Waste Valorization via Hermetia Illucens to Produce Protein-Rich Biomass for Feed: Insight into the Critical Nutrient Taurine

 ,
,  ,
, 

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. H. Illucens Culture and Experimental Design
2.2. Nutritional Composition and Protein Yield
2.3. Determination of Amino Acid Profile
2.4. Minerals and Heavy Metals Analyses
2.5. Identification of the Genes Associated with Taurine Biosynthesis
2.6. Sequence Characterization and Phylogenetic Analysis
2.7. mRNA Levels of the Taurine Biosynthesis Genes
2.8. Statistical Analysis
3. Results
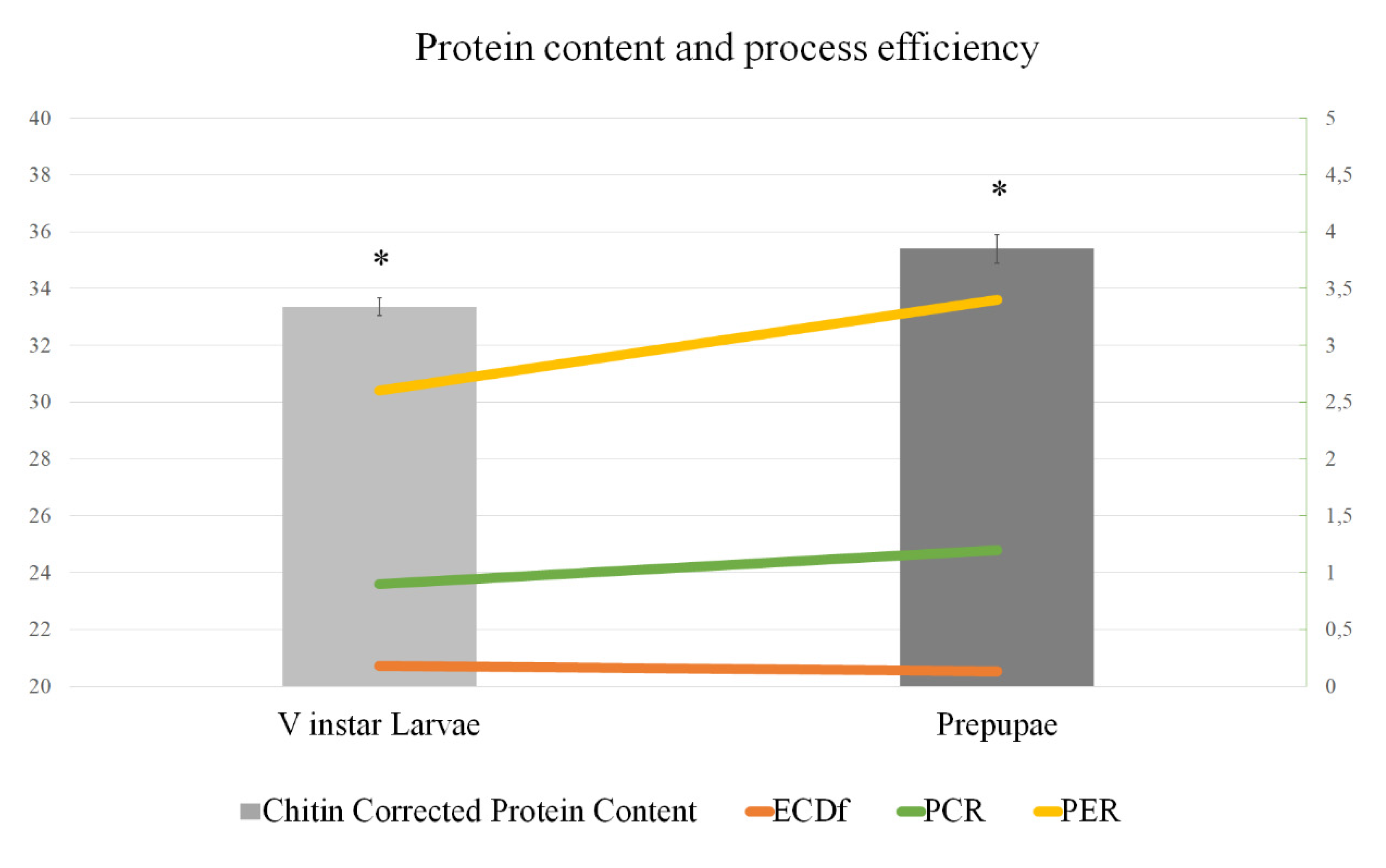
3.1. Nutrient Content of BSF V Instar Larvae and Prepupae and Efficiency of Protein Use
3.2. Amino Acid Profile of BSF V Instar Larvae and Prepupae
3.3. Minerals and Heavy Metals of BSF V Instar Larvae and Prepupae
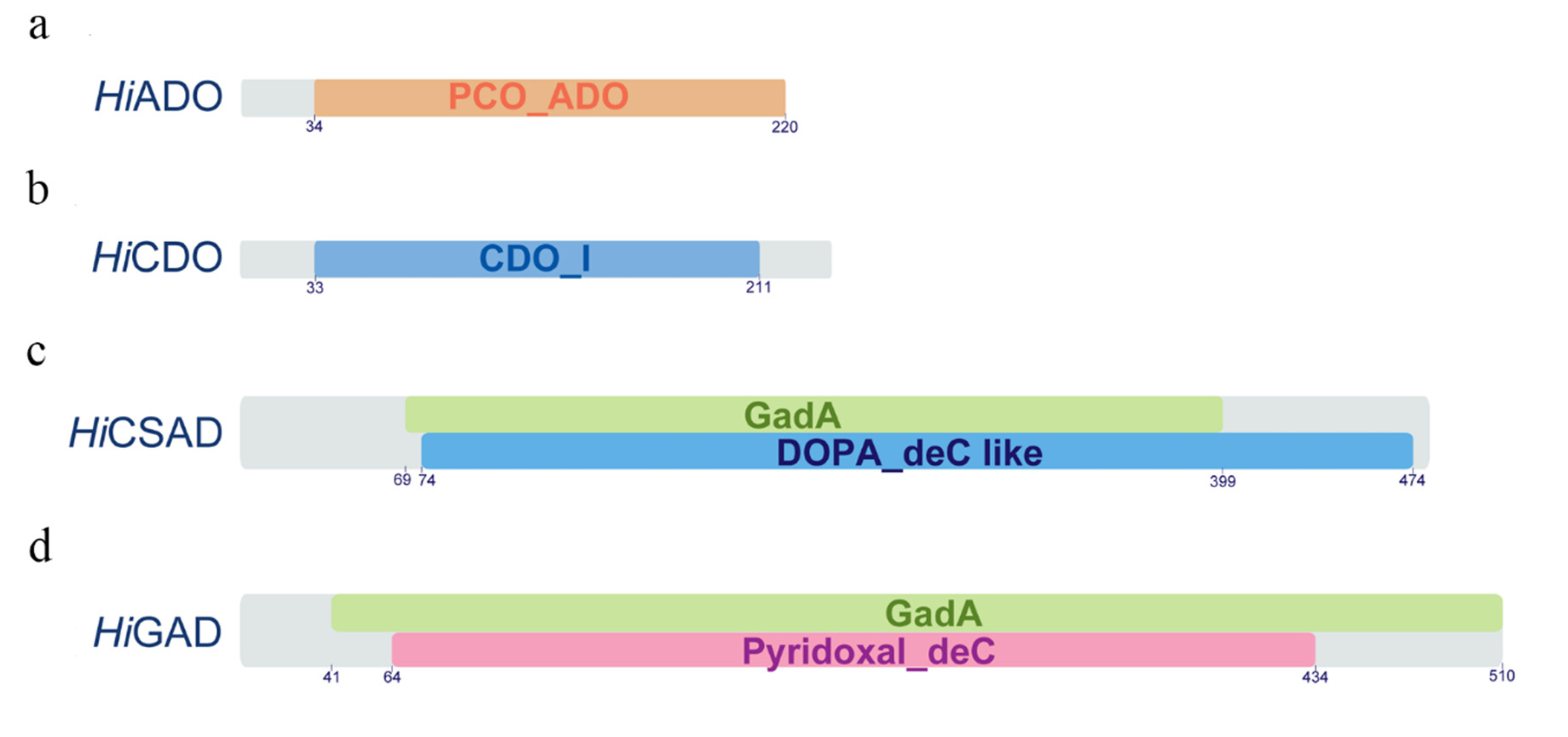
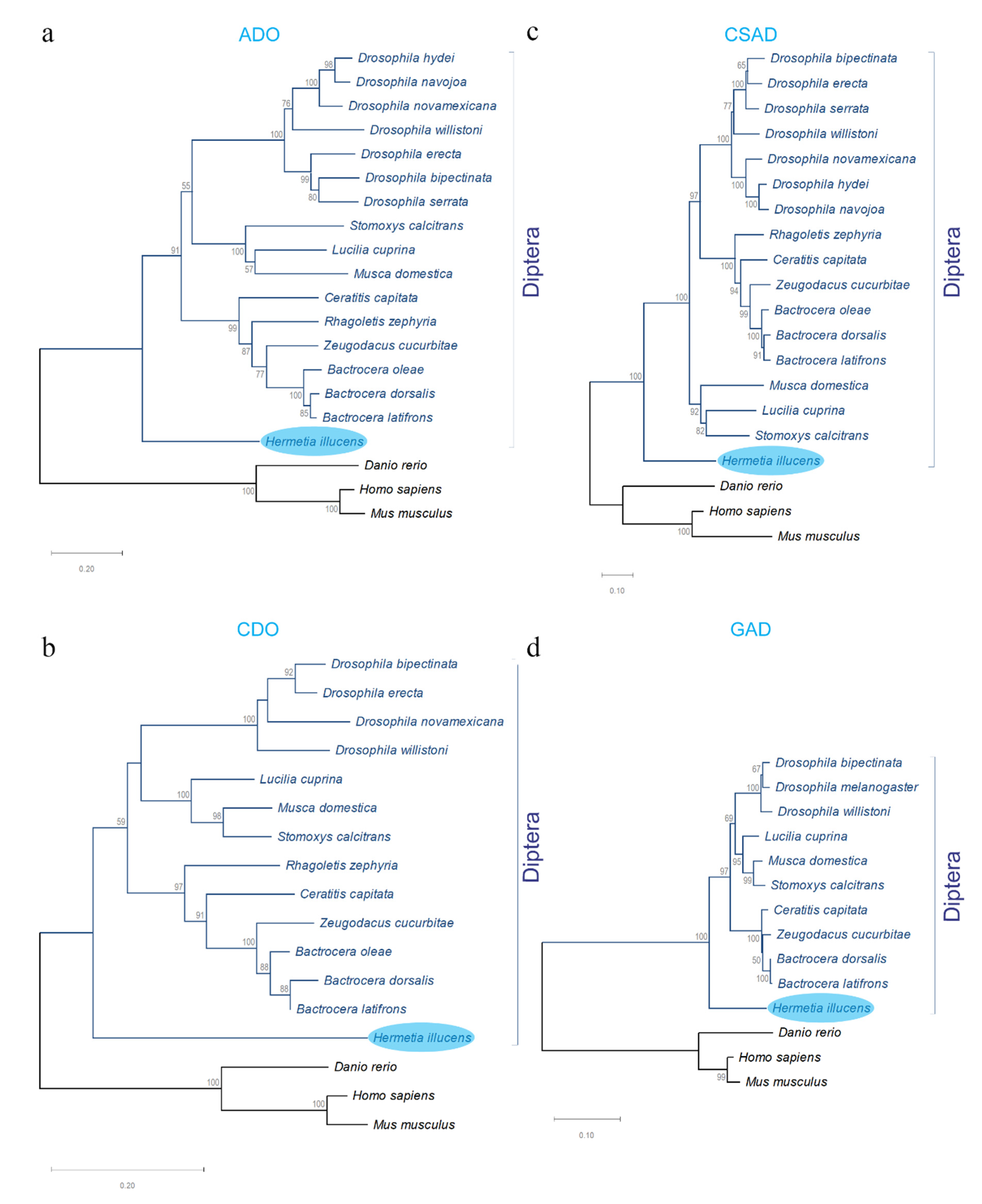
3.4. Molecular Characterization of the Genes Associated with Taurine Biosynthesis
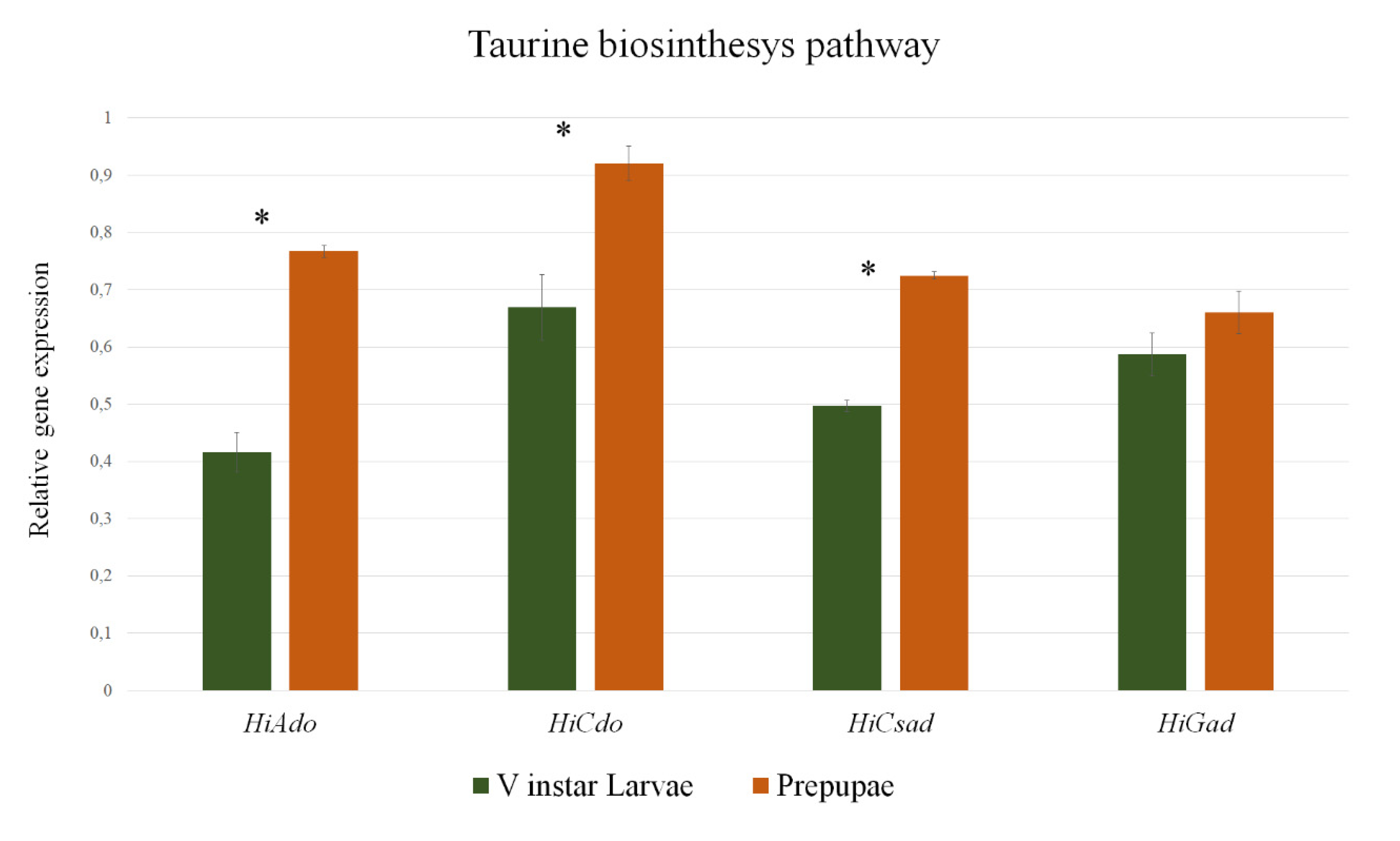
3.5. Quantitative PCR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Spranghers, T.; Ottoboni, M.; Klootwijk, C.; Ovyn, A.; Deboosere, S.; De Meulenaer, B.; Michiels, J.; Eeckhout, M.; De Clercq, P.; De Smet, S. Nutritional composition of black soldier fly (Hermetia illucens) prepupae reared on different organic waste substrates. J. Sci. Food Agric. 2017, 97, 2594–2600. [Google Scholar] [CrossRef] [PubMed]
- Van Huis, A.; Van Itterbeeck, J.; Klunder, H.; Mertens, E.; Halloran, A.; Muir, G.; Vantomme, P. Edible Insects —Future Prospects for Food and Feed Security; Forestry Paper 171; FAO (Food and Agriculture Organization of the United Nations): Rome, Italy, 2013. [Google Scholar]
- Makkar, H.P.S.; Tran, G.; Heuzé, V.; Ankers, P. State-of-the-art on use of insects as animal feed. Anim. Feed Sci. Tech. 2014, 97, 1–33. [Google Scholar] [CrossRef]
- Kroeckel, S.; Harjes, A.G.E.; Roth, I.; Katz, H.; Wuertz, S.; Susenbeth, A.; Schulz, C. When a turbot catches a fly: Evaluation of a pre-pupae meal of the Black Soldier Fly (Hermetia illucens) as fish meal substitute—Growth performance and chitin degradation in juvenile turbot (Psetta maxima). Aquaculture 2012, 364, 345–352. [Google Scholar] [CrossRef]
- Veldkamp, T.; Bosch, G. Insects: A protein rich feed ingredient in pig and poultry diets. Anim. Front. 2015, 5, 45–50. [Google Scholar] [CrossRef]
- De Marco, M.; Martínez, S.; Hernandez, F.; Madrid, J.; Gai, F.; Rotolo, L.; Belforti, M.; Bergero, D.; Katz, H.; Dabbou, S.; et al. Nutritional value of two insect larval meals (Tenebrio molitor and Hermetia illucens) for broiler chickens: Apparent nutrient digestibility, apparent ileal amino acid digestibility and apparent metabolizable energy. Anim. Feed Sci. Tech. 2015, 209, 211–218. [Google Scholar] [CrossRef]
- Tomberlin, J.K.; van Huis, A. Black soldier fly from pest to ‘crown jewel’ of the insects as feed industry: An historical perspective. J. Insects Food Feed 2020, 6, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Henry, M.; Gasco, L.; Piccolo, G.; Fountoulaki, E. Review on the use of insects in the diet of farmed fish: Past and future. Anim. Feed Sci. Tech. 2015, 203, 1–22. [Google Scholar] [CrossRef]
- St-Hilaire, S.; Cranfill, K.; McGuire, M.A.; Mosley, E.E.; Tomberlin, J.K.; Newton, L.; Sealey, W.; Sheppard, C.; Irving, S. Fish offal recycling by the black soldier fly produces a foodstuff high in omega-3 fatty acids. J. World Aquac. Soc. 2007, 38, 309–313. [Google Scholar] [CrossRef]
- Cardinaletti, G.; Randazzo, B.; Messina, M.; Zarantoniello, M.; Giorgini, E.; Zimbelli, A.; Bruni, L.; Parisi, G.; Olivotto, I.; Tulli, F. Effects of Graded Dietary Inclusion Level of Full-Fat Hermetia illucens Prepupae Meal in Practical Diets for Rainbow Trout (Oncorhynchus mykiss). Animals 2019, 9, 251. [Google Scholar] [CrossRef] [Green Version]
- Vargas, A.; Randazzo, B.; Riolo, P.; Truzzi, C.; Gioacchini, G.; Giorgini, E.; Loreto, N.; Ruschioni, S.; Zarantoniello, M.; Antonucci, M.; et al. Rearing Zebrafish on Black Soldier Fly (Hermetia illucens): Biometric, Histological, Spectroscopic, Biochemical, and Molecular Implications. Zebrafish 2018, 15, 404–419. [Google Scholar] [CrossRef]
- Zarantoniello, M.; Bruni, L.; Randazzo, B.; Vargas, A.; Gioacchini, G.; Truzzi, C.; Annibaldi, A.; Riolo, P.; Parisi, G.; Cardinaletti, G.; et al. Partial Dietary Inclusion of Hermetia illucens (Black Soldier Fly) Full-Fat Prepupae in Zebrafish Feed: Biometric, Histological, Biochemical, and Molecular Implications. Zebrafish 2018, 15, 519–532. [Google Scholar] [CrossRef] [PubMed]
- Zarantoniello, M.; Randazzo, B.; Truzzi, C.; Giorgini, E.; Marcellucci, C.; Vargas-Abúndez, J.A.; Zimbelli, A.; Annibaldi, A.; Parisi, G.; Tulli, F.; et al. A six-months study on Black Soldier Fly (Hermetia illucens) based diets in zebrafish. Sci. Rep. 2019, 9, 8598. [Google Scholar] [CrossRef] [PubMed]
- Salze, G.P.; Davis, D.A. Taurine: A critical nutrient for future fish feeds. Aquaculture 2015, 437, 215–229. [Google Scholar] [CrossRef]
- Gaylord, T.; Barrows, F.; Teague, A.; Johansen, K.; Overturf, K.; Shepherd, B. Supplementation of taurine and methionine to all-plant protein diets for rainbow trout (Oncorhynchus mykiss). Aquaculture 2007, 269, 514–524. [Google Scholar] [CrossRef]
- Kim, S.K.; Matsunari, H.; Nomura, K.; Tanaka, H.; Yokoyama, M.; Murata, Y.; Ishihara, K.; Takeuchi, T. Effect of dietary taurine and lipid contents on conjugated bile acid composition and growth performance of juvenile Japanese flounder Paralichthys olivaceus. Fish. Sci. 2008, 74, 875–881. [Google Scholar] [CrossRef]
- Wang, Q.; He, G.; Wang, X.; Mai, K.; Xu, W.; Zhou, H. Dietary sulfur amino acid modulations of taurine biosynthesis in juvenile turbot (Psetta maxima). Aquaculture 2014, 422, 141–145. [Google Scholar] [CrossRef]
- Kim, Y.S.; Sasaki, T.; Awa, M.; Inomata, M.; Honryo, T.; Agawa, Y.; Ando, M.; Sawada, Y. Effect of dietary taurine enhancement on growth and development in red sea bream Pagrus major larvae. Aquac. Res. 2016, 47, 1168–1179. [Google Scholar] [CrossRef]
- Espe, M.; Ruohonen, K.; El-Mowafi, A. Effect of taurine supplementation on the metabolism and body lipid-to-protein ratio in juvenile Atlantic salmon (Salmo salar). Aquac. Res. 2012, 271, 349–360. [Google Scholar] [CrossRef]
- Liu, P.; Ge, X.; Ding, H.; Jiang, H.; Christensen, B.M.; Li, J. Role of Glutamate Decarboxylase-like Protein 1 (GADL1) in Taurine Biosynthesis. J. Biol. Chem. 2012, 287, 40898–40906. [Google Scholar] [CrossRef] [Green Version]
- Finke, M.D. Complete nutrient content of four species of commercially available feeder insects fed enhanced diets during growth. Zoo Biol. 2015, 34, 554–564. [Google Scholar] [CrossRef]
- McCusker, S.; Buff, P.R.; Yu, Z.; Fascetti, A.J. Amino acid content of selected plant, algae and insect species: A search for alternative protein sources for use in pet foods. J. Nutr. Sci. 2014, 3, e39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gobbi, P.; Martínez-Sánchez, A.; Rojo, S. The effects of larval diet on adult life-history traits of the black soldier fly, Hermetia illucens (Diptera: Stratiomyidae). Eur. J. Entomol. 2013, 110, 461–468. [Google Scholar] [CrossRef]
- Liu, X.; Chen, X.; Wang, H.; Yang, Q.; Ur Rehman, K.; Li, W.; Cai, M.; Li, Q.; Mazza, L.; Zhang, J.; et al. Dynamic changes of nutrient composition throughout the entire life cycle of black soldier fly. PLoS ONE 2017, 12, e0182601. [Google Scholar] [CrossRef] [Green Version]
- Rachmawati, R.; Buchori, D.; Hidayat, P.; Hem, S.; Fahmi, M.R. Perkembangandan Kandungan Nutrisi Larva Hermetia illucens (Linnaeus) (Diptera: Stratiomyidae) pada Bungkil Kelapa Sawit. J. Entomol. Indones. 2017, 7, 28–41. [Google Scholar] [CrossRef] [Green Version]
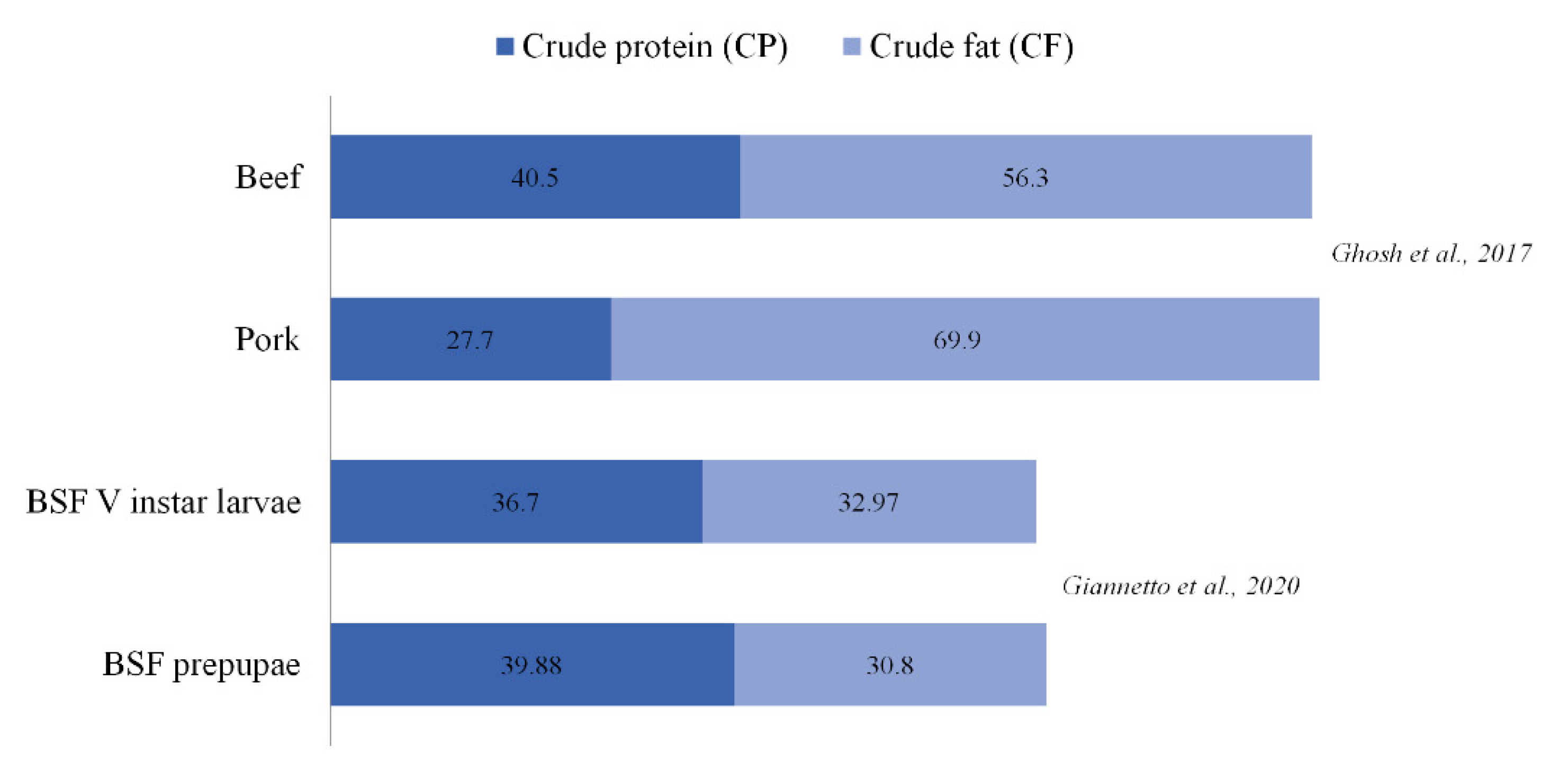
- Giannetto, A.; Oliva, S.; Ceccon Lanes, C.F.; de Araújo Pedron, F.; Savastano, D.; Baviera, C.; Parrino, V.; Lo Paro, G.; Spanò, N.; Cappello, T.; et al. Hermetia illucens (Diptera: Stratiomydae) larvae and prepupae: Biomass production, fatty acid profile and expression of key genes involved in lipid metabolism. J. Biotechnol. 2020, 307, 44–54. [Google Scholar] [CrossRef] [PubMed]
- Barragan-Fonseca, K.B.; Dicke, M.; van Loon, J.J.A. Nutritional value of the black soldier fly (Hermetia illucens L.) and its suitability as animal feed—A review. J. Insects Food Feed 2017, 3, 105–120. [Google Scholar] [CrossRef]
- Kim, W.; Bae, S.; Park, H.; Park, K.; Lee, S.; Choi, Y.; Han, S.; Koh, Y.H. The Larval Age and Mouth Morphology of the Black Soldier Fly, Hermetia illucens (Diptera: Stratiomyidae). Int. J. Indust. Entomol. 2010, 21, 185–187. [Google Scholar]
- Tomberlin, J.K.; Adler, P.H.; Myers, H.M. Development of the black soldier fly (Diptera: Stratiomyidae) in relation to temperature. Environ. Entomol. 2009, 38, 930–934. [Google Scholar] [CrossRef]
- Finke, M.D. Complete Nutrient Content of Four Species of Feeder Insects. Zoo Biol. 2013, 32, 27–36. [Google Scholar] [CrossRef]
- Boyd, C.E. Feed efficiency indicators for responsible aquaculture. Glob. Aquac. Advocate 2005, 8, 73–74. [Google Scholar]
- Barroso, F.G.; de Haro, C.; Sánchez-Muros, M.J.; Venegas, E.; Martínez-Sánchez, A.; Pérez-Bañón, C. The potential of various insect species for use as food for fish. Aquaculture 2014, 422, 193–201. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis of the Association of Official Analytical Chemists International, 18th ed.; AOAC International: Gaitherburg, MD, USA, 2005. [Google Scholar]
- Giannetto, A.; Oliva, S.; Mazza, L.; Mondello, G.; Savastano, D.; Mauceri, A.; Fasulo, S. Molecular characterization and expression analysis of heat shock protein 70 and 90 from Hermetia illucens reared in a food waste bioconversion pilot plant. Gene 2017, 627, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Lazado, C.C.; Kumaratunga, H.P.S.; Nagasawa, K.; Babiak, I.; Giannetto, A.; Fernandes, J.M.O. Daily rhythmicity of clock gene transcripts in Atlantic cod fast skeletal muscle. PLoS ONE 2014, 9, e99172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Giannetto, A.; Nagasawa, K.; Fasulo, S.; Fernandes, J.M.O. Influence of photoperiod on expression of DNA (cytosine-5) methyltransferases in Atlantic cod. Gene 2013, 519, 222–230. [Google Scholar] [CrossRef] [PubMed]
- Nagasawa, K.; Giannetto, A.; Fernandes, J.M.O. Photoperiod influences growth and MLL (mixed-lineage leukaemia) expression in Atlantic cod. PLoS ONE 2012, 7, e36908. [Google Scholar] [CrossRef] [Green Version]
- Lalander, C.; Diener, S.; Zurbrügg, C.; Vinneras, B. Effects of feedstock on larval development and process efficiency in waste treatment with black soldier fly (Hermetia illucens). J. Clean. Prod. 2019, 208, 211–219. [Google Scholar] [CrossRef]
- Davis, D.; Nguyen, T.; Li, M.; Gatlin, D.M.; O’Keefe, T. Advances in aquaculture nutrition: Catfish, tilapia and carp nutrition. In New Technologies in Aquaculture; Burnell, G., Allan, G., Eds.; Woodhead Publishing: Witney/Oxford, UK, 2009; pp. 440–458. [Google Scholar] [CrossRef]
- Ghosh, S.; Lee, S.M.; Jung, C.; Meyer-Rochow, V.B. Nutritional composition of five commercial edible insects in South Korea. J. Asia Pac. Entomol. 2017, 20, 686–694. [Google Scholar] [CrossRef]
- Directive 2002/32/EC of the European Parliament and of the Council of 7 May 2002 on undesirable substances in animal feed. Off. J. Eur. Comm. 2002, OJ L 140, 10.
- Barroso, F.G.; Sánchez-Muros, M.J.; Segura, M.; Morote, E.; Torres, A.; Ramos, R.; Guil, J.L. Insects as food: Enrichment of larvae of Hermetia illucens with omega 3 fatty acids by means of dietary modifications. J. Food Compos. Anal. 2017, 62, 8–13. [Google Scholar] [CrossRef]
- Freccia, A.; Tubin, J.; Rombenso, A.; Emerenciano, M. Insects in Aquaculture Nutrition: An Emerging Eco-Friendly Approach or Commercial Reality? In Emerging Technologies and Research for Eco-Friendly Aquaculture; Lu, Q., Serajuddin, M., Eds.; IntechOpen: London, UK, 2020; pp. 1–14. [Google Scholar] [CrossRef] [Green Version]
- Oonincx, D.G.A.B.; Van Broekhoven, S.; Van Huis, A.; Van Loon, J.J.A. Feed conversion, survival and development, and composition of four insect species on diets composed of food by-products. PLoS ONE 2015, 10, e0144601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez-Muros, M.J.; Barroso, F.G.; Manzano-Agugliaro, F. Insect meal as renewable source of food for animal feeding: A review. J. Clean. Prod. 2014, 65, 16–27. [Google Scholar] [CrossRef]
- Van Huis, A. Potential of insects as food and feed in assuring food security. Annu. Rev. Entomol. 2013, 58, 563–583. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.S.; Shelomi, M. Review of Black Soldier Fly (Hermetia illucens) as Animal Feed and Human Food. Foods 2017, 6, 91. [Google Scholar] [CrossRef] [Green Version]
- EFSA Scientific Committee. Risk profile related to production and consumption of insects as food and feed. EFSA J. 2015, 13, 4257. [Google Scholar] [CrossRef] [Green Version]
- Jucker, C.; Erba, D.; Leonardi, M.G.; Lupi, D.; Savoldelli, S. Assessment of Vegetable and Fruit Substrates as Potential Rearing Media for Hermetia illucens (Diptera: Stratiomyidae) Larvae. Environ. Entomol. 2017, 46, 1415–1423. [Google Scholar] [CrossRef]
- Nguyen, T.T.X.; Tomberlin, J.K.; Vanlaerhoven, S. Ability of Black Soldier Fly (Diptera: Stratiomyidae) Larvae to Recycle Food Waste. Environ. Entomol. 2015, 44, 406–410. [Google Scholar] [CrossRef]
- Nyakeri, E.M.; Ogola, H.J.O.; Ayieko, M.A.; Amimo, F.A. Valorisation of organic waste material: Growth performance of wild black soldier fly larvae (Hermetia illucens) reared on different organic wastes. J. Insects Food Feed 2017, 3, 193–202. [Google Scholar] [CrossRef]
- Sheppard, C.D.; Newton, L.G.; Sidney, T.A.; Stan, S. A value added manure management system using the black soldier fly. Bioresource Technol. 1994, 50, 275–279. [Google Scholar] [CrossRef]
- Salomone, R.; Saija, G.; Mondello, G.; Giannetto, A.; Fasulo, S.; Savastano, D. Environmental impact of food waste bioconversion by insects: Application of Life Cycle Assessment to process using Hermetia illucens. J. Clean. Prod. 2017, 140, 890–905. [Google Scholar] [CrossRef]
- Diener, S.; Zurbrügg, C.; Tockner, K. Conversion of organic material by black soldier fly larvae: Establishing optimal feeding rates. Waste Manag. Res. 2009, 27, 603–610. [Google Scholar] [CrossRef] [PubMed]
- Meneguz, M.; Schiavone, A.; Gai, F.; Dama, A.; Lussiana, C.; Renna, M.; Gasco, L. Effect of rearing substrate on growth performance, waste reduction efficiency and chemical composition of black soldier fly (Hermetia illucens) larvae. J. Sci. Food Agric. 2018, 98, 5776–5784. [Google Scholar] [CrossRef]
- Li, Q.; Zheng, L.; Qiu, N.; Cai, H.; Tomberlin, J.K. Bioconversion of dairy manure by black soldier fly (Diptera: Stratiomyidae) for biodiesel and sugar production. Waste Manag. 2011, 31, 1316–1320. [Google Scholar] [CrossRef] [PubMed]
- Leong, S.Y.; Kutty, S.R.M.; Malakahmad, A.; Tan, C.K. Feasibility study of biodiesel production using lipids of Hermetia illucens larva fed with organic waste. Waste Manag. 2016, 47, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Surendra, K.C.; Olivier, R.; Tomberlin, J.K.; Jha, R.; Khanal, S.K. Bioconversion of organic wastes into biodiesel and animal feed via insect farming. Renew. Energy 2016, 98, 197–202. [Google Scholar] [CrossRef] [Green Version]
- Zheng, L.; Li, Q.; Zhang, J.; Yu, Z. Double the biodiesel yield: Rearing black soldier fly larvae, Hermetia illucens, on solid residual fraction of restaurant waste after grease extraction for biodiesel production. Renew. Energy 2012, 41, 75–79. [Google Scholar] [CrossRef]
- Liu, C.; Wang, C.; Yao, H. Comprehensive resource utilization of waste using the Black Soldier Fly (Hermetia illucens (L.)) (Diptera: Stratiomyidae). Animals 2019, 9, 349. [Google Scholar] [CrossRef] [Green Version]
- Cappellozza, S.; Leonardi, M.G.; Savoldelli, S.; Carminati, D.; Rizzolo, A.; Cortellino, G.; Terova, G.; Moretto, E.; Badaile, A.; Concheri, G.; et al. A first attempt to produce proteins from insects by means of a circular economy. Animals 2019, 9, 278. [Google Scholar] [CrossRef] [Green Version]
- Müller, A.; Wolf, D.; Gutzeit, H.O. The black soldier fly, Hermetia illucens—A promising source for sustainable production of proteins, lipids and bioactive substances. Z. Nat. 2017, 72, 351–363. [Google Scholar] [CrossRef]
- Huyben, D.; Vidaković, A.; Werner Hallgren, S.; Langeland, M. High-throughput sequencing of gut microbiota in rainbow trout (Oncorhynchus mykiss) fed larval and pre-pupae stages of black soldier fly (Hermetia illucens). Aquaculture 2019, 500, 485–491. [Google Scholar] [CrossRef]
- Sealey, W.M.; Gaylord, T.G.; Barrows, F.T.; Tomberlin, J.K.; McGuire, M.A.; Ross, C.; St-Hilaire, S. Sensory Analysis of Rainbow Trout, Oncorhynchus mykiss, Fed Enriched Black Soldier Fly Prepupae, Hermetia illucens. J. World Aquac. Soc. 2011, 42, 34–45. [Google Scholar] [CrossRef]
- Bicker, G. Taurine in the insect central nervous system. Comp. Biochem. Physiol. C Comp. Pharmacol. 1992, 103, 423–428. [Google Scholar] [CrossRef]
- Whitton, P.S.; Nicholson, R.A.; Bell, M.F.; Strang, R.H.C. Biosynthesis of taurine in tissues of the locust (Schistocerca americana gregaria) and the effect of physiological and toxicological stresses on biosynthetic rate of this amino acid. Insect Biochem. Mol. Biol. 1995, 25, 83–87. [Google Scholar] [CrossRef]
- Rumpold, B.A.; Schluter, O.K. Nutritional composition and safety aspects of edible insects. Mol. Nutr. Food Res. 2013, 57, 802–823. [Google Scholar] [CrossRef] [PubMed]
- Adamse, P.; Van der Fels-Klerx, H.J.I.; de Jong, J. Cadmium, lead, mercury and arsenic in animal feed and feed materials—Trend analysis of monitoring results. Food Addit. Contam. Part A Chem. Anal. Control. Expo. Risk Assess. 2017, 34, 1298–1311. [Google Scholar] [CrossRef]
- Diener, S.; Zurbrügg, C.; Tockner, K. Bioaccumulation of heavy metals in the black soldier fly, Hermetia illucens and effects on its life cycle. J. Insects Food Feed 2015, 1, 261–270. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amino Acid (%) | V Instar Larvae | Prepupae | Edible Insects * | Recommended Minimum Amino Acid Levels in Fish Diets # |
|---|---|---|---|---|
| Essential | ||||
| Argininef | 6.49 ± 0.2 a | 8.87 ± 0.5 b | 2.09–3.71 | 4.2–4.3 |
| Histidinehf | 7.49 ± 0.4 a | 8.12 ± 0.7 b | 1.8–2.8 | 1.5–2.1 |
| Isoleucinehf | 1.71 ± 0.02 a | 1.37 ± 0.07 a | 1.62–2.16 | 2.5–3.1 |
| Leucinehf | 2.25 ± 0.1 a | 1.79 ± 0.02 b | 2.3–3.97 | 3.3–3.4 |
| Lysine hf | 3.65 ± 0.2 a | 3.35 ± 0.2 a | 1.7–2.6 | 5.1–5.7 |
| Methioninehf | 0.21 ± 0.04 a | 0.19 ± 0.1 a | 0–0.3 | |
| Methionine and cystine | 2.3–3.2 | |||
| Phenylalaninehf | 1.28 ± 0.06 a | 1.03 ± 0.09 a | 1.62–1.83 | |
| Phenylalanine and tyrosine | 5–6.5 | |||
| Threoninehf | 2.49 ± 0.3 a | 2.29 ± 0.07 a | 1.5–2 | 2–3.9 |
| Tyrosine h | 0.73 ± 0.06 a | 0.92 ± 0.5 a | 2.6–3.7 | |
| Valinehf | 3.04 ± 0.2 a | 2.59 ± 0.06 b | 2.7–3.2 | 2.8–3.6 |
| Non-essential | ||||
| Alanine | 16.49 ± 0.02 a | 11.67 ± 0.3 b | ||
| Aspartic acid | 4.08 ± 0.4 a | 2.72 ± 0.08 b | ||
| Glutamic acid | 21.76 ± 0.09 a | 17.74 ± 0.05 b | ||
| Glycine | 2.08 ± 0.3 a | 2.02 ± 0.2 a | ||
| Serine | 2.51 ± 0.07 a | 1.80 ± 0.06 b | ||
| Others | 23.94 ± 0.1 a | 32.95 ± 0.05 b | ||
| EAA | 29.34 ± 0.2 a | 30.52 ± 0.02 b | ||
| NEAA | 46.92 ± 0.3 a | 35.95 ± 0.06 b | ||
| Taurine (mg/Kg) | 24 ± 0.02 a | 45 ± 0.01 b |
| Mineral | V Instar Larvae | Prepupae | EU Directive 2002/32/EC (mg/kg) |
|---|---|---|---|
| Ca | 0.72 | 0.92 | - |
| P | 0.58 | 0.71 | - |
| K | 0.79 | 0.84 | - |
| Mg | 0.26 | 0.33 | - |
| Fe | 84 | 89 | - |
| Zn | 68 | 98 | - |
| Mn | 212 | 268 | - |
| As | 1.01 | 0.96 | 2 |
| Cd | 0.37 | 0.24 | 2 |
| Hg | <0.05 | <0.05 | 0.1 |
| Pb | 0.39 | 0.32 | 10 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giannetto, A.; Oliva, S.; Riolo, K.; Savastano, D.; Parrino, V.; Cappello, T.; Maisano, M.; Fasulo, S.; Mauceri, A. Waste Valorization via Hermetia Illucens to Produce Protein-Rich Biomass for Feed: Insight into the Critical Nutrient Taurine. Animals 2020, 10, 1710. https://doi.org/10.3390/ani10091710
Giannetto A, Oliva S, Riolo K, Savastano D, Parrino V, Cappello T, Maisano M, Fasulo S, Mauceri A. Waste Valorization via Hermetia Illucens to Produce Protein-Rich Biomass for Feed: Insight into the Critical Nutrient Taurine. Animals. 2020; 10(9):1710. https://doi.org/10.3390/ani10091710
Chicago/Turabian StyleGiannetto, Alessia, Sabrina Oliva, Kristian Riolo, Domenico Savastano, Vincenzo Parrino, Tiziana Cappello, Maria Maisano, Salvatore Fasulo, and Angela Mauceri. 2020. "Waste Valorization via Hermetia Illucens to Produce Protein-Rich Biomass for Feed: Insight into the Critical Nutrient Taurine" Animals 10, no. 9: 1710. https://doi.org/10.3390/ani10091710