
Experimental Studies on the Impact of the Projected Ocean Acidification on Fish Survival, Health, Growth, and Meat Quality; Black Sea Bream (Acanthopagrus schlegelii), Physiological and Histological Studies

 , , , and
, , , and
Abstract
:Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Experimental Design and Procedure, Seawater Parameters
pH Stabilization Curve
2.2. Sampling for Growth Parameters, Proximate Composition, Histological Studies
2.3. Method for the Proximate Composition of the Whole-Body and Dorsal Muscle
2.4. Histological Studies: H&E, SEM & TEM, Sample Preparation
2.4.1. Sample Preparation H&E (Hematoxylin and Eosin) Stain and Light Microscope Observation of Gills, Liver, Skin, Foregut, Midgut, and Hindgut
2.4.2. Foregut, Midgut, and Hindgut Preparation for SEM Observations
2.4.3. Liver Sample Treatment and TEM Observation
2.5. Statistical Analysis
3. Results
3.1. Seawater Physicochemical Parameters
3.2. Growth Parameters
3.3. Proximate Composition of the Fish Samples
3.4. SEM Observation of the Foregut Tissue
3.5. H&E Stain: Observation of Foregut, Midgut, and Hindgut
3.6. H&E Stain: Light Microscope Observation of the Liver
3.7. H&E Stain: Light Microscope Observation of Skin
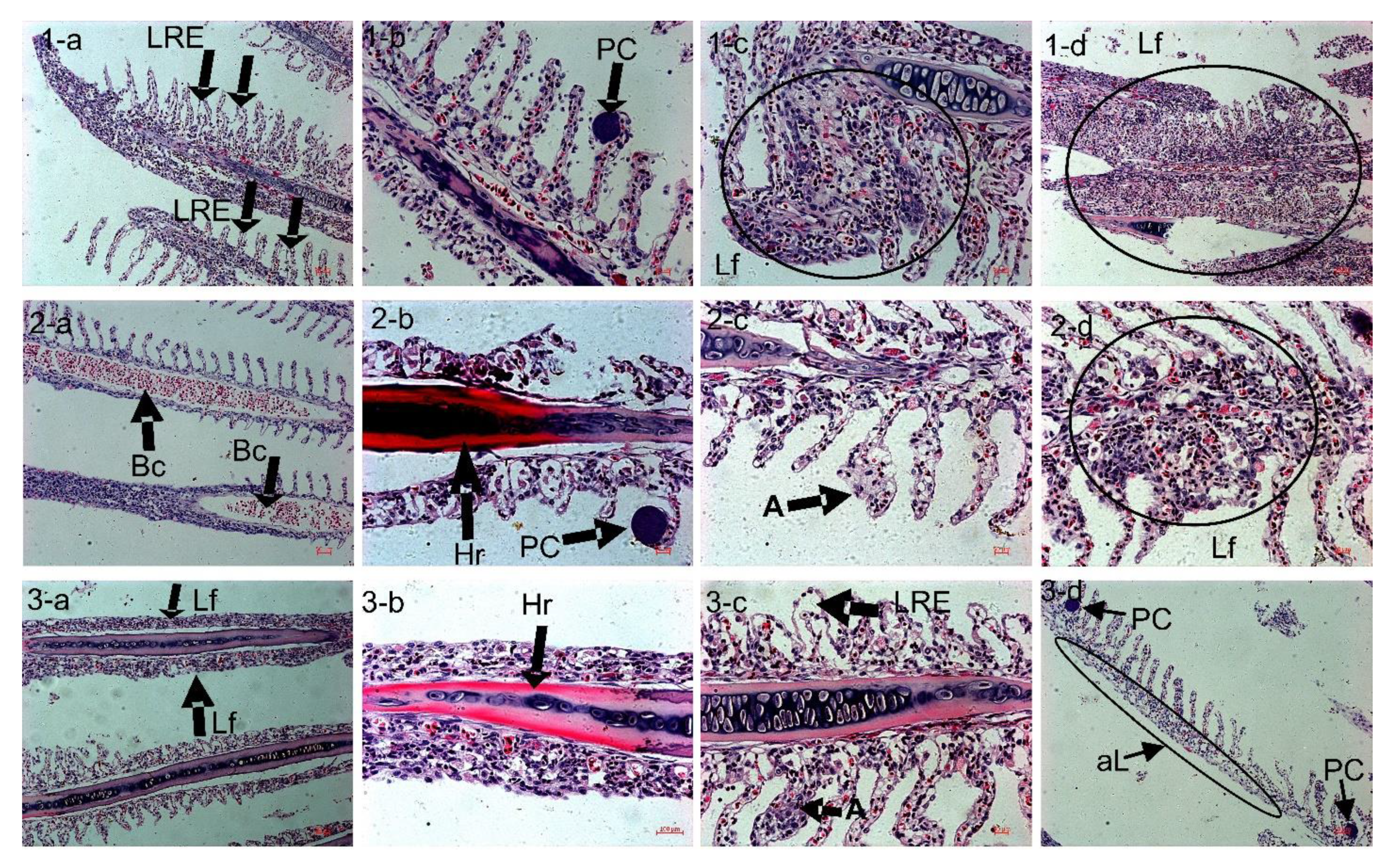
3.8. H&E Stain: Light Microscope Observation of the Gill
3.9. TEM Observation of Liver Cells (Inner Structure)
4. Discussion
4.1. Water Physicochemical Parameters
4.2. Proximate Composition Showing a Reduction in Meat Quality
4.3. Growth Parameters Revealing a Significant Low Growth and Confirming the Reduction in Meat Quality
4.4. The Survival Rate and the Histology of Liver and Gill Tissue Showing Degrading Effects on Fish Health
4.5. Histology Showing No Atrophy on Fish Skin Tissue
4.6. TEM Observation of Fish Liver Cell’s Inner Structure, Revealing Signs of Metabolic Acidosis
4.7. Histological Studies of Small Intestine Revealing Microvilli Atrophy
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shukla, P.; Skea, J.; Calvo Buendia, E.; Masson-Delmotte, V.; Pörtner, H.; Roberts, D.; Zhai, P.; Slade, R.; Connors, S.; Van Diemen, R. Climate Change and Land: An IPCC Special Report on Climate Change, Desertification, Land Degradation, Sustainable Land Management, Food Security, and Greenhouse Gas Fluxes in Terrestrial Ecosystems; IPCC: Geneva, Switzerland, 2019. [Google Scholar]
- Dell’acqua, O.; Ferrando, S.; Chiantore, M.; Asnaghi, V. The impact of ocean acidification on the gonads of three key Antarctic benthic macroinvertebrates. Aquat. Toxicol. 2019, 210, 19–29. [Google Scholar] [CrossRef]
- Han, Y.; Shi, W.; Tang, Y.; Zhao, X.; Du, X.; Sun, S.; Zhou, W.; Liu, G. Ocean acidification increases polyspermy of a broadcast spawning bivalve species by hampering membrane depolarization and cortical granule exocytosis. Aquat. Toxicol. 2021, 231, 105740. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Lu, Y. Metabolite profiling of Breviolum minutum in response to acidification. Aquat. Toxicol. 2019, 213, 105215. [Google Scholar] [CrossRef]
- Lee, Y.H.; Kang, H.-M.; Kim, M.-S.; Wang, M.; Kim, J.H.; Jeong, C.-B.; Lee, J.-S. Effects of ocean acidification on life parameters and antioxidant system in the marine copepod Tigriopus japonicus. Aquat. Toxicol. 2019, 212, 186–193. [Google Scholar] [CrossRef] [PubMed]
- Shrivastava, J.; Ndugwa, M.; Caneos, W.; De Boeck, G. Physiological trade-offs, acid-base balance and ion-osmoregulatory plasticity in European sea bass (Dicentrarchus labrax) juveniles under complex scenarios of salinity variation, ocean acidification and high ammonia challenge. Aquat. Toxicol. 2019, 212, 54–69. [Google Scholar] [CrossRef]
- Sousa, G.T.; Neto, M.C.L.; Choueri, R.B.; Castro, Í.B. Photoprotection and antioxidative metabolism in Ulva lactuca exposed to coastal oceanic acidification scenarios in the presence of Irgarol. Aquat. Toxicol. 2021, 230, 105717. [Google Scholar] [CrossRef]
- Wang, M.; Jeong, C.-B.; Lee, Y.H.; Lee, J.-S. Effects of ocean acidification on copepods. Aquat. Toxicol. 2018, 196, 17–24. [Google Scholar] [CrossRef]
- Xing, Q.; Liao, H.; Peng, C.; Zheng, G.; Yang, Z.; Wang, J.; Lu, W.; Huang, X.; Bao, Z. Identification, characterization and expression analyses of cholinesterases genes in Yesso scallop (Patinopecten yessoensis) reveal molecular function allocation in responses to ocean acidification. Aquat. Toxicol. 2021, 231, 105736. [Google Scholar] [CrossRef]
- Zhao, L.; Liu, L.; Liu, B.; Liang, J.; Lu, Y.; Yang, F. Antioxidant responses to seawater acidification in an invasive fouling mussel are alleviated by transgenerational acclimation. Aquat. Toxicol. 2019, 217, 105331. [Google Scholar] [CrossRef] [PubMed]
- Ciais, P.; Sabine, C.; Bala, G.; Bopp, L.; Brovkin, V.; Canadell, J.; Chhabra, A.; Defries, R.; Galloway, J.; Heimann, M. Carbon and other biogeochemical cycles. Climate change 2013: The physical science basis. In Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2014; pp. 465–570. [Google Scholar]
- Cao, R.; Wang, Q.; Yang, D.; Liu, Y.; Ran, W.; Qu, Y.; Wu, H.; Cong, M.; Li, F.; Ji, C.; et al. CO2-induced ocean acidification impairs the immune function of the Pacific oyster against Vibrio splendidus challenge: An integrated study from a cellular and proteomic perspective. Sci. Total Environ. 2018, 625, 1574–1583. [Google Scholar] [CrossRef]
- Cattano, C.; Agostini, S.; Harvey, B.P.; Wada, S.; Quattrocchi, F.; Turco, G.; Inaba, K.; Hall-Spencer, J.M.; Milazzo, M. Changes in fish communities due to benthic habitat shifts under ocean acidification conditions. Sci. Total Environ. 2020, 725, 138501. [Google Scholar] [CrossRef]
- Gallo, A.; Boni, R.; Buia, M.C.; Monfrecola, V.; Esposito, M.C.; Tosti, E. Ocean acidification impact on ascidian Ciona robusta spermatozoa: New evidence for stress resilience. Sci. Total Environ. 2019, 697, 134100. [Google Scholar] [CrossRef] [PubMed]
- Kang, E.J.; Han, A.R.; Kim, J.-H.; Kim, I.N.; Lee, S.; Min, J.O.; Nam, B.R.; Choi, Y.J.; Edwards, M.S.; Diaz-Pulido, G. Evaluating bloom potential of the green-tide forming alga Ulva ohnoi under ocean acidification and warming. Sci. Total Environ. 2021, 769, 144443. [Google Scholar] [CrossRef] [PubMed]
- Porzio, L.; Arena, C.; Lorenti, M.; De Maio, A.; Buia, M.C. Long-term response of Dictyota dichotoma var. intricata (C. Agardh) Greville (Phaeophyceae) to ocean acidification: Insights from high pCO2 vents. Sci. Total Environ. 2020, 731, 138896. [Google Scholar] [CrossRef]
- Sampaio, E.; Lopes, A.R.; Francisco, S.; Paula, J.R.; Pimentel, M.; Maulvault, A.L.; Repolho, T.; Grilo, T.F.; Pousão-Ferreira, P.; Marques, A.; et al. Ocean acidification dampens physiological stress response to warming and contamination in a commercially-important fish (Argyrosomus regius). Sci. Total Environ. 2018, 618, 388–398. [Google Scholar] [CrossRef]
- Zhong, J.; Guo, Y.; Liang, Z.; Huang, Q.; Lu, H.; Pan, J.; Li, P.; Jin, P.; Xia, J. Adaptation of a marine diatom to ocean acidification and warming reveals constraints and trade-offs. Sci. Total Environ. 2021, 771, 145167. [Google Scholar] [CrossRef] [PubMed]
- Feely, R.A.; Doney, S.C.; Cooley, S.R. Ocean acidification: Present conditions and future changes in a high-CO2 world. Oceanography 2009, 22, 36–47. [Google Scholar] [CrossRef] [Green Version]
- Araujo, J.E.; Madeira, D.; Vitorino, R.; Repolho, T.; Rosa, R.; Diniz, M. Negative synergistic impacts of ocean warming and acidification on the survival and proteome of the commercial sea bream, sparus aurata. J. Sea Res. 2018, 139, 50–61. [Google Scholar] [CrossRef]
- Fabry, V.J.; Mcclintock, J.B.; Mathis, J.T.; Grebmeier, J.M. Ocean acidification at high latitudes: The bellweather. Oceanography 2009, 22, 160–171. [Google Scholar] [CrossRef] [Green Version]
- Gattuso, J.P.; Lavigne, H. Technical Note: Approaches and software tools to investigate the impact of ocean acidification. Biogeosciences 2009, 6, 2121–2133. [Google Scholar] [CrossRef] [Green Version]
- Harney, E.; Artigaud, S.; Souchu, P.L.; Miner, P.; Corporeau, C.; Essid, H.; Pichereau, V.; Nunes, F.L.D. Non-additive effects of ocean acidification in combination with warming on the larval proteome of the Pacific oyster, Crassostrea gigas. J. Proteom. 2016, 135, 151–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leis, J.M. Paradigm Lost: Ocean Acidification Will Overturn the Concept of Larval-Fish Biophysical Dispersal. Front. Mar. Sci. 2018, 5, 47. [Google Scholar] [CrossRef] [Green Version]
- Orr, J.; Fabry, V.; Aumont, O.; Bopp, L.; Doney, S.; Feely, R.; Gnanadesikan, A.; Gruber, N.; Ishida, A.; Joos, F.; et al. Anthropogenic ocean acidification over the twenty-first century and its impact on calcifying organisms. Nature 2005, 437, 681–686. [Google Scholar] [CrossRef] [PubMed]
- Watson, S.-A.; Allan, B.J.M.; Mcqueen, D.E.; Nicol, S.; Parsons, D.M.; Pether, S.M.J.; Pope, S. Ocean warming has a greater effect than acidification on the early life history development and swimming performance of a large circumglobal pelagic fish. Glob. Chang. Biol. 2018, 24, 4368–4385. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Guo, C.; Han, Y.; Che, Z.; Wang, Y.; Wand, X.; Chai, X.; Wu, H.; Liu, G. Ocean acidification decreases mussel byssal attachment strength and induces molecular byssal responses. Mar. Ecol. Progess Ser. 2017, 568, 67–77. [Google Scholar] [CrossRef]
- Laubenstein, T.D.; Rummer, J.L.; Nicol, S.; Parsons, D.M.; Pether, S.M.J.; Pope, S.; Smith, N.; Munday, P.L. Correlated Effects of Ocean Acidification and Warming on Behavioral and Metabolic Traits of a Large Pelagic Fish. Diversity 2018, 10, 35. [Google Scholar] [CrossRef] [Green Version]
- Peng., C.; Zhao, X.; Liu, S.; Shi, W.; Han, Y.; Guo, C.; Peng, X.; Chai, X.; Liu, G. Ocean acidification alters the burrowing behaviour, Ca2+/Mg2+-ATPase activity, metabolism, and gene expression of a bivalve species, Sinonovacula constricta. Mar. Ecol. Prog. Ser. 2017, 575, 107–117. [Google Scholar] [CrossRef]
- Mora, C.; Wei, C.L.; Rollo, A.; Amaro, T.; Basco, A.R.; Billett, D.; Bopp, L.; Chen, Q.; Collier, M.; Danovaro, R.; et al. Biotic and human vulnerability to projected changes in ocean biogeochemistry over the 21st century. PLoS Biol. 2013, 11, e1001682. [Google Scholar] [CrossRef] [Green Version]
- Gazeau, F.; Parker, L.; Comeau, S.; Gattuso, J.-P.; O’connor, W.; Martin, S.; Pörtner, H.-O.; Ross, P. Impacts of ocean acidification on marine shelled molluscs. Mar. Biol. 2013, 160, 2207–2245. [Google Scholar] [CrossRef] [Green Version]
- Caldeira, K.; Wickett, M. Oceanography: Anthropogenic carbon and ocean pH. Nature 2003, 425, 365. [Google Scholar] [CrossRef]
- Doney, S.; Balch, W.; Fabry, V.; Feely, R. Ocean Acidification: A Critical Emerging Problem for the Ocean Sciences. Oceanography 2009, 22, 16–25. [Google Scholar] [CrossRef] [Green Version]
- Eumofa, E.U. The EU fish market. Eur. Mark. Obs. Fish. Aquac. Prod. 2017. [Google Scholar]
- Marino, G.; Crosetti, D.; Petochi, T.; FAO 2006-2015. National Aquaculture Sector Overview. In National Aquaculture Sector Overview Fact Sheets; Marino, G., Crosetti, D., Petochi, T., Eds.; Updated 19 October 2015; FAO Fisheries and Aquaculture Department: Rome, Italy; Available online: http://www.fao.org/fishery/countrysector/naso_italy/en (accessed on 12 March 2020).
- Zhao, X.; Han, Y.; Chen, B.; Xia, B.; Qu, K.; Liu, G. CO2-driven ocean acidification weakens mussel shell defense capacity and induces global molecular compensatory responses. Chemosphere 2020, 243, 125415. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, E.B.; Nagasawa, K.; Umino, T. Stock enhancement program for black sea bream (Acanthopagrus schlegelii) in Hiroshima Bay: Monitoring the genetic effects. Aquaculture 2008, 276, 36–43. [Google Scholar] [CrossRef] [Green Version]
- Shao, Q.; Ma, J.; Xu, Z.; Hu, W.; Xu, J.; Xie, S. Dietary phosphorus requirement of juvenile black seabream, Sparus macrocephalus. Aquaculture 2008, 277, 92–100. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, W.; Gladstone, S.; Ng, W.-K.; Zhang, J.; Shao, Q. Effects of isoenergetic diets with varying protein and lipid levels on the growth, feed utilization, metabolic enzymes activities, antioxidative status and serum biochemical parameters of black sea bream (Acanthopagrus schlegelii). Aquaculture 2019, 513, 734397. [Google Scholar] [CrossRef]
- Irm, M.; Taj, S.; Jin, M.; Timothée Andriamialinirina, H.J.; Cheng, X.; Zhou, Q. Influence of dietary replacement of fish meal with fish soluble meal on growth and TOR signaling pathway in juvenile black sea bream (Acanthopagrus schlegelii). Fish Shellfish Immunol. 2020, 101, 269–276. [Google Scholar] [CrossRef]
- Sako, H. Recent trends in fish diseases in Japan. In Towards Sustainable Aquaculture in Southeast Asia and Japan: Proceedings of the Seminar-Workshop on Aquaculture Development in Southeast Asia, Iloilo City, Philippines, 26–28 July 1994; Aquaculture Department, Southeast Asian Fisheries Development Center: Bangkok, Thailand, 1995. [Google Scholar]
- Hong, W.; Zhang, Q. Review of captive bred species and fry production of marine fish in China. Aquaculture 2003, 227, 305–318. [Google Scholar] [CrossRef]
- Tan, P.; Wu, X.; Zhu, W.; Lou, B.; Chen, R.; Wang, L. Effect of tributyrin supplementation in high-soya bean meal diet on growth performance, body composition, intestine morphology and microbiota of juvenile yellow drum (Nibea albiflora). Aquac. Res. 2020, 51, 2004–2019. [Google Scholar] [CrossRef]
- Wang, L.; Xiao, J.-X.; Hua, Y.; Xiang, X.-W.; Zhou, Y.-F.; Ye, L.; Shao, Q.-J. Effects of dietary selenium polysaccharide on growth performance, oxidative stress and tissue selenium accumulation of juvenile black sea bream, Acanthopagrus schlegelii. Aquaculture 2019, 503, 389–395. [Google Scholar] [CrossRef]
- Habiba, M.M.; Hussein, E.E.; Ashry, A.M.; El-Zayat, A.M.; Hassan, A.M.; El-Shehawi, A.M.; Sewilam, H.; Van Doan, H.; Dawood, M.A.O. Dietary Cinnamon Successfully Enhanced the Growth Performance, Growth Hormone, Antibacterial Capacity, and Immunity of European Sea Bass (Dicentrarchus labrax). Animals 2021, 11, 2128. [Google Scholar] [CrossRef]
- Sagada, G.; Gray, N.; Wang, L.; Xu, B.; Zheng, L.; Zhong, Z.; Ullah, S.; Tegomo, A.F.; Shao, Q. Effect of dietary inactivated Lactobacillus plantarum on growth performance, antioxidative capacity, and intestinal integrity of black sea bream (Acanthopagrus schlegelii) fingerlings. Aquaculture 2021, 535, 736370. [Google Scholar] [CrossRef]
- Alvarez, A.; Garcia Garcia, B.; Garrido, M.D.; Garrido, M.D.; Hernández, M. The influence of starvation time prior to slaughter on the quality of commercial-sized gilthead seabream (Sparus aurata) during ice storage. Aquaculture 2008, 284, 106–114. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis of the Association of Official Analytical Chemists; AOAC: Rockville, MD, USA, 2000. [Google Scholar]
- Wang, C.; Shen, Z.; Cao, S.; Zhang, Q.; Peng, Y.; Hong, Q.; Feng, J.; Hu, C. Effects of tributyrin on growth performance, intestinal microflora and barrier function of weaned pigs. Anim. Feed. Sci. Technol. 2019, 258, 114311. [Google Scholar] [CrossRef]
- Zhang, K.; Yu, D.; Li, Z.; Xie, J.; Wang, G.; Gong, W.; Yu, E.; Tian, J. Influence of eco-substrate addition on organic carbon, nitrogen and phosphorus budgets of intensive aquaculture ponds of the Pearl River, China. Aquaculture 2020, 520, 734868. [Google Scholar] [CrossRef]
- Munro, B.H. Manual of Histologic Staining Methods of the Armed Forces Institute of Pathology. Pathology 1971, 3, 249. [Google Scholar] [CrossRef]
- Titford, M. Progress in the Development of Microscopical Techniques for Diagnostic Pathology. J. Histotechnol. 2009, 32, 9–19. [Google Scholar] [CrossRef]
- Mzengereza, K.; Ishikawa, M.; Koshio, S.; Yokoyama, S.; Yukun, Z.; Shadrack, R.S.; Seo, S.; Duy Khoa, T.N.; Moss, A.; Dossou, S.; et al. Effect of Substituting Fish Oil with Camelina Oil on Growth Performance, Fatty Acid Profile, Digestibility, Liver Histology, and Antioxidative Status of Red Seabream (Pagrus major). Animals 2021, 11, 1990. [Google Scholar] [CrossRef]
- Adamek-Urbańska, D.; Błażewicz, E.; Sobień, M.; Kasprzak, R.; Kamaszewski, M. Histological Study of Suprabranchial Chamber Membranes in Anabantoidei and Clariidae Fishes. Animals 2021, 11, 1158. [Google Scholar] [CrossRef]
- Al-Khalaifah, H.S.; Amer, S.A.; Al-Sadek, D.M.M.; Khalil A, A.; Zaki E, M.; El-Araby, D.A. Optimizing the Growth, Health, Reproductive Performance, and Gonadal Histology of Broodstock Fantail Goldfish (Carassius auratus, L.) by Dietary Cacao Bean Meal. Animals 2020, 10, 1808. [Google Scholar] [CrossRef]
- Lynch, J.M.; Barbano, D.M. Kjeldahl nitrogen analysis as a reference method for protein determination in dairy products. J. AOAC Int. 1999, 82, 1389–1398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, V.A.S.; Gelcich, S.; Vásquez Lavín, F.; Ponce Oliva R, D.; Hernández J, I.; Lagos N, A.; Birchenough S N, R.; Vargas, C.A. Linking social preferences and ocean acidification impacts in mussel aquaculture. Sci. Rep. 2019, 9, 4719. [Google Scholar] [CrossRef] [PubMed]
- Rossoll, D.; Bermúdez, R.; Hauss, H.; Schulz, K.G.; Riebesell, U.; Sommer, U.; Winder, M. Ocean Acidification-Induced Food Quality Deterioration Constrains Trophic Transfer. PLoS ONE 2012, 7, e34737. [Google Scholar]
- Jin, P.; Hutchins, D.A.; Gao, K. The Impacts of Ocean Acidification on Marine Food Quality and Its Potential Food Chain Consequences. Front. Mar. Sci. 2020, 7, 543979. [Google Scholar] [CrossRef]
- Zhang, H.; Shin, P.K.S.; Cheung, S.G. Physiological responses and scope for growth upon medium-term exposure to the combined effects of ocean acidification and temperature in a subtidal scavenger Nassarius conoidalis. Mar. Environ. Res. 2015, 106, 51–60. [Google Scholar] [CrossRef]
- Ruiz-Jarabo, I.; Gregório, S.F.; Alves, A.; Mancera J, M.; Fuentes, J. Ocean acidification compromises energy management in Sparus aurata (Pisces: Teleostei). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2021, 256, 110911. [Google Scholar] [CrossRef]
- Bernet, D.; Schmidt, H.; Meier, W.; Burkhardt-Holm, P.; Wahli, T. Histopathology in fish: Proposal for a protocol to assess aquatic pollution. J. Fish Dis. 1999, 22, 25–34. [Google Scholar] [CrossRef] [Green Version]
- Faheem, M.; Jahan, N.; Lone, K. Histopathological effects of bisphenol-a on liver, kidneys and gills of Indian major carp, Catla catla (Hamilton, 1822). J. Anim. Plant Sci. 2016, 26, 514–522. [Google Scholar]
- Santos, D.M.S.; Melo, M.R.S.; Mendes, D.C.S.; Rocha I K B, S.; Silva J P, L.; Cantanhêde S, M.; Meletti, P.C. Histological Changes in Gills of Two Fish Species as Indicators of Water Quality in Jansen Lagoon (São Luís, Maranhão State, Brazil). Int. J. Environ. Res. Public Health 2014, 11, 12927–12937. [Google Scholar] [CrossRef] [Green Version]
- Fromm, P.O. A review of some physiological and toxicological responses of freshwater fish to acid stress. Environ. Biol. Fishes 1980, 5, 79–93. [Google Scholar] [CrossRef]
- Wurts, W. Interactions of pH, Carbon Dioxide, Alkalinity and Hardness in Fish Ponds. South. Reg. Aquac. Cent. 1992. [Google Scholar]
- Iancu, T.; Manov, I. Electron Microscopy of Liver Biopsies. Nutrition 2011, 16, 246–267. [Google Scholar]
- Liao, H.; Yang, Z.; Dou, Z.; Sun, F.; Kou, S.; Zhang, Z.; Huang, X.; Bao, Z. Impact of Ocean Acidification on the Energy Metabolism and Antioxidant Responses of the Yesso Scallop (Patinopecten yessoensis). Front. Physiol. 2019, 9, 1967. [Google Scholar] [CrossRef] [Green Version]
- Goss, G.G.; Perry, S.F.; Fryer, J.N.; Laurent, P. Gill Morphology and Acid-Base Regulation in Freshwater Fishes. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 1998, 119, 107–115. [Google Scholar] [CrossRef]
- Allmon, E.B.; Esbaugh, A.J. Carbon dioxide induced plasticity of branchial acid-base pathways in an estuarine teleost. Sci. Rep. 2017, 7, 45680. [Google Scholar] [CrossRef] [PubMed]
- Guzoglu, N.; Aliefendioglu, D.; Gulerman, F.; Gucer, S.; Kaymaz, F. Congenital Microvillus Inclusion Disease in the Differential Diagnosis of Intractable Metabolic Acidosis. Pediatr. Neonatol. 2017, 58, 285–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagen, S.J.; Trier, J.S.; Dambrauskas, R. Exposure of the rat small intestine to raw kidney beans results in reorganization of absorptive cell microvilli. Gastroenterology 1994, 106, 73–84. [Google Scholar] [CrossRef]
- Sidhaye, J.; Pinto, C.; Dharap, S.; Jacob, T.; Bhargava, S.; Sonawane, M. The zebrafish goosepimples/myosin Vb mutant exhibits cellular attributes of human microvillus inclusion disease. Mech. Dev. 2016, 142, 62–74. [Google Scholar] [CrossRef]
- Bunn, S.K.; Beath, S.V.; Mckeirnan, P.J.; Kelly D, A.; Buckles, J.A.C.; Mirza, D.; Mayer, A.D.; De Goyet, J.D. Treatment of microvillus inclusion disease by intestinal transplantation. J. Pediatric Gastroenterol. Nutr. 2000, 31, 176–180. [Google Scholar] [CrossRef]
- Caamano, B.F.; Blanco, M.J.Q.; Tome, L.F.; Lizaldez E, B.; Oses J, S.; Arias M, M.; Bozano, G.P. Intestinal failure and transplantation in microvillous inclusion disease. An. Pediatría 2015, 83, 160–165. [Google Scholar]
- Heissler, S.M.; Chinthalapudi, K.; Sellers, J.R. Kinetic signatures of myosin-5B, the motor involved in microvillus inclusion disease. J. Biol. Chem. 2017, 292, 18372–18385. [Google Scholar] [CrossRef] [Green Version]
- Phulware, R.H.; Gahlot, G.P.S.; Malik, R.; Gupta S, D.; Das, P. Microvillous Inclusion Disease as a Cause of Protracted Diarrhea. Indian J. Pediatrics 2019, 86, 854–856. [Google Scholar] [CrossRef] [PubMed]
- Randak, C.; Langnas, A.N.; Kaufman, S.S.; Phillips A, D.; Wisecarver J, L.; Hadorn H, B.; Vanderhoof, J.A. Pretransplant management and small bowel-liver transplantation in an infant with microvillus inclusion disease. J. Pediatric Gastroenterol. Nutr. 1998, 27, 333–337. [Google Scholar] [CrossRef]
- Ruemmele, F.M.; Schmitz, J.; Goulet, O. Microvillous inclusion disease (microvillous atrophy). Orphanet J. Rare Dis. 2006, 1, 22. [Google Scholar] [CrossRef] [Green Version]
- Saadah, O.I.; Bokhary, R.Y.; Jalalah, S.M.; Bin-Taleb, Y.Y. Microvillus inclusion disease: A clinicopathological study from western region of Saudi Arabia. J. Microsc. Ultrastruct. 2013, 1, 84–88. [Google Scholar] [CrossRef] [Green Version]
- Van Hoeve, K.; Hoffman, I.; Fusaro, F.; Pirenne, J.; Vander Auwera, A.; Dieltjens A, M.; De Hertogh, G.; Monbaliu, D.; Miserez, M. Microvillus inclusion disease: A subtotal enterectomy as a bridge to transplantation. Acta Chir. Belg. 2016, 116, 333–339. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | Predict_A | Predict_B | |
|---|---|---|---|
| Targeted pH | 8.10 | 7.80 | 7.40 |
| Seawater parameters | |||
| Salinity (g∙L−1) | 27.13 ± 1.06 | 27.10 ± 1.10 | 27.17 ± 1.09 |
| Temperature (°C) | 27.53 ± 0.85 | 27.51 ± 0.74 | 27.47 ± 0.87 |
| Measured pH | 8.10 ± 0.01 a | 7.80 ± 0.02 b | 7.40 ± 0.02 c |
| TA | 2051.43 ± 16.11 b | 2097.40 ± 20.15 a | 2065.39 ± 17.22 ab |
| pCO2 (µatm) | 321.37 ± 11.48 c | 749.12 ± 27.03 b | 1993.71 ± 102.12 a |
| HCO3− (µmol∙kg−1) | 1573.43 ± 20.10 c | 1823.60 ± 14.65 b | 1947.91 ± 18.77 a |
| CO32− (µmol∙kg−1) | 196.82 ± 1.45 a | 113.41 ± 4.53 b | 48.57 ± 1.93 c |
| DIC (µmol∙kg−1) | 1779.15 ± 19.01 c | 1957.74 ± 16.45 b | 2054.48 ± 20.80 a |
| Ωara | 3.29 ± 0.024 a | 1.90 ± 0.08 b | 0.81 ± 0.03 c |
| Ωcal | 5.07 ± 0.04 a | 2.92 ± 0.12 b | 1.25 ± 0.50 c |
| Control | Predict_A | Predict_B | |
|---|---|---|---|
| Targeted pH | 8.10 | 7.80 | 7.40 |
| Initial weight (IW) (g/fish) | 2.73 ± 0.01 | 2.72 ± 0.02 | 2.73 ± 0.01 |
| Final weight (FW) (g/fish) | 20.86 ± 0.25 a | 17.54 ± 0.49 b | 15.80 ± 0.06 c |
| SR 1 (%) | 100 ± 0.00 | 100 ± 0.00 | 98.33 ± 2.89 |
| WG 2 (%) | 664.01 ± 6.79 a | 545.80 ± 21.57 b | 479.43 ± 3.73 c |
| SGR 3 (%/day) | 4.07 ± 0.02 a | 3.73 ± 0.07 b | 3.51 ± 0.01 c |
| FI 4 (%/day) | 29.38 ± 0.32 b | 30.81 ± 0.66 a | 30.39 ± 0.21 ab |
| FCR 5 | 1.04 ± 0.01 b | 1.14 ± 0.04 a | 1.17 ± 0.01 a |
| FE 6 (%) | 95.84 ± 0.88 a | 87.45 ± 3.21 b | 85.17 ± 0.99 b |
| HIS 7 (%) | 2.07 ± 0.24 | 1.91 ± 0.24 | 1.90 ± 0.23 |
| CF 8 (g/cm3) | 2.47 ± 0.09 | 2.93 ± 0.23 | 2.76 ± 0.22 |
| PER 9 | 2.44 ± 0.02 a | 2.23 ± 0.08 b | 2.17 ± 0.03 b |
| Control | Predict_A | Predict_B | |
|---|---|---|---|
| Proximate composition (%) | 8.10 | 7.80 | 7.40 |
| Whole-body | |||
| Moisture | 71.08 ± 0.18 | 70.48 ± 0.76 | 70.96 ± 0.69 |
| Crude Protein | 52.86 ± 1.08 a | 50.43 ± 1.20 ab | 48.88 ± 1.16 b |
| Crude Lipid | 20.71 ± 0.71 | 21.38 ± 0.53 | 20.35 ± 0.60 |
| Ash | 5.02 ± 0.05 | 5.08 ± 0.18 | 5.08 ± 0.17 |
| Dorsal Muscle | |||
| Moisture | 76.39 ± 0.42 | 76.58 ± 0.35 | 76.57 ± 0.10 |
| Crude Protein | 70.98 ± 1.12 a | 69.61 ± 1.52 ab | 67.87 ± 0.87 b |
| Crude Lipid | 6.06 ± 1.08 | 5.56 ± 0.57 | 5.63 ± 0.20 |
| Ash | 4.94 ± 0.08 | 5.24 ± 0.53 | 5.03 ± 0.23 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tegomo, F.A.; Zhong, Z.; Njomoue, A.P.; Okon, S.U.; Ullah, S.; Gray, N.A.; Chen, K.; Sun, Y.; Xiao, J.; Wang, L.; et al. Experimental Studies on the Impact of the Projected Ocean Acidification on Fish Survival, Health, Growth, and Meat Quality; Black Sea Bream (Acanthopagrus schlegelii), Physiological and Histological Studies. Animals 2021, 11, 3119. https://doi.org/10.3390/ani11113119
Tegomo FA, Zhong Z, Njomoue AP, Okon SU, Ullah S, Gray NA, Chen K, Sun Y, Xiao J, Wang L, et al. Experimental Studies on the Impact of the Projected Ocean Acidification on Fish Survival, Health, Growth, and Meat Quality; Black Sea Bream (Acanthopagrus schlegelii), Physiological and Histological Studies. Animals. 2021; 11(11):3119. https://doi.org/10.3390/ani11113119
Chicago/Turabian StyleTegomo, Fabrice Arnaud, Zhiwen Zhong, Achille Pandong Njomoue, Samuel Ukpong Okon, Sami Ullah, Neveen Anandi Gray, Kai Chen, Yuxiao Sun, Jinxing Xiao, Lei Wang, and et al. 2021. "Experimental Studies on the Impact of the Projected Ocean Acidification on Fish Survival, Health, Growth, and Meat Quality; Black Sea Bream (Acanthopagrus schlegelii), Physiological and Histological Studies" Animals 11, no. 11: 3119. https://doi.org/10.3390/ani11113119