Estradiol-17β Pharmacokinetics and Histological Assessment of the Ovaries and Uterine Horns following Intramuscular Administration of Estradiol Cypionate in Feral Cats

 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Ethics Statement
2.2. Study Animals
2.3. Housing
2.4. Trial
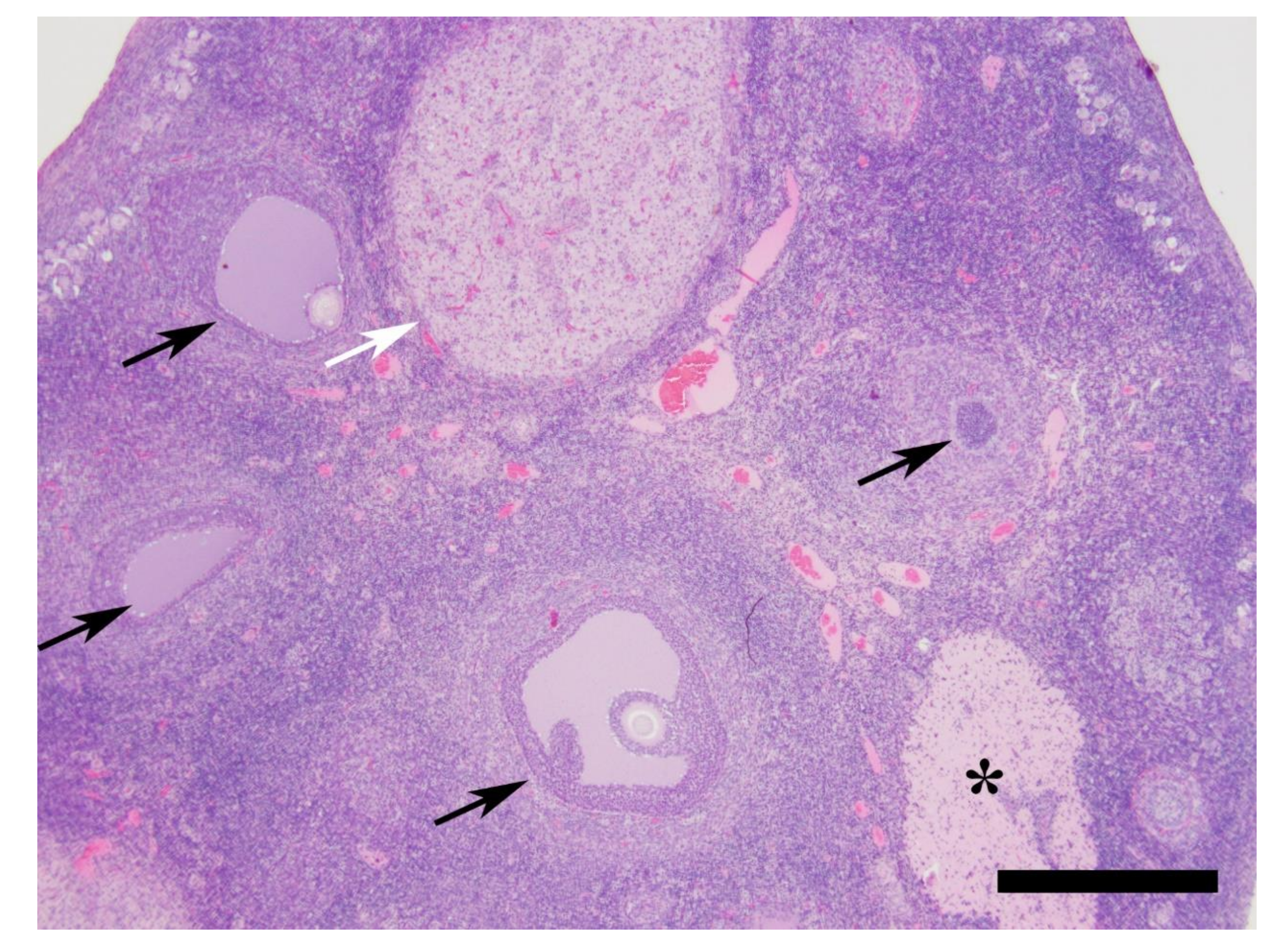
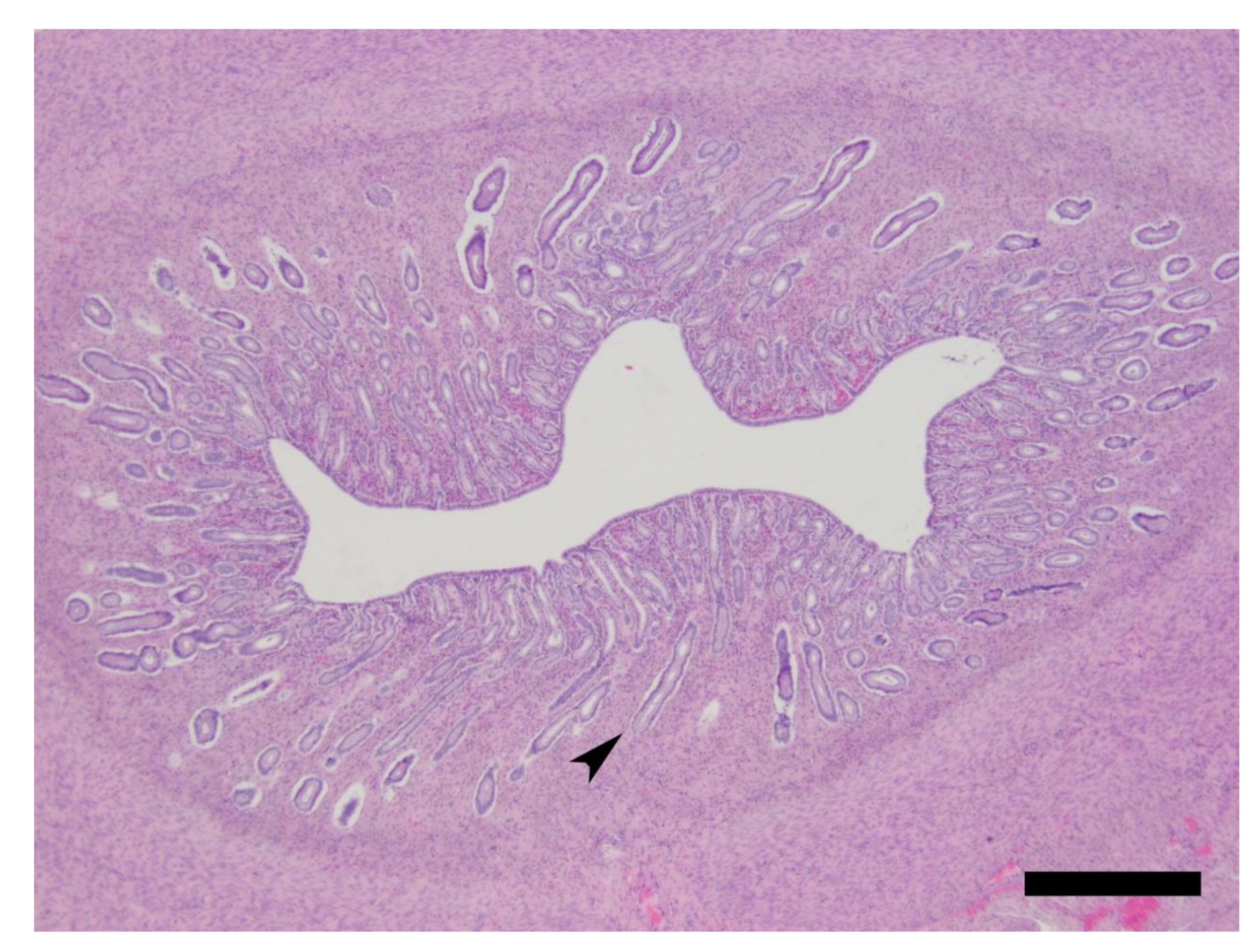
2.5. Histology
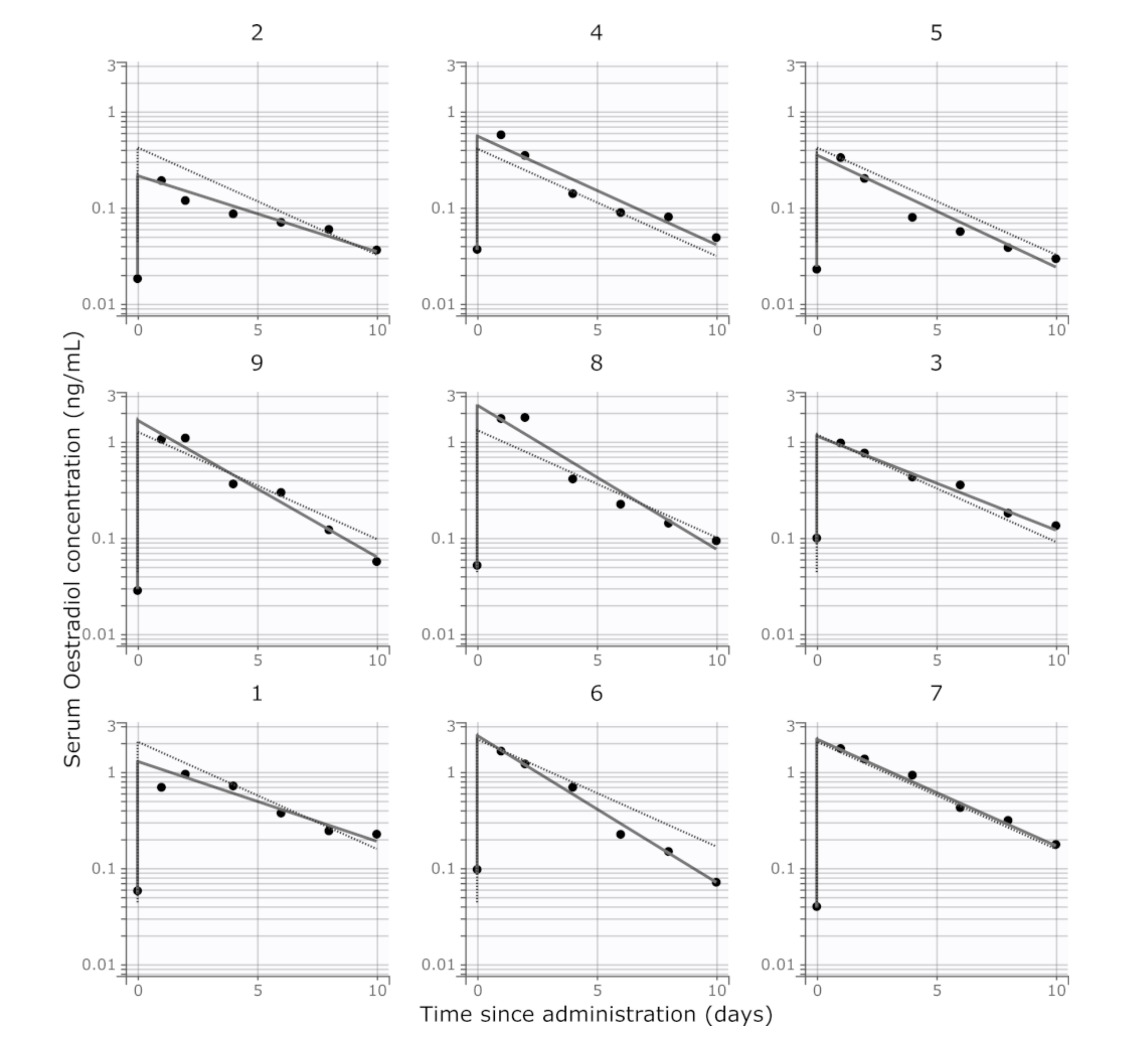
2.6. Non-Compartmental Pharmacokinetic Calculations
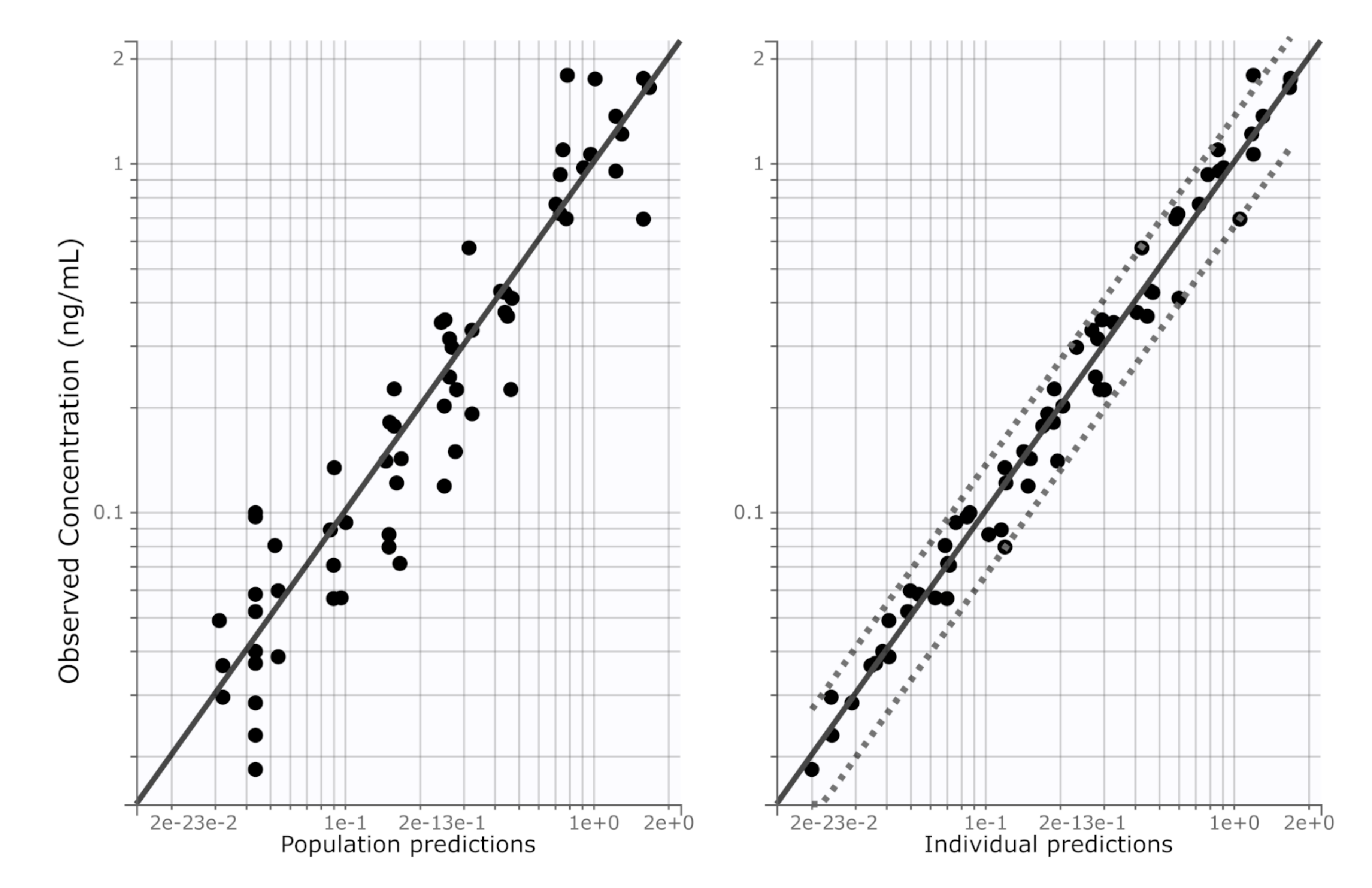
2.7. Pharmacokinetic Modelling
- ka: Absorption rate constant
- VC/F: Central compartment volume of distribution
- C1: Difference between absorption and elimination (ke) rate constants
- BX: Baseline estradiol-17β concentration
- σε: Residual error standard deviation
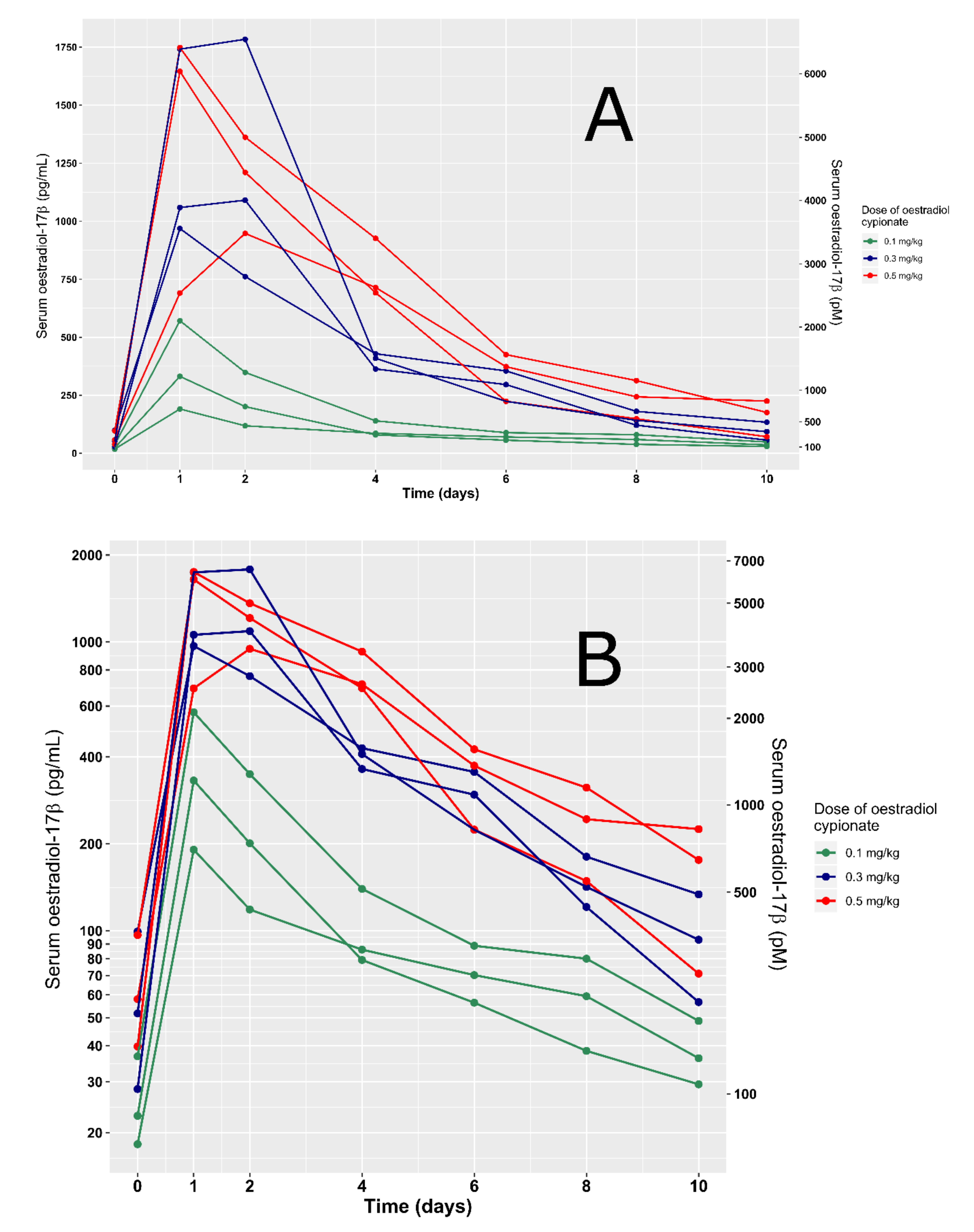
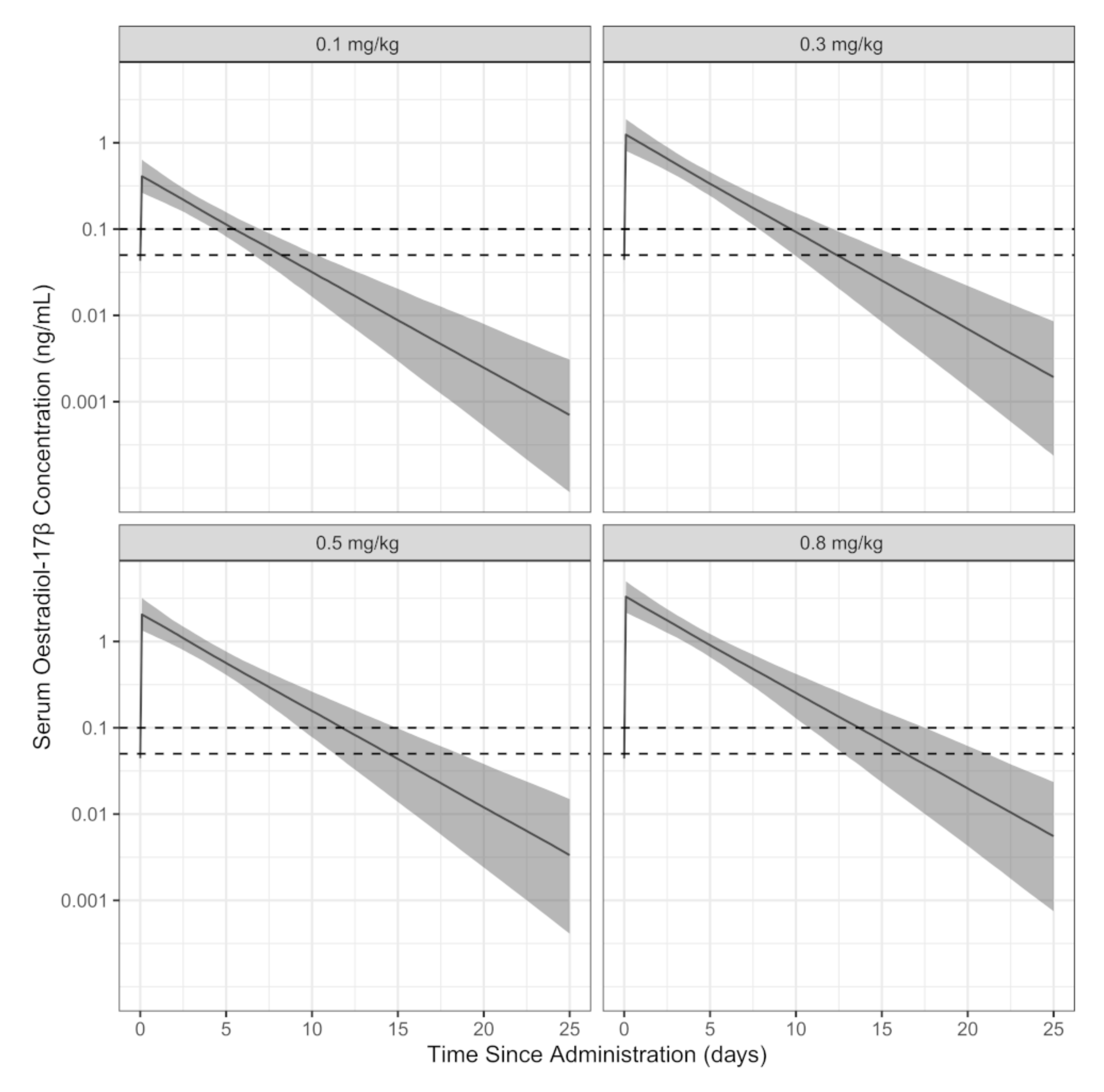
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Woinarski, J.C.; Burbidge, A.A.; Harrison, P.L. Ongoing unraveling of a continental fauna: Decline and extinction of Australian mammals since European settlement. Proc. Natl. Acad. Sci. USA 2015, 112, 4531–4540. [Google Scholar] [CrossRef] [Green Version]
- Short, J.; Turner, B.; Risbey, D.A.; Carnamah, R. Control of feral cats for nature conservation. II. Population reduction by poisoning. Wildl. Res. 1997, 24, 703–714. [Google Scholar] [CrossRef]
- Algar, D.; Burbidge, A.; Angus, G. Cat eradication on Hermite Island, Montebello Islands, Western Australia. In Turning the Tide: The Eradication of Invasive Species; Veitch, C.R., Clout, M.N., Eds.; Invasive Species Specialist Group of the World Conservation Union (IUCN): Auckland, New Zealand, 2002; pp. 14–18. [Google Scholar]
- Algar, D.; Angus, G.; Onus, M. Eradication of feral cats on Rottnest Island, Western Australia. J. R. Soc. West. Aust. 2011, 94, 439–443. [Google Scholar]
- Edwards, G.; Piddington, K.; Paltridge, R. Field evaluation of olfactory lures for feral cats (Felis catus L.) in central Australia. Wildl. Res. 1997, 24, 173–183. [Google Scholar] [CrossRef]
- Spencer, P.B.; Hampton, J.O.; Pacioni, C.; Kennedy, M.S.; Saalfeld, K.; Rose, K.; Woolnough, A.P. Genetic relationships within social groups influence the application of the Judas technique: A case study with wild dromedary camels. J. Wildl. Manag. 2015, 79, 102–111. [Google Scholar] [CrossRef] [Green Version]
- Woolnough, A.P.; Hampton, J.O.; Campbell, S.; Lethbridge, M.R.; Boardman, W.S.; Sharp, T.; Rose, K. Field immobilization of feral ‘Judas’ donkeys (Equus asinus) by remote injection of medetomidine and ketamine and antagonism with atipamezole. J. Wildl. Dis. 2012, 48, 435–443. [Google Scholar] [CrossRef] [Green Version]
- McIlroy, J.; Gifford, E. The ‘Judas’ pig technique: A method that could enhance control programmes against feral pigs, Sus scrofa. Wildl. Res. 1997, 24, 483–491. [Google Scholar] [CrossRef]
- Taylor, D.; Katahira, L. Radio telemetry as an aid in eradicating remnant feral goats. Wildl. Soc. Bull. 1988, 16, 297–299. [Google Scholar]
- Campbell, K.J.; Baxter, G.S.; Murray, P.J.; Coblentz, B.E.; Donlan, C.J. Development of a prolonged estrus effect for use in Judas goats. Appl. Anim. Behav. Sci. 2007, 102, 12–23. [Google Scholar] [CrossRef]
- Carrion, V.; Donlan, C.J.; Campbell, K.J.; Lavoie, C.; Cruz, F. Archipelago-wide island restoration in the Galápagos Islands: Reducing costs of invasive mammal eradication programs and reinvasion risk. PLoS ONE 2011, 6. [Google Scholar] [CrossRef]
- Michael, R.; Scott, P.P. The activation of sexual behaviour in cats by the subcutaneous administration of oestrogen. J. Physiol. 1964, 171, 254–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, G.; Michael, R. The activation of sexual behaviour by hypothalamic implants of oestrogen. J. Physiol. 1964, 171, 275–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hart, B.L. Facilitation by estrogen of sexual reflexes in female cats. Physiol. Behav. 1971, 7, 675–678. [Google Scholar] [CrossRef]
- Beyer, C.; Morali, G.; Larsson, K.; Södersten, P. Steroid regulation of sexual behavior. J. Steroid Biochem. 1976, 7, 1171–1176. [Google Scholar] [CrossRef]
- Johnson, L.M.; Gay, V.L. Luteinizing hormone in the cat. II. Mating-induced secretion. Endocrinology 1981, 109, 247–252. [Google Scholar] [CrossRef]
- Robison, B.L.; Sawyer, C.H. Hypothalamic control of ovulation and behavioral estrus in the cat. Brain Res. 1987, 418, 41–51. [Google Scholar] [CrossRef]
- Quadagno, D.M.; McCullough, J.; Langan, R. The effect of varying amounts of exogenous estradiol benzoate on estrous behavior in the rat. Horm. Behav. 1972, 3, 175–179. [Google Scholar] [CrossRef]
- Dial, G.; Dial, O.; Bevier, G.; Glenn, S.; Dziuk, P. Estrous behavior and circadian discharge of luteinizing hormone in the prepubertal gilt in response to exogenous estrogen. Biol. Reprod. 1983, 29, 1047–1056. [Google Scholar] [CrossRef] [Green Version]
- McCarthy, M.M.; Albrecht, E.D. Steroid regulation of sexual behavior. Trends Endocrinol. Metab. 1996, 7, 324–327. [Google Scholar] [CrossRef]
- Lindsay, D. Modification of behavioural oestrus in the ewe by social and hormonal factors. Anim. Behav. 1966, 14, 73–83. [Google Scholar] [CrossRef]
- Peretz, E. Estrogen dose and the duration of the mating period in cats. Physiol. Behav. 1968, 3, 41–43. [Google Scholar] [CrossRef]
- Kuhl, H. Pharmacology of estrogens and progestogens: Influence of different routes of administration. Climacteric 2005, 8, 3–63. [Google Scholar] [CrossRef] [PubMed]
- Papich, M.G. Saunders Handbook of Veterinary Drugs: Small and Large Animal, 4th ed.; Elsevier Health Sciences: St. Louis, MO, USA, 2016. [Google Scholar]
- Plumb, D.C. Plumb’s Veterinary Drug Handbook, 7th ed.; PharmaVet Inc.: Stockholm, WI, USA, 2011. [Google Scholar]
- Oriowo, M.; Landgren, B.-M.; Stenström, B.; Diczfalusy, E. A comparison of the pharmacokinetic properties of three estradiol esters. Contraception 1980, 21, 415–424. [Google Scholar] [PubMed]
- Cavilla, M.V.; Bianchi, C.P.; Aba, M.A. Oestradiol-17β plasma concentrations after intramuscular injection of oestradiol benzoate or oestradiol cypionate in llamas (Lama glama). Acta Vet. Scand. 2010, 52, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuhl, H. Pharmacokinetics of oestrogens and progestogens. Maturitas 1990, 12, 171–197. [Google Scholar] [CrossRef]
- Shille, V.; Haggerty, M.; Shackleton, C.; Lasley, B. Metabolites of estradiol in serum, bile, intestine and feces of the domestic cat (Felis catus). Theriogenology 1990, 34, 779–794. [Google Scholar] [CrossRef]
- Hart, B.L.; Leedy, M.G. Female sexual responses in male cats facilitated by olfactory bulbectomy and medial preoptic/anterior hypothalamic lesions. Behav. Neurosci. 1983, 97, 608. [Google Scholar] [CrossRef]
- Herron, M.; Sis, R. Ovum transport in the cat and the effect of estrogen administration. Am. J. Vet. Res. 1974, 35, 1277. [Google Scholar]
- Hart, J. Endocrine pathology of estrogens: Species differences. Pharmacol. Ther. 1990, 47, 203–218. [Google Scholar] [CrossRef]
- McGregor, H.; Legge, S.; Jones, M.E.; Johnson, C.N. Feral cats are better killers in open habitats, revealed by animal-borne video. PLoS ONE 2015, 10. [Google Scholar] [CrossRef] [Green Version]
- Molsher, R. Trapping and demographics of feral cats (Felis catus) in central New South Wales. Wildl. Res. 2001, 28, 631–636. [Google Scholar] [CrossRef]
- Coucke, W.; Devleeschouwer, N.; Libeer, J.-C.; Schiettecatte, J.; Martin, M.; Smitz, J. Accuracy and reproducibility of automated estradiol-17 β and progesterone assays using native serum samples: Results obtained in the Belgian external assessment scheme. Hum. Reprod. 2007, 22, 3204–3209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reynaud, K.; Gicquel, C.; Thoumire, S.; Chebrout, M.; Ficheux, C.; Bestandji, M.; Chastant-Maillard, S. Folliculogenesis and morphometry of oocyte and follicle growth in the feline ovary. Reprod. Domest. Anim. 2009, 44, 174–179. [Google Scholar] [CrossRef] [PubMed]
- Amelkina, O.; Braun, B.C.; Dehnhard, M.; Jewgenow, K. The corpus luteum of the domestic cat: Histologic classification and intraluteal hormone profile. Theriogenology 2015, 83, 711–720. [Google Scholar] [CrossRef] [PubMed]
- Chatdarong, K.; Rungsipipat, A.; Axnér, E.; Forsberg, C.L. Hysterographic appearance and uterine histology at different stages of the reproductive cycle and after progestagen treatment in the domestic cat. Theriogenology 2005, 64, 12–29. [Google Scholar] [CrossRef]
- Toutain, P.-L.; Bousquet-Mélou, A. Volumes of distribution. J. Vet. Pharmacol. Ther. 2004, 27, 441–453. [Google Scholar] [CrossRef]
- Bon, C.; Toutain, P.; Concordet, D.; Gehring, R.; Martin-Jimenez, T.; Smith, J.; Pelligand, L.; Martinez, M.; Whittem, T.; Riviere, J. Mathematical modeling and simulation in animal health. Part III: Using nonlinear mixed-effects to characterize and quantify variability in drug pharmacokinetics. J. Vet. Pharmacol. Ther. 2018, 41, 171–183. [Google Scholar] [CrossRef] [Green Version]
- Yáñez, J.A.; Remsberg, C.M.; Sayre, C.L.; Forrest, M.L.; Davies, N.M. Flip-flop pharmacokinetics–delivering a reversal of disposition: Challenges and opportunities during drug development. Ther. Deliv. 2011, 2, 643–672. [Google Scholar] [CrossRef] [Green Version]
- Shille, V.; Lundström, K.E.; Stabenfeldt, G. Follicular function in the domestic cat as determined by estradiol-17 β concentrations in plasma: Relation to estrous behavior and cornification of exfoliated vaginal epithelium. Biol. Reprod. 1979, 21, 953–963. [Google Scholar] [CrossRef] [Green Version]
- Verhage, H.; Beamer, N.; Brenner, R. Plasma levels of estradiol and progesterone in the cat during polyestrus, pregnancy and pseudopregnancy. Biol. Reprod. 1976, 14, 579–585. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Vynckier, L.; Debackere, M.; De Kruif, A.; Coryn, M. Plasma estradiol-17ß concentrations in the cow during induced estrus and after injection of estradiol-17ß benzoate and estradiol-17ß cypionate-a preliminary study. J. Vet. Pharmacol. Ther. 1990, 13, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Martins, R.S.; Antunes, N.J.; Comerlatti, G.; Caraccio, G.; Moreno, R.A.; Frecentese, F.; Caliendo, G.; De Nucci, G. Quantification of estradiol cypionate in plasma by liquid chromatography coupled with tandem mass spectrometry: Application in a pharmacokinetic study in healthy female volunteers. J. Pharm. Biomed. Anal. 2019, 170, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Lokind, K.B.; Lorenzen, F.H.; Bundgaard, H. Oral bioavailability of 17β-estradiol and various ester prodrugs in the rat. Int. J. Pharm. 1991, 76, 177–182. [Google Scholar] [CrossRef]
- Shaw, N.D.; Histed, S.N.; Srouji, S.S.; Yang, J.; Lee, H.; Hall, J.E. Estrogen negative feedback on gonadotropin secretion: Evidence for a direct pituitary effect in women. J. Clin. Endocrinol. Metab. 2010, 95, 1955–1961. [Google Scholar] [CrossRef] [Green Version]
- Clarke, I.J. Multifarious effects of estrogen on the pituitary gonadotrope with special emphasis on studies in the ovine species. Arch. Physiol. Biochem. 2002, 110, 62–73. [Google Scholar] [CrossRef]
- Johnson, L.M.; Gay, V.L. Luteinizing hormone in the cat. I. Tonic secretion. Endocrinology 1981, 109, 240–246. [Google Scholar] [CrossRef]
- Inselman-Temkin, B.R.; Flynn, J.P. Sex-dependent effects of gonadal and gonadotropic hormones on centrally-elicited attack in cats. Brain Res. 1973, 60, 393–410. [Google Scholar] [CrossRef]
- Larsen, B.; Markovetz, A.; Galask, R. Role of estrogen in controlling the genital microflora of female rats. Appl. Environ. Microbiol. 1977, 34, 534–540. [Google Scholar] [CrossRef] [Green Version]
- Grunert, G.; Porcia, M.; Tchernitchin, A. Differential potency of oestradiol-17β and diethylstilboestrol on separate groups of responses in the rat uterus. J. Endocrinol. 1986, 110, 103–114. [Google Scholar] [CrossRef]
- Jöchle, W.; Jöchle, M. Reproductive and behavioral control in the male and female cat with progestins: Long-term field observations in individual animals. Theriogenology 1975, 3, 179–185. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Animal Number/Population | Dose of Estradiol Cypionate (mg/kg) | Analyte Quantified in Assay | Cmax(ng/mL) | tmax (Day) | λz (/Day) | t1/2β (Day) | AUC0→∞ (ng.Day/mL) | Reference |
|---|---|---|---|---|---|---|---|---|
| 2 | 0.10 | Estradiol-17β | 0.191 | 1 | 0.137 | 5.06 | 1.11 | this study |
| 4 | 0.10 | Estradiol-17β | 0.571 | 1 | 0.163 | 4.25 | 2.08 | this study |
| 5 | 0.10 | Estradiol-17β | 0.331 | 1 | 0.168 | 4.13 | 1.20 | this study |
| 3 | 0.28 | Estradiol-17β | 0.969 | 1 | 0.217 | 3.19 | 4.84 | this study |
| 8 | 0.31 | Estradiol-17β | 1.784 | 2 | 0.245 | 2.83 | 6.47 | this study |
| 9 | 0.30 | Estradiol-17β | 1.091 | 2 | 0.351 | 1.98 | 4.49 | this study |
| 1 | 0.47 | Estradiol-17β | 0.947 | 2 | 0.198 | 3.51 | 6.17 | this study |
| 6 | 0.52 | Estradiol-17β | 1.646 | 1 | 0.360 | 1.92 | 5.91 | this study |
| 7 | 0.47 | Estradiol-17β | 1.748 | 1 | 0.259 | 2.68 | 8.00 | this study |
| Sexually-mature female llamas (n = 6) | 2.5 mg/llama (~0.023 mg/kg) im | Estradiol-17β | 0.043 a (±SD = 0.002) | 1 | n.p. | n.p. | 0.32 b (±SD = 0.024) | [27] |
| Female cows (n = 5) | 10 mg/cow (~0.017 mg/kg) c im | Estradiol-17β | 0.087 a (range: 0.057, 0.128) | 1.66 (range: 0.54, 4.96) | n.p. | n.p. | n.p. | [45] |
| 20–35 year old women (n = 10) | 5 mg/woman im | Estradiol-17β | 0.338 a,d (95% CI: 0.208, 0.550) | 3.9 d (95% CI: 2.4, 6.2) | n.p. | n.p. | n.p. | [26] |
| 22–41 year old women (n = 12) | 5 mg/woman e im | Estradiol-17β cypionate | 0.14 (±SD = 0.08) | 0.70 f (±SD = 0.88) | 0.288 f (±SD = 0.168) | 3.74 f (±SD = 3.17) | 0.71 f (±SD = 0.40) | [46] |
| Parameter | Maximum Likelihood Estimate | Relative Standard Error (%) | |
|---|---|---|---|
| Fixed Effects | ka | 0.257 | 10.7 |
| C1 | 2.57 × 104 | 11.1 | |
| VC/F | 1.63 | 2.54 | |
| base_conc | 0.0439 | 18 | |
| Random Effects | ka | 0.251 | 31.9 |
| C1 | 0.229 | 32.8 | |
| base_conc | 0.516 | 25.7 | |
| Error | b | 0.211 | 11.5 |
| Dose of Estradiol Cypionate | Predicted Population Time-to-Reach-Concentration (Days) | ||
|---|---|---|---|
| 10th Percentile | Median | 90th Percentile | |
| Target 0.1 ng/mL (100 pg/mL) | |||
| 0.1 mg/kg | 4.28 | 5.44 | 6.90 |
| 0.3 mg/kg | 7.76 | 9.83 | 12.1 |
| 0.5 mg/kg | 9.40 | 11.8 | 14.9 |
| 0.8 mg/kg | 10.7 | 13.8 | 17.5 |
| Target 0.05 ng/mL (50 pg/mL) | |||
| 0.1 mg/kg | 6.52 | 8.22 | 10.3 |
| 0.3 mg/kg | 9.86 | 12.6 | 15.8 |
| 0.5 mg/kg | 11.4 | 14.6 | 18.5 |
| 0.8 mg/kg | 12.7 | 16.6 | 21.3 |
| Cat ID | Ovary Primary Follicles | Ovary Secondary Follicles | Ovary Early Tertiary Follicles | Ovary Graafian Follicles | Ovary CL | Ovary CA | Endometrial Surface Epithelium a | Endometrial Glands b | Stage of Cycle c |
|---|---|---|---|---|---|---|---|---|---|
| 1 | Numerous | Numerous | Multiple | None | Multiple PP3 | None | S to PS HC with MOD HYE | NLG with B, MIN D and E | Luteal/PP3 |
| 2 | Numerous | Numerous | Multiple | None | Multiple PP3 | None | S to PS HC with MIL HYE | NLG with B, MIN D and E | Luteal/PP3 |
| 3 | Numerous | Numerous | Multiple | None | Multiple PP1 | None | PS HC with MOD HYE | NLG with B, MOD D and E | Luteal/PP1 |
| 4 | Numerous | Numerous | Multiple | None | Multiple PP3/4 | None | S to PS HC with MOD HYE | NLG with B, MIN D and E | Luteal/PP3/4 |
| 5 | Numerous | Numerous | Multiple | None | Multiple PP3 | None | S to PS HC with MOD HYE | NLG with B, MIL D and E | Luteal/PP3 |
| 6 | Numerous | Numerous | Multiple | None | Multiple PP3 | None | S to PS HC with MIL HYE | NLG with B, MIL D and E | Luteal/PP3 |
| 7 | Numerous | Numerous | Multiple | None | Multiple PP2/3 | One | S to PS HC with MIL HYE | NLG with B, MIN D and E | Luteal/PP2/3 |
| 8 | Numerous | Numerous | Multiple | None | Single PP2 | None | S to PS C | FLG with E | Luteal/PP2 |
| 9 | Numerous | Numerous | Multiple | None | Multiple PP3 | None | S to PS HC with mil HYE | NLG with B, mil D and E | Luteal/PP3 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hyndman, T.H.; Algar, K.L.; Woodward, A.P.; Coiacetto, F.; Hampton, J.O.; Nickels, D.; Hamilton, N.; Barnes, A.; Algar, D. Estradiol-17β Pharmacokinetics and Histological Assessment of the Ovaries and Uterine Horns following Intramuscular Administration of Estradiol Cypionate in Feral Cats. Animals 2020, 10, 1708. https://doi.org/10.3390/ani10091708
Hyndman TH, Algar KL, Woodward AP, Coiacetto F, Hampton JO, Nickels D, Hamilton N, Barnes A, Algar D. Estradiol-17β Pharmacokinetics and Histological Assessment of the Ovaries and Uterine Horns following Intramuscular Administration of Estradiol Cypionate in Feral Cats. Animals. 2020; 10(9):1708. https://doi.org/10.3390/ani10091708
Chicago/Turabian StyleHyndman, Timothy H., Kelly L. Algar, Andrew P. Woodward, Flaminia Coiacetto, Jordan O. Hampton, Donald Nickels, Neil Hamilton, Anne Barnes, and David Algar. 2020. "Estradiol-17β Pharmacokinetics and Histological Assessment of the Ovaries and Uterine Horns following Intramuscular Administration of Estradiol Cypionate in Feral Cats" Animals 10, no. 9: 1708. https://doi.org/10.3390/ani10091708