54.0 | 55.8 |
| Lysine | 13.9 | 13.1 |
| Methionine | 5.21 | 4.55 |
| GE 7 (MJ/kg DM) | 19.04 | 18.67 |
| Item | Diet 1 | SE | p | ||
|---|---|---|---|---|---|
| C | T1 | T2 | |||
| Initial BW (kg) | 8.9 | 9.0 | 9.1 | 0.286 | 0.921 |
| Final BW (kg) | 39.8 | 38.7 | 38.2 | 1.17 | 0.678 |
| ADG (kg) | 0.717 | 0.690 | 0.679 | 0.025 | 0.554 |
| Feed intake (kg/d) | 4.27 | 4.13 | 4.10 | 0.253 | 0.776 |
| FCR (kg/kg) | 1.492 | 1.496 | 1.505 | 0.022 | 0.853 |
| Item | Diet 1 | SE | p | ||
|---|---|---|---|---|---|
| C | T1 | T2 | |||
| Energy and lipid metabolism | |||||
| Glucose, mmol/L | 8.14 | 8.75 | 7.01 | 0.537 | 0.078 |
| Total cholesterol, mmol/L | 2.88 | 2.89 | 2.65 | 0.114 | 0.234 |
| Triglycerides, mmol/L | 0.755 | 0.604 | 0.701 | 0.170 | 0.809 |
| NEFA 2, mmol/L | 0.177 | 0.176 | 0.234 | 0.036 | 0.428 |
| BHB 3, mmol/L | 0.073 | 0.012 | 0.036 | 0.032 | 0.403 |
| Nitrogen and protein metabolism | |||||
| Urea, mmol/L | 3.92 ab | 5.09 a | 3.61 b | 0.396 | 0.021 |
| Creatinine, µmol/L | 127 | 150 | 123 | 13.2 | 0.280 |
| Total protein, g/L | 65.9 | 68.0 | 64.9 | 1.39 | 0.270 |
| Albumin, g/L | 37.7 | 40.2 | 40.1 | 1.24 | 0.282 |
| Globulins, g/L | 28.2 | 27.8 | 24.8 | 1.71 | 0.319 |
| Albumin: Globulins | 1.42 | 1.48 | 1.69 | 0.116 | 0.238 |
| Minerals | |||||
| Ca, mmol/L | 3.10 | 3.09 | 2.99 | 0.058 | 0.341 |
| P, mmol/L | 3.59 | 3.81 | 3.46 | 0.139 | 0.191 |
| Mg, mmol/L | 1.06 | 1.13 | 1.09 | 0.092 | 0.850 |
| Na, mmol/L | 151 | 151 | 150 | 1.33 | 0.772 |
| K, mmol/L | 7.46 | 8.53 | 7.72 | 0.500 | 0.278 |
| Cl, mmol/L | 106 | 105 | 106 | 0.984 | 0.760 |
| Fe, µmol/L | 34.2 | 35.8 | 30.9 | 2.60 | 0.388 |
| Enzyme activities and bilirubin | |||||
| Amylase, U/L | 1784 | 2370 | 2321 | 229 | 0.146 |
| ALT 4, U/L | 75.9 | 80.6 | 80.0 | 5.02 | 0.766 |
| AST 5, U/L | 78.1 b | 162 a | 96.6 ab | 20.2 | 0.013 |
| GGT 6, U/L | 27.3 | 31.4 | 29.0 | 3.59 | 0.714 |
| ALP 7, U/L | 276 | 270 | 256 | 13.1 | 0.564 |
| TRAP 8, U/L | 15.8 | 28.1 | 19.2 | 5.74 | 0.289 |
| LDH 9, U/L | 1498 ab | 1822 a | 1390 b | 107 | 0.017 |
| Total bilirubin, μmol/L | 0.410 | 0.309 | 0.342 | 0.113 | 0.808 |
| Item | Diet 1 | SE | p | ||
|---|---|---|---|---|---|
| C | T1 | T2 | |||
| Jejunum | |||||
| Villus length (μm) | 1288 | 1257 | 1214 | 228 | 0.935 |
| Crypt depth (μm) | 217 | 195 | 210 | 17.4 | 0.465 |
| Villus/Crypt ratio (V:C) | 5.93 | 6.46 | 5.78 | 1.38 | 0.940 |
| Colon | |||||
| Crypt depth (μm) | 290 | 273 | 303 | 17.3 | 0.293 |
| Domain/Genus/Species | Diet 1 | SE | p | |
|---|---|---|---|---|
| C | T2 | |||
| Eubacteria | 10.8 | 11.0 | 0.123 | 0.395 |
| Bacteroides–Prevotella | 9.07 | 9.15 | 0.183 | 0.833 |
| Bifidobacterium | 8.71 | 8.73 | 0.176 | 0.837 |
| Clostridium histolyticum | 8.97 | 9.06 | 0.149 | 0.782 |
| Clostridium coccoides–Eubacterium rectale | 9.63 | 9.77 | 0.269 | 0.803 |
| Escherichia coli | 8.31 | 8.38 | 0.074 | 0.632 |
| Faecalibacterium prausnitzii | 9.48 | 9.59 | 0.218 | 0.821 |
| Lactobacillus–Enterococcus | 8.77 | 9.28 | 0.167 | 0.100 |
| Ruminococcus | 8.28 | 9.09 | 0.234 | 0.022 |
| Item | Diet 1 | SE | p | ||
|---|---|---|---|---|---|
| C | T1 | T2 | |||
| DM 2 | 86.0 b | 88.9 a | 89.3 a | 0.888 | 0.026 |
| OM 3 | 87.6 b | 90.2 a | 90.5 a | 0.797 | 0.030 |
| CP 4 | 87.0 b | 90.2 a | 90.0 a | 0.939 | 0.040 |
| EE 5 | 82.7 b | 87.3 a | 85.8 a | 1.05 | 0.017 |
| NDF 6 | 45.7 | 53.6 | 57.7 | 3.59 | 0.059 |
| ADF 7 | 37.2 | 38.3 | 43.2 | 4.20 | 0.124 |
| Ash | 58.1 b | 67.6 a | 68.6 a | 2.70 | 0.019 |
| GE 8 | 86.2 b | 89.1 a | 89.5 a | 0.873 | 0.024 |
| Item | Diet 1 | SE | p | ||
|---|---|---|---|---|---|
| C | T1 | T2 | |||
| N balance, g/d | |||||
| Intake (NI) | 57.9 b | 64.8 a | 58.2 b | 0.979 | <0.001 |
| Fecal | 7.53 | 6.37 | 5.81 | 0.558 | 0.085 |
| Urinary | 28.9 | 30.1 | 27.0 | 1.79 | 0.436 |
| Retained | 21.5 b | 28.3 a | 25.4 ab | 1.70 | 0.031 |
| N balance, g/BW0.75 | |||||
| Intake | 2.30 b | 2.36 b | 2.46 a | 0.031 | 0.004 |
| Fecal | 0.297 | 0.232 | 0.247 | 0.021 | 0.095 |
| Urinary | 1.14 | 1.10 | 1.14 | 0.056 | 0.820 |
| Retained | 0.864 | 1.03 | 1.07 | 0.081 | 0.141 |
| N balance, % NI | |||||
| Fecal | 13.0 a | 9.83 b | 10.0 b | 0.939 | 0.040 |
| Urinary | 49.7 | 46.4 | 46.4 | 2.62 | 0.559 |
| Retained | 37.3 | 43.7 | 43.6 | 2.77 | 0.221 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Galassi, G.; Battelli, M.; Verdile, N.; Rapetti, L.; Zanchi, R.; Arcuri, S.; Petrera, F.; Abeni, F.; Crovetto, G.M. Effect of a Polyphenol-Based Additive in Pig Diets in the Early Stages of Growth. Animals 2021, 11, 3241. https://doi.org/10.3390/ani11113241
Galassi G, Battelli M, Verdile N, Rapetti L, Zanchi R, Arcuri S, Petrera F, Abeni F, Crovetto GM. Effect of a Polyphenol-Based Additive in Pig Diets in the Early Stages of Growth. Animals. 2021; 11(11):3241. https://doi.org/10.3390/ani11113241
Chicago/Turabian StyleGalassi, Gianluca, Marco Battelli, Nicole Verdile, Luca Rapetti, Raffaella Zanchi, Sharon Arcuri, Francesca Petrera, Fabio Abeni, and Gianni Matteo Crovetto. 2021. "Effect of a Polyphenol-Based Additive in Pig Diets in the Early Stages of Growth" Animals 11, no. 11: 3241. https://doi.org/10.3390/ani11113241