1. Introduction
High dietary crude fiber (CF) levels usually had a negative impact on feed utilization, such as anti-nutritional factors and the decrease of nutrient digestibility, which was related to the concentration and composition of CF, processing technology, and the animal itself [
1]. However, because of a good tolerance to fibrous feed, the growth rate and nutrient digestibility of Chinese local pig breeds did not change after pigs were fed diets with 7.57% or 9.7% CF [
2,
3], and finishing pigs could digest and utilize CF better than piglets [
4,
5]. Although CF was not called a nutrient and had a certain anti-nutritional effect, it still had nutritional value, especially for hindgut health and intestinal microbes [
6]. CF had the benefit of promoting intestine microbiota fermentation in animal, and feeding diets with high fiber levels could increase the relative abundance of probiotic bacteria and promote SCFAs production, which was beneficial to intestinal health and microbial balance [
7,
8]. Therefore, the research focus on how to avoid the negative effects of CF on nutrition and to better exert its probiotic effects.
Solid-state fermentation referred to the fermentation by microorganisms in a solid substrate with little free water. Cellulase and xylanase produced via this process could degrade CF in feed and promote the digestion and utilization of CF [
9]. In addition, studies had shown that the role of probiotics had genus, species, and strain specificity, and the multi-bacteria fermentation with synergy would be more effective than a single probiotic [
10,
11]. For example, as aerobic bacteria,
Bacillus subtilis and
Candida utilis were able to consume oxygen, create an anaerobic environment for
Lactobacillus amylovorus and
Enterococcus faecalis, and promote their growth. Besides,
Lactobacillus amylovorus could produce lactic acid to reduce the pH of fermented feed, inhibit the growth and reproduction of Gram-negative pathogens, reduce body serum cholesterol, ammonia nitrogen, the amount of hemicellulose and lignin in fermented feed, and increase soluble carbohydrate content [
12].
Enterococcus faecalis could produce lactic acid and SCFAs and regulate lipid metabolism. Its metabolized hydrogen peroxide and mannan peptide could kill harmful bacteria such as
Escherichia coli and
Salmonella [
13,
14].
Bacillus subtilis was used to consume free oxygen, to create a favorable environment for anaerobic probiotics such as
Lactobacillus, and to inhibit the growth of harmful bacteria to adjust the digestive tract flora [
15].
Bacillus subtilis secreted a variety of digestive enzymes for proteins and starch, and eliminated anti-nutritional factors as well [
16].
Candida utilis was an excellent source of amino acids, nucleic acids, vitamins, and a variety of digestive enzymes that are beneficial to animals (α-amylase, protease, cellulase, etc.), benefiting animal digestion [
17]. The cell wall of
Candida utilis contained dextran, mannan, and other active substances, which had been proven to increase the expression of Occludin and β-definsin-2, promote the development of the intestinal barrier, and enhance animal immune function [
18,
19].
To study multi-bacteria solid-state fermentation, improving high dietary CF levels’ adverse effect on the nutrient digestibility, the Lactobacillus amylovorus TZR-PI001, Enterococcus faecalis TZR-PI003, Bacillus subtilis ATCC 19659, and Candida utilis CICC 1314 strains were applied in solid-state fermentation diets with dietary 2.52% or 7.00% CF levels.
2. Materials and Methods
2.1. Strain, Culture Media, and Reagents
Lactobacillus amylovorus TZR-PI001 and Enterococcus faecalis TZR-PI003 were provided by the Applied Microbiology Laboratory of the College of Animal Science and Technology of Southwest University. Bacillus subtilis ATCC 19659 was purchased from American Type Culture Collection (ATCC). Candida utilis CICC 1314 was purchased from the China Center of Industrial Culture Collection (CICC).
MRS culture medium comprised 10.0 g beef extract, 10.0 g peptone, 5.0 g sodium acetate, 5.0 g yeast powder, 20.0 g glucose, 2.0 g diammonium citrate, 1.0 g Tween 80, 2.0 g K2HPO4, 0.58 g MgSO4·7H2O, 0.25 g MnSO4·H2O, 15.0 g agar, and 1.0 L distilled water. This medium was adjusted to pH 6.8 and sterilized for 15 min at 115 °C.
TSA medium comprised 15.0 g tryptone, 5.0 g peptone, 5.0 g NaCl, 13.0 g agar, and 1.0 L distilled water. This medium was adjusted to pH 7.3 and sterilized for 15 min at 115 °C.
YPD culture medium comprised 20.0 g glucose, 10.0 g yeast powder, 20.0 g peptone, 15.0 g agar, and 1.0 L distilled water. This medium was adjusted to pH 6.0 and sterilized for 15 min at 115 °C.
Detection kits of serum total cholesterol (TC) (CV ≤ 3%), total triglycerides (TG) (CV ≤ 5%), high-density lipoprotein (HDL-c) (CV ≤ 3%), low-density lipoprotein (LDL-c) (CV ≤ 8%), total protein (TP) (CV = 1.03%), albumin (ALB) (CV = 2.3%), urea nitrogen (BUN) (CV ≤ 5%), glucose (GLU) (CV ≤ 5%), total antioxidant capacity (T-AOC) (CV ≤ 3.6%), glutathione peroxidase (GSH-Px) (CV ≤ 3.56%), superoxide dismutase (SOD) (CV ≤ 1.7%), catalase (CAT) (CV ≤ 1.7%), and malondialdehyde (MDA) (CV = 1.5%) were purchased from Nanjing Jiancheng Bioengineering Institute (Nanjing, China).
2.2. The Preparation of Fermented Basal Diet
The Lactobacillus amylovorus TZR-PI001, Enterococcus faecalis TZR-PI003, Bacillus subtilis ATCC 19659, and Candida utilis CICC 1314 strains were taken out from the −80 °C refrigerator and activated on solid culture medium. After purification twice, they were respectively inoculated into a liquid culture medium and adjusted to a concentration of 108 CFU/mL to prepare the seed culture solution. L. amylovorus and E. faecalis used MRS culture medium, B. subtilis used TSA culture medium, and C. utilis used YPD culture medium. L. amylovorus, E. faecalis, B. subtilis, and C. utilis were mixed in a ratio of 2:1:1:1 to prepare a mixed bacterial liquid. When making fermented feed, 1 kg of complete feed was added to 21 mL of mixed bacteria liquid and 540 mL of water, and then the uniformly mixed fermented feed was put into a fermentation bag, sealed, and fermented at 30 °C for 3.5 days. The fermented feed was stirred evenly before feeding.
2.3. Feeding Trial
2.3.1. Experimental Design, Animals, and Diets
The experiment was established according to a 2 (factors) × 2 (levels) design with multi-bacteria fermentation (fed a non-fermented diet or multi-bacteria fermentation) or CF level (fed basal diet containing 2.52% CF or 7.00% CF). A total of 36 finishing pigs (Landrace × Large White × Duroc) with a similar initial body weight (70.80 ± 5.75 kg) were randomly divided into 4 treatments of 9 barrows per group: (1) pigs fed a basal diet containing 7.00% CF (HF), (2) pigs fed a basal multi-bacteria fermentation diet containing 7.00% CF (HFM), (3) pigs fed a basal diet containing 2.52% CF (LF), and (4) piglets fed a basal multi-bacteria fermentation diet containing 2.52% CF (LFM). The ingredients and the compositions of the diet were formulated according to NRC (2012) recommendations (
Table 1).
2.3.2. Feeding Management and Sample Collection
The experimental period lasted 28 days. Each pig was kept in 300 m2 rooms with individual pens (1.5 m length × 0.5 m width × 0.8 m height) and temperature-controlled (30 ± 1.2 °C), and fed at 08:00 and 18:00. Food and water were provided ad libitum.
Feed intake of pigs was recorded daily. The pigs without intake feed were weighed on day 0 and day 28. The pigs were measured for the backfat thickness, body length, body width, and chest circumference on day 28. On the morning of day 29, a total of 6 pigs without intake feed were selected from each group and a 10 mL blood sample was collected. The blood sample was kept for 60 min and centrifuged at 3500 × g for 10 min at 4 °C to harvest the serum. Serum was stored at −80 °C for biochemical analysis. Five days before the end of the test, six pigs were randomly selected collected rectal feces sample using sterile rectal swabs for three days, and stored at −80 °C for 16S rRNA gene sequencing. The License of Experimental Animals (SYXK 2014-0002) of the Animal Experimentation Ethics Committee of Southwest University (Chongqing, China) approved all experimental procedures.
2.3.3. Biochemical Analysis
The presence of TC, TG, HDL-c, LDL-c, TP, ALB, BUN, GLU, T-AOC, GSH-Px, SOD, CAT, and MDA in serum was determined using colorimetric methods with a reagent kit according to the manufacturer’s instructions (Nanjing Jiancheng Institute of Bioengineering, Nanjing, Jiangsu, China).
2.3.4. The 16S rRNA Gene Sequencing Analysis
According to the introduction of the Power Fecal DNA Isolation Kit (Mobio, USA), the DNA samples were extracted from samples of feces, stained using the Quant-iT Pico Green dsDNA Kit (Invitrogen Ltd., Paisley, UK), and quantified using a Nanodrop spectrophotometer (Nyxor Biotech, Paris, France). This experiment used the Illumina platform to perform paired-end sequencing of community DNA fragments, and the Greengenes database was selected. According to the QIIME2 DADA2 analysis process and Vsearch software analysis process, we performed sequence denoising or OTU clustering and output the ASV/OTU table, and finally removed the singleton ASVs/OTUs in the table. Through statistics on the ASV/OTU table, the specific composition information at each classification level of the microbial community in each sample was obtained. On this basis, different leveling depths were selected to calculate the alpha diversity index of the microbial community, including Chao1 and the observed species index representing richness, Shannon and Simpson index representing diversity, Faith’s PD index representing evolution-based diversity, Pielou’s evenness index representing evenness, and Good’s coverage index representing coverage. At the ASV/OTU level, we calculated the distance matrix of each sample and used a variety of unsupervised sorting and clustering methods, combined with corresponding statistical testing methods, to measure the difference in beta diversity between different groups and the significance of the difference. We also measured the differences in species’ abundance composition between different groups and searched for symbolic species at the species taxonomic composition level. Then, based on the 16S rRNA gene sequencing results, PICRUSt2 (Phylogenetic Investigation of Communities by Reconstruction of Unobserved States) software was used to analyze the abundance of marker gene sequences in the sample to predict the functional abundance of the sample. After obtaining the functional unit, the data were processed according to the MetaCyc Pathway Database to predict the metabolic function of the sample flora and find the differential pathways.
2.4. Digestion Trial
2.4.1. Experimental Design, Treatment, and Sample Collection
A total of 5 test pigs were randomly selected from each group and kept in 30 m2 rooms, with 4 mechanically ventilated on the 5th day before the end of the trial period. One g/kg of titanium dioxide (TiO2) was added to their feed in advance, and their daily feces and feed were collected through the incomplete feces collection method for three days. To every 100 g feces, 10 mL of 10% H2SO4 was added, then stored at 4 °C for later use after being stirred evenly.
2.4.2. Chemical Analysis
Samples of diets and feces were analyzed for the content of DM (Method 930.15; AOAC Int., 2007), CP (Method 990.03; AOAC Int., 2007), EE (ISO 6492: 1999), CF (ISO 6865: 2000), NDF (GB/T 20806-2006), ADF (NY/T 1459-2007), Ca (GB 5009.92-2016), total P(GB 5009.87-2016), and amino acids (Hitachi L-8800 automatic amino acid analyzer). All diets and feces samples were analyzed for the content of TiO
2 using the method of Schürch et al. [
20].
2.5. Data Calculation Formula and Statistical Analysis
The average daily weight gain, the average daily feed intake, and the ratio of feed to gain of piglets were calculated according to the following formula:
The apparent total tract digestibility (ATTD) of CP, CF, ADF, NDF, EE, Ca, total P, and amino acids was calculated according to the following equation:
where AD
nutrient is the ATTD of a nutrient in the diet (%), Nutrient
(diet) and Nutrient
(feces) are the nutrient concentration in the diet and the feces samples respectively, and TiO
2(diet) and TiO
2(feces) are the TiO
2 concentration (g/kg) in the diet and the feces samples, respectively.
The total tract digestion amount of CP, CF, EE, Ca, totalP, and amino acids was calculated according to the following formula:
Data were analyzed according to two-way analysis of variance with CF levels (2 levels) and multi-bacteria solid-state fermentation (2 levels) using the GLM procedure (SAS Institute 222 Inc.; Cary, NC, USA). The values presented in the tables represent means and pooled standard error of the means (SEMs). The Student-Neuman-Keuls (SNK) test was performed to identify differences among groups. Significance was set at p < 0.05.
4. Discussion
Multi-bacteria fermentation and dietary CF levels had little effect on the weight gain of finishing pigs in this study. The effects of crude fiber and fermented feed on growth performance varied in different experiments. Feeding
Bacillus fully fermented feed significantly increased the ADG of weaned piglets and reduced the F:G [
21], while adding fermented grain feed or flaxseed meal to piglets’ diet did not significantly affect growth performance and ATTD in other trials [
22,
23].
Dietary CF levels showed the most important impact in this study. Compared with the 2.52% CF diet, the 7.00% diet significantly reduced the ATTD and digestion amount of CP, CF, NDF, ADF, EE, and amino acids, indicating that the 7.00% CF level exceeded the tolerance of the finishing pigs in this experiment. Excess crude fiber would increase the flow of the digestive tract, and the soluble fiber with high water-holding capacity wrapped the chyme and caused insufficient contact between the contents and the intestinal wall, resulting in a reduction of nutrient digestibility [
24]. This result was consistent with the results of Chen et al. [
25], in whose study the CF level of 10% alfalfa (four CF levels were set at 0%, 5%, 10%, and 20%) was similar to our experiment and the total tract digestibility of CP, DM, ADF, and NDF in the 10% alfalfa diet group was significantly lower than that in the 0% and 5% group. Asosta et al. [
26] applied distillers dried grains with soluble (DDGS) in a gradient, and the apparent ileal digestibility (AID) of each amino acid was reduced to varying degrees, similar to the results of this study. The optimum fiber tolerance of finishing pigs in our experiment was less than 7%, and the follow-up experiment continued to explore this. Solid-state fermentation was able to improve the nutrient digestion of CP, CF, NDF, ADF, EAA, NEAA, and TAA, and alleviate the side effect of high fiber on the nutrients.
Enterococcus faecalis is a common lactic acid bacteria. Solid-state fermentation with
Enterococcus faecalis was able to reduce pH, destroy plant cell walls, and increase the content of soluble dietary fiber [
27].
Bacillus subtilis were capable of secreting xylanase and
β-mannanase, decomposing plant hemicellulose, and reducing the content of non-starch polysaccharides (NSP) and anti-nutritional factors in grain feeds [
28,
29].
Candida utilis could decompose lignin and cellulose into monosaccharides and ferment them under aerobic conditions as well.
Candida utilis was particularly efficient in metabolizing hexoses [
30]. Solid-state fermentation could promote nutrient digestion and utilization of crude fiber.
Usually, high-fiber diets can reduce blood sugar, blood lipids, and cholesterol [
31,
32], and the fermentability of crude fiber leads to an increase in the production of colonic short-chain fatty acids (SCFA), including the increase of the ratio of propionate and butyrate and the reduction of the ratio of acetate [
33]. Acetate participates in de novo lipogenesis through the conversion to acetyl-CoA, while propionate participates in gluconeogenesis and was regarded as an inhibitor of fat formation. Butyrate plays an important role in maintaining the integrity of the mucosa and controlling inflammation [
34]. Besides, SCFAs affected the activity of metabolic hormones such as leptin, insulin, and glucagon [
35]. However, the serum TC, TG, and LDL of the high-fiber diet group in this experiment were significantly higher than those of the low-fiber diet group, which contradicted the above conclusions and might be due to a metabolic disorder caused by a too-high CF level. Previous studies found that there was a strong negative correlation between branched-chain amino acids and TC [
36], and the ATTD of leucine, isoleucine, and valine in the high-fiber diet group was significantly lower than that in the low-fiber diet group in this experiment, which was consistent with this finding. High-fiber diets increased the content of serum BUN, and decreased the ATTD of glutamic acid, arginine, proline, and aspartic acid, which are involved in the urea cycle. These results indicated that a high-fiber diet of up to 7.00% could affect the urea cycle and nitrogen balance in finishing pigs and reduce the utilization of nitrogen. However, solid-state fermentation was able to alleviate the adverse effects of high-fiber diets in many ways, such as reducing the content of serum TC and TG, increasing antioxidant capacity, and improving the utilization of nitrogen.
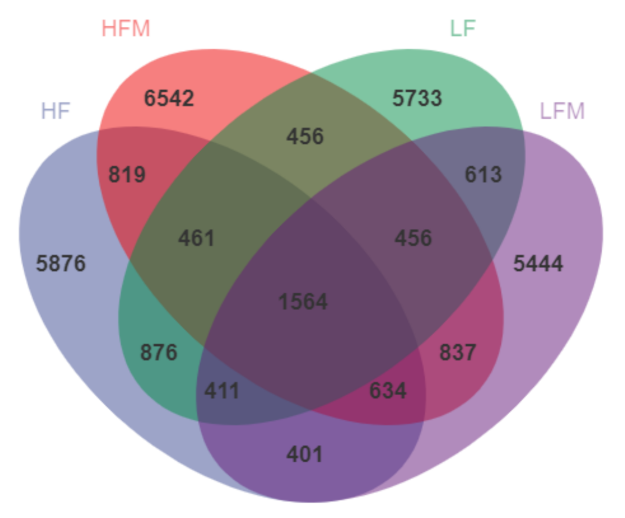
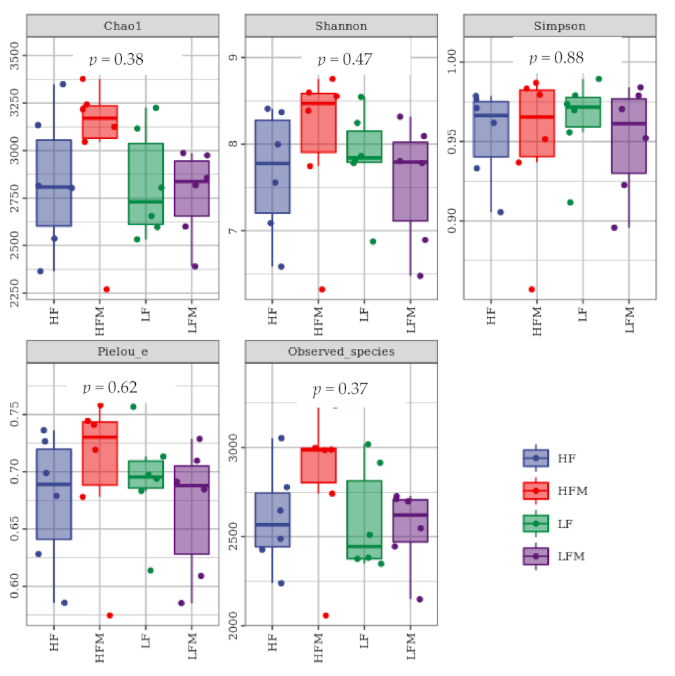
In general, high-fiber diets and fermented feed had a positive effect on the composition and diversity of pig fecal microorganisms and the relative abundance of beneficial bacteria. The HF group had the largest number of species, and the highest species richness and uniformity. Through microbial community analysis,
Firmicutes, accounting for 70–80% of the total, was still the most important phyla in the four groups at the phylum level, followed by
Bacteroidetes. The relative abundance of
Firmicutes in the fermented feed group was relatively high, which was related to the fact that
Lactobacillus amylovorus,
Enterococcus faecalis, and
Bacillus subtilis participating in fermentation all belong to
Firmicutes. At the same time,
Firmicutes were the main degradation bacteria of macromolecules such as cellulose and resistant starch. The produced SCFAs, especially butyric acid, mainly provide energy for the host colonic epithelial cells, and also have a probiotic effect, such as anti-inflammatory and antioxidant [
37]. Studies have shown that the body fat rate of pigs is related to the relative abundance of
Firmicutes and
Bacteroidetes.
Bacteroidetes are common microorganisms that degrade polysaccharides [
38]. Researchers found that removing
Bacteroidetes could promote the secretion of glucagon-like peptide 1 [
39], and the relative abundance of
Bacteroidetes in obese pigs was lower than that in lean pigs [
40]. In this experiment, feeding fermented feed significantly reduced the relative abundance of
Bacteroidetes, and the backfat thickness of fermentation groups was significantly higher than that of the unfermented feed group, which is in accordance with the above conclusion and is also similar to the experimental results of Cui et al. [
41]. The experiment of Cui et al. also proved that the effect of probiotics on lipid metabolism enzyme mRNAs is the opposite in subcutaneous fat and liver, which explains why fermented feed reduced serum TC and TG while increasing the backfat depth of finishing pigs in this experiment.
At the family level,
Lactobacillaceae and
Ruminococcaceaes were the two dominant groups, and the multi-bacteria fermented feed significantly improved the relative abundance of
Lactobacillaceae. Previous studies revealed that
Lactobacillaceae was involved in the biotransformation of secondary bile acids [
42,
43,
44]. Compared with primary bile acids, secondary bile acids are less soluble and easier to be excreted. Large excretion of secondary bile acids required the supplement of cholesterol from de novo synthesis. Therefore,
Lactobacillaceae had the potential of reducing cholesterol [
45], which was consistent with the results of serum biochemical indicators in this experiment.
Lactobacillaceae was able to secrete lactic acid and decrease the pH in the intestines, which is beneficial to the degradation of macromolecular substances. Multi-bacteria fermentation tended to increase the relative abundance of
Peptostreptococcaceae and reduce the relative abundance of
S24-7.
Peptostreptococcaceae, as an intestinal symbiotic bacteria, has been proven to be able to produce volatile fatty acids, and maintain the host intestinal homeostasis and energy metabolism [
46].
S24-7 is a type of microorganism related to intra-abdominal infections [
47].
At the genus level,
Lactobacillus and
Streptococcus were the two dominant groups in the fermented feed group and the non-fermented feed group respectively, similar to the family level. Fermentation significantly reduced the abundance of the pathogenic bacteria
Streptococcus. The abundance of
Clostridiaceae_Clostridium was closely related to dietary protein levels and is usually the most important genus in high-protein diets [
48]. High-fiber diets significantly reduced the relative abundance of
Clostridiaceae_Clostridium, which was presumably due to the fact that excessive fiber concentration leads to excessive protein loss in the digestive tract and a decrease in fermentable protein substrates [
49]. Although the difference in the abundance of
Bifidobacterium at the genus level was not significant,
Bifidobacterium was still a dominant species in the LF group.
Bifidobacterium had an active role in anti-inflammatory and anti-pathogenic microorganisms and prevented pathogenic infection by producing acetate [
50]. As the dominant species in the HF group,
Coprococcus was a kind of beneficial bacteria that can use carbohydrates to produce butyrate [
49] to achieve probiotic effects. Both fermentation and high-fiber diet had a significant effect on it, and the interaction was significant as well.
Oscillospira is a biomarker of the LN group and a kind of butyric acid-producing bacteria as well, but the substrate glucuronate of
Oscillospira is a common host-derived sugar [
51], which makes
Oscillospira related to body thinness [
52], and studies have shown that the relative abundance of
Oscillospira in the cecum of birds, fish, and mice increased after long-term fasting [
53].
In terms of function prediction, feeding fermented feed had a significant effect on metabolic pathways associated with synthesis, degradation, and transformation of substances in finishing pigs. For example, fermented feed promoted the biosynthesis of polyamine (YAMINSYN3-PWY), which affects RNA and DNA structure, ribosome function, and the activities of many enzymes, including kinase and phosphatase, and plays a role in controlling membrane potential and potassium homeostasis in glutamate receptor ion channels [
54]. Fermented feed also increased the pathways of ARGORNPROST-PWY (arginine, ornithine, and proline interconversion), GOLPDLCAT-PWY (glycerol degradation), PWY-7013 (propanediol degradation), and PWY-5022 (4-aminobutanoate degradation), which meant improvements of urea cycle efficiency and fatty acid utilization. In addition, the fermented feed was able to inhibit the biosynthesis of peptidoglycan (PWY-6470) and queuosine (PWY-6700). Peptidoglycan was the main component of the cell wall of many cocci [
55], and queuosine was the raw material of DNA and RNA, indicating that fermented feed has the potential to inhibit pathogens. High-fiber diets were able to increase the pathway of glycolysis, pyruvate dehydrogenase, TCA, and glyoxylate bypass (TCA-GLYOX-BYPASS and GLYCOLYSIS-TCA-GLYOX-BYPASS) to improve the catabolism of soluble sugar and promote chitin derivatives’ degradation (PWY-6906), which has an inhibition effect on pathogenic bacteria such as
Escherichia coli and
Salmonella typhimurium [
56].





{kind=link}
{kind=link}