1. Introduction
Soybean meal (SBM), a co-product of soybean oil extraction from soybeans, is commonly used in the poultry and swine industries because of its high quality protein and relatively well-balanced amino acid content [
1]. However, a variety of antinutritional factors (ANFs) in SBM, such as oligosaccharides, lectin, and allergenic antigens, hinder the digestion and absorption of its nutrients by animals [
2,
3,
4]. Glycinin and β-conglycinin are the two main types of allergenic soybean proteins, accounting for about 30% and 40% of the total protein in SBM, respectively [
5]. Previous studies have demonstrated that glycinin damages the intestinal morphology, with effects such as intestinal atrophy and necrosis, and causes malabsorption syndrome in animals [
6,
7,
8]. β-Conglycinin causes hypersensitive immune response and negatively affects the growth performance of animals [
9,
10]. These drawbacks limit the extensive use of SBM in animal feeds. Therefore, the development of processing techniques that remove the harmful and antinutritional factors of SBM may produce a high-quality protein with functional properties [
11].
Fermentation is considered an economically viable processing technique in which microorganisms are used to increase the protein concentration, reduce the content of ANFs, and enhance the nutritional value of SBM [
12]. It has been documented that the apparent ileal digestibility of the dry matter, nitrogen and energy in fermented soybean meal (FSBM) is similar to that of fish meal [
13]. Bacteria and fungi are the two main types of microorganisms used to ferment animal feed or feed ingredients.
Bacillus spp. have a stronger capacity to resist heat and acid [
14], and secrete many kinds of extracellular enzymes, including non-starch polysacchrides degrading enzymes and proteinase, and are therefore commonly used in bacteria-based fermentation [
15]. Seo and Cho reported that the contents of glycinin and β-conglycinin in SBM decreased by 70% and 50%, respectively, after fermentation with
B. subtilis KCCM11438P for 24 h [
16]. Teng et al. reported that fermentation of SBM with
B. subtilis SB102 degraded its large antigenic proteins, and increased the small-size proteins and the contents of some essential amino acids compared with those in unfermented SBM [
17]. In addition to
Bacillus spp.,
Lactobacillus spp. are also frequently used in solid state fermentation to produce organic acids and reduce the pH of SBM. Chi and Cho demonstrated that
Lactobacillus spp. significantly reduced the pH of SBM, but did not decompose protein-based ANFs other than trypsin inhibitors after solid state fermentation for 36 h [
18]. Fungal fermentation with several species of the genus
Aspergillus, such as
A.
niger and
A.
oryzae, caused much smaller increases in small proteins and smaller reductions in larger antigenic proteins in SBM than bacterial fermentation [
15]. The efficiency of fermentation in improving nutritional value of SBM depends on growth rate, extracellular enzymes and its activities of microorganisms involved [
18]. Therefore, it is vital to choose a highly efficient allergen-degrading microorganism for the production of FSBM with high nutritional quality.
The glycinin and β-conglycinin in SBM does not cause hypersensitive immune response in gastrointestinal tract or affect the growth performance of ruminants as they do in pre-ruminant animals and nonruminant animals because the rumen contains a mixed community of microbes [
19] that can inactivate and degrade glycinin and β-conglycinin [
20]. About 12% to 38% of the total bacterial population in rumen are strains with proteolytic activities [
21], so the rumen is an ideal place to select an appropriate bacterium for the decomposition of glycinin and β-conglycinin in SBM. In previous studies, target bacteria were successfully isolated from a preserved vegetable [
22], traditional fermented soy foods [
23], and animal intestinal tracts [
24]. However, to our knowledge, no research has used a strain derived from the rumen flora to ferment SBM. The selected bacteria have previously been used to ferment SBM directly, and animal experiments have been conducted without considering the safety of the strain in these experiments. However, some bacteria may produce hemolysin, which is considered a virulence-related factor that impairs the intestinal epithelial cells [
25,
26]. Therefore, the objective of this study was to screen a highly efficient bacterial strain in a cow rumen with which to degrade the antigenic proteins in SBM, and to improve the nutritional quality of SBM by fermentation with this strain. The safety of this bacterium was evaluated with an acute oral toxicity test.
2. Materials and Methods
2.1. Isolation of Bacterial Strain
Rumen liquid was collected from a fistulated cow feeding at the Experimental Nankou Base of Chinese Academy of Agricultural Science (Beijing, China), and was filtered through double cheesecloth. An aliquot (1 mL) of the rumen liquid was homogenized in 9 mL of 8.5% (
w/
v) sterile salt water, and then incubated at 80 °C for 20 min to destroy other microbial cell [
23,
27]. Defatted milk agar pates were inoculated with this homogenate for bacterial culture by gradient dilution (10
−2–10
−7). These inoculated plates were cultured in a biochemical incubator at 37 °C for 24 h. After incubation, strains were selected according to the size of the transparent zone around colonies (data not shown). Six strains were isolated for a further screening experiment, and denoted ´GYL’. Two strains of
B. subtilis stored previously in our laboratory, designated BSD2 and CICC21237, were used as the positive controls.
2.2. SBM and Medium Preparation
SBM was purchased from the Hopefull Grain & Oil Co. Ltd. (Langfang, Hebei, China). Agar plates containing antigenic protein (glycinin or β-conglycinin), starch, or xylan were used to determine the protease, amylase, and xylanase activities of the strains, respectively. The antigenic proteins were extracted from SBM according to the method reported by Liu et al. [
28] and Deak et al. [
29], with minor modification. Briefly, about 15 g of defatted soy flour was extracted with 0.03 M Tris-HCl (pH 8.5) in a 15:1 (
w/
v) buffer-to-flour ratio. The resulting slurry was stirred 200 r/min for 1 h at 45 °C and centrifuged at 9000×
g for 40 min at 4 °C. After centrifugation, the supernatant was discarded and the precipitate was extracted two more times. The appropriate amount of solid NaHSO
3 was added to the supernatant to achieve a concentration of 10 mM NaHSO
3 and the pH was adjusted to 6.4 with 2 M HCl. The slurry was stored at 4 °C overnight and then centrifuged at 6500×
g for 20 min at 4 °C. The glycinin-rich fraction was thus obtained. Solid NaCl was added to the supernatant to a final concentration of 0.25 M and the pH adjusted to 5.5 with 2 M HCl. The slurry stirred at 45 °C and 200 r/min for 30 min, and then centrifuged at 9000×
g for 40 min at 4 °C. The supernatant was diluted two-fold with pure water and adjusted to pH 4.8 with 2 M HCl. The β-conglycinin-rich fraction was obtained after centrifugation at 6500×
g for 20 min at 4 °C. The total precipitate was dissolved in pure water to prepare protein plates. Starch agar plates were prepared by adding 1 g of corn starch and 1.5 g of agar to 100 mL of Luria-Bertani (LB) medium (1% tryptone, 1% NaCl, and 0.5% yeast extract). Xylan agar plates were prepared by mixing 0.3 g of xylan and 1.5 g of agar to 100 mL of LB. To prepare defatted milk agar plates, 5 g of defatted milk (Yili, Inner Mongolia, China) and 1.5 g of agar were added to 100 mL of LB. All plates were autoclaved for 21min at 121 °C.
2.3. Solid State Fermentation
The SBM for fermentation was sieved through a 10 mesh screen. The inocula of eight bacteria were prepared by incubating a single colony in LB medium at 37 °C for 21 h. SBM (20 g) was mixed with distilled water to 50% (w/v) in a 150 mL conical flask and inoculated with fresh inoculum in a 5% ratio (w/v). Each mixture was incubated at 37 °C for 24 h, each treatment was performed in triplicate. After fermentation, the FSBM samples were dried at 60 °C for 4 h, ground, and passed through a 60-mesh screen. The fermented powder samples were stored at 4 °C before analysis.
2.4. Chemical Composition of FSBM
The crude protein content was estimated with a combustion analyzer (Dumatherm, Gerhardt, Germany) using ethylenediaminetetraacetic acid as the calibration standard. The content of trichloroacetic acid-soluble protein (TCA-SP) in SBM or FSBM was measured with trichloroacetic acid (TCA) according to the China National Standard (GB/T 22492-2008). Briefly, 1 g of sample was mixed with 20 mL of 15% (w/v) TCA, and the slurry was stirred at room temperature for 5 min and centrifuged at 4000× g for 10 min. The supernatant was collected, and the nitrogen content of the supernatant was determined with an automatic Kjeldahl Nitrogen Analyzer (FOSS, Hillerod, Denmark).
The contents of glycinin and β-conglycinin in the SBM or FSBM were analyzed using competitive enzyme-linked immunosorbent assay (ELISA) kits (Longkefangzhou Bio-Engineering Technology Company, Beijing, China). The total amino acid content was extracted with 6 M HCl and measured with an automatic amino acid analyzer (Hitachi L-8900, Tokyo, Japan).
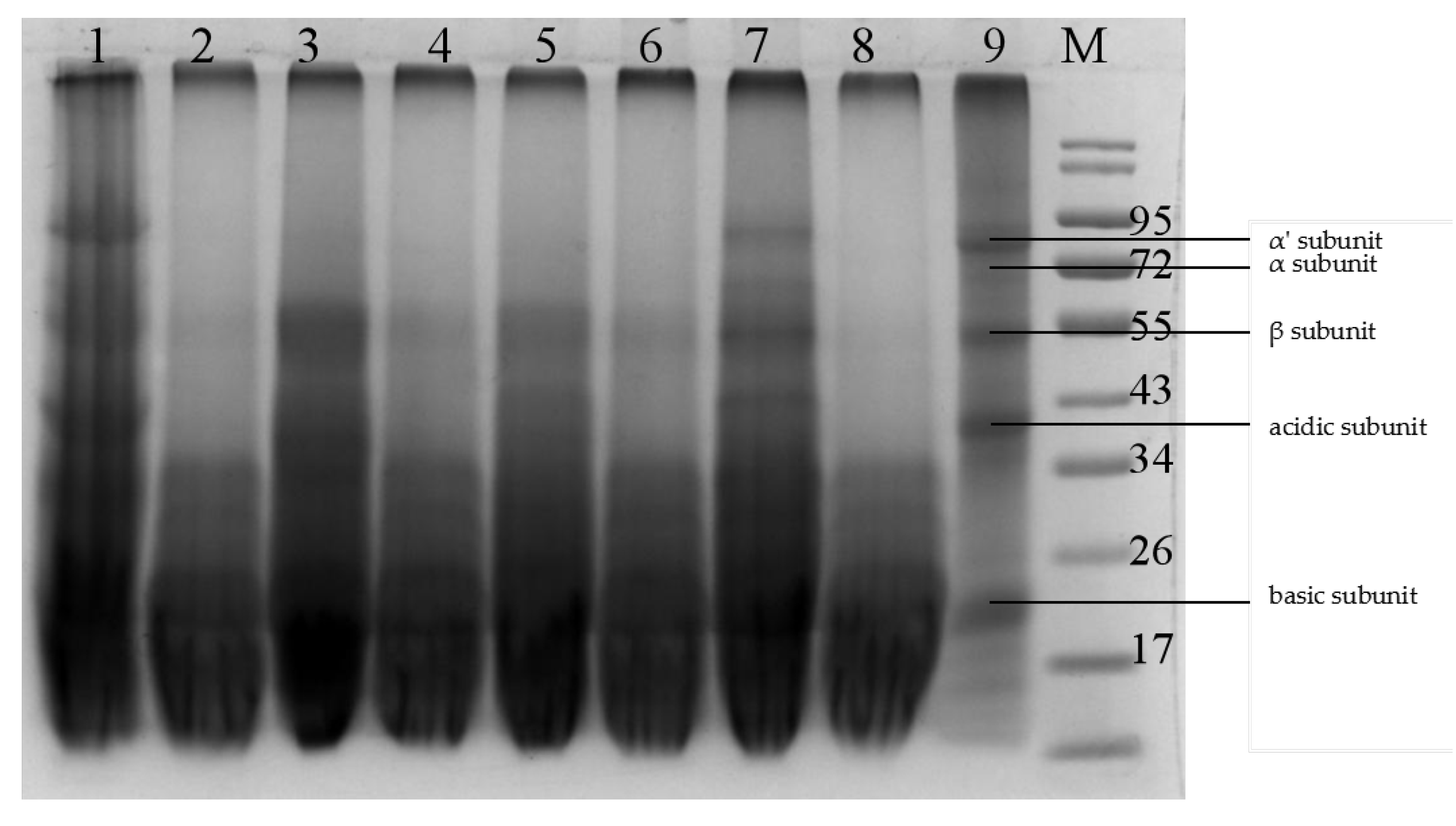
2.5. Protein Extract and SDS-PAGE
The soluble proteins in SBM and FSBM were extracted as previously described by Yang et al. [
1]. The soluble protein concentrations were quantified with Bio-Rad Protein Assay Kit (Bio-Rad, Hercules, CA, USA), according to the manufacturer’s protocol. The fractions of soluble protein in the SBM and FSBM were detected with 12% polyacrylamide separating gels containing 0.1% SDS in Tris-glycine buffer. About 15 μL of total denatured soluble protein sample was loaded into a well. The extracted soluble protein sample was concentrated at 80 V for 30 min and separated at 120 V for 60 min. The gel was stained with Coomassie Brilliant Blue R-250 (Bio-Rad) for 45 min, and destained with 7% (
v/
v) acetic acid.
2.6. Biochemical Characteristics and Molecular Identification of the Candidate Strain
The biochemical characteristics of the strain GYL6 were determined using a commercial kit (Hopebio Co., Ltd., Qingdao, China) according to the manufacturer’s instructions. The genomic DNA of strain GYL6 was extracted with the Ezup Genomic DNA Extraction Kit (Sangon Biotech Co., Ltd., Shanghai, China), according to the manufacturer’s protocol. The full-length sequence of 16S rRNA of strain GYL6 was amplified by polymerase chain reaction (PCR) with the primers 27F (5′-AGAGTTTGATCMTGGCTCAG-3′) and 1492R (5′-TAGGG TTACCTTACGACTT-3′) (Sangon Biotech Co., Ltd.). The gyrase B (gyrB) gene of strain GYL6 was amplified with PCR using primers UP1F (5′-GAAGTCATCATGACCGTTCTGCA-3′) and UP2R (5′-AGCAGGGTACGGATGTGC GAGCC-3′) (Sangon Biotech Co., Ltd.). The PCR amplification was carried out in a 25 μL reaction mixture containing 25 ng of template DNA, 0.5 μL of each primer, 1 μL PCR premix, 0.2 μL Taq mix, 2.5 μL buffer and PCR-grade water to adjust the volume. The PCR reaction was conducted under the following conditions, 5 min at 94 °C, followed by 35 cycles at 94 °C for 30 s, 54 °C for 30 s, and 72 °C for 90 s, with final extension for 10 min at 72 °C. Amplified PCR products were analyzed by agarose gel (1%) electrophoresis. After gel extraction, the products were purified by SanPrep pillar PCR purification kit (Sangon Biotech Co., Ltd.). Finally, the PCR products of strain GYL6 were sequenced on Illumina Miseq platform (Illumina Inc., San Diego, CA, USA) provided by Sangon Biotech Co., Ltd. The ribosomal sequences obtained were compared with those released in GenBank (National Center for Biotechnology Information, Bethesda, MD, USA) database using the Nucleotide Basic Local Alignment Search Tool program.
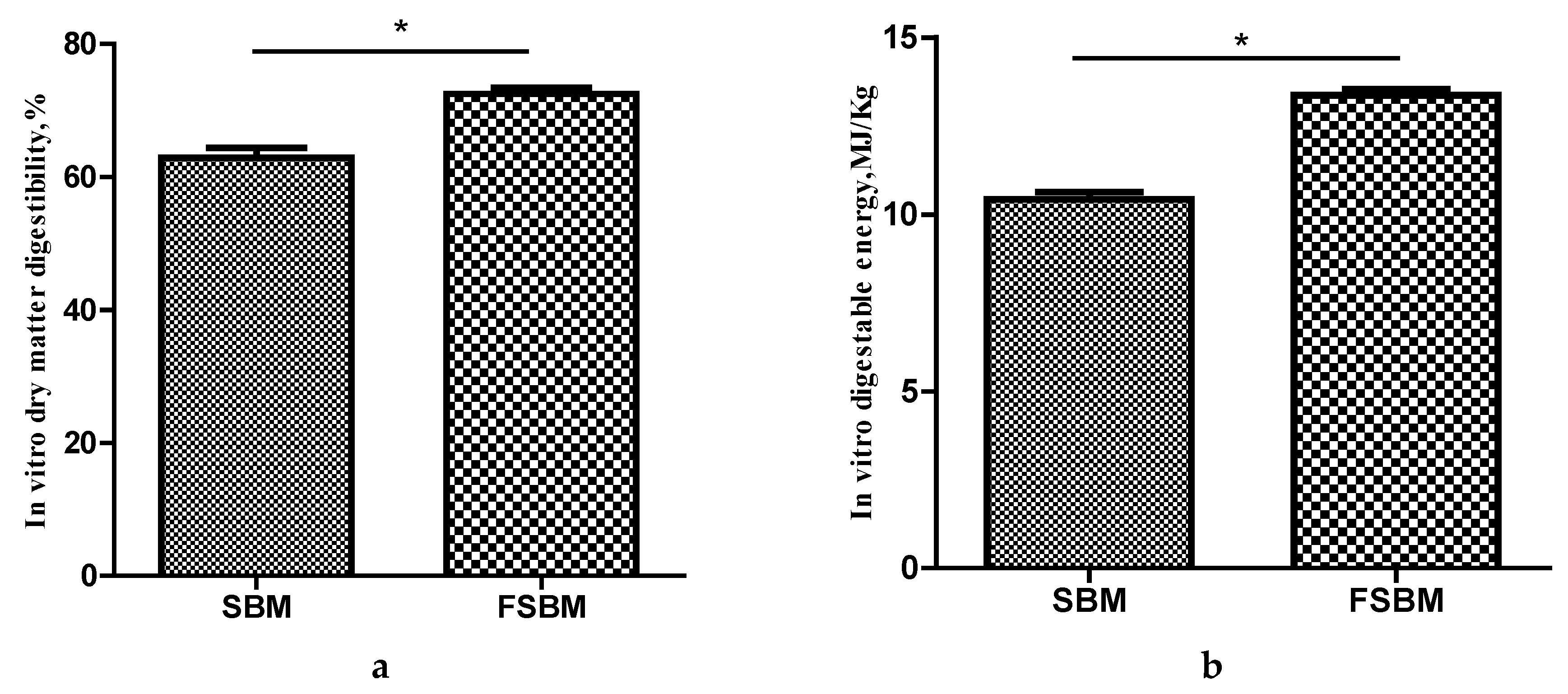
2.7. In vitro Digestibility
The in vitro digestibility of dry matter and digestible energy of FSBM and SBM were assayed with a computer-controlled simulated digestion system (CCSDS), which was developed and designed by Zhao et al. and Chen et al. [
30,
31] to simulate the gastrointestinal digestion system of monogastric animals. The machine automatically finishes the in vitro digestion process, including the gastric, upper- and lower-intestinal digestion, when the procedural instructions are followed. In brief, 2 g of FSBM or SBM and 20 mL simulated gastric fluid (1500 U/mL pepsin, pH 2.0) was placed in the dialysis tubing within the digestion chamber, and then the gastric buffer solution (pH 2.0, 41 °C) was pumped (120 mL/min) into the digestion chambers and circulated for 4 h. After gastric digestion, the upper-intestinal digestion was performed, 2 mL of concentrated simulated small-intestinal fluid (401.46 U/mL amylase, 49.28 U/mL trypsin, 11.31 U/mL chymotrypsin, pH 6.50) was injected into the digestion chamber, followed by injection of the upper-small-intestinal buffer solution (120 mL/min; pH 6.50, 41 °C) and circulation for 7.5 h. Finally, the lower-small-intestinal buffer solution (pH 8.0, 41 °C) was pumped into the digestion chambers and circulated for 7.5 h. During in vitro digestion, a temperature of 41 °C was maintained in a shaking incubator set to 180 rpm/min, to allow the sample and digestive fluid to mix adequately. After the simulated digestion was completed, the undigested residues were transferred to a preweighed vessel and dried overnight at 65 °C and then at 105 °C for 5 h until a constant weight was achieved. The vitro digestibility of dry matter and the digestible energy were calculated with the following equations:
2.8. Safety Assessment of the Candidate Strain
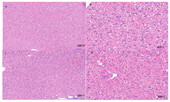
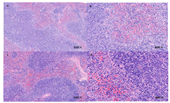
In the preliminary experiment, the candidate strain showed no harmful effect on the health of mice, so an acute oral toxicity test was performed with a maximal tolerated dose. Twenty healthy specific-pathogen-free (SPF) grade male C57BL/6J mice (7–8 weeks old) were obtained from Sibeifu (Beijing, China) Biotechnology Co., Ltd. (Beijing, China), certificate of conformity no. SCXK (Jing) 2016–0002). The mice were randomly divided into two groups based on body weight and housed five per cage. The room for mouse growth was maintained at a temperature of 23 ± 2 °C, a relative humidity of 50–55%, and a 12 h light-dark cycle. The mice were fed ad libitum and had free access to drinking water. After acclimation for 7 days, the mice in the treatment group were administered 0.4 mL of bacterial suspension (1.0 × 109 CFU/mL) by oral gavage daily for 7 consecutive days, which was approximately equivalent to 2.0 × 1010 CFU per kg bodyweight, and the control group of mice was administered the same amount of physiological saline. After gavage, clinical observations were carried out and recorded daily, including any changes in skin, eyes, or fur, the occurrence of secretions or excretions, and autonomic activity, such as lacrimation, piloerection, and unusual respiratory patterns. Two weeks after gavage, the mice were killed by cervical dislocation. Their spleens, livers, and intestinal tracts were subjected to necropsy and histopathological examination. The spleens, livers, and hearts were also immediately weighed and the organ ratios calculated. The present study was approved by the Animal Ethics Committee of Chinese Academy of Agricultural Science (code number: AEC-CAAS-20190721; approval date: 21 July 2019) and was conducted in accordance with the principles outlined in the National Institutes of Health (NIH) Guide for the Care and Use of Laboratory Animals.
2.9. Statistical Analysis
All statistical analyses were performed with the SPSS 19.0 software (SPSS Inc., Chicago, IL, USA). The data on crude protein, TCA-SP, and antigenic proteins were analyzed with one-way ANOVA and Duncan’s post-hoc test. The comparison of two means was tested using Student’s t-test. Difference were considered significant at p < 0.05.
4. Discussion
Fermented feed has the advantages of improving the feed quality and gut ecology of animals compared with supplementation with feed with additives, such as probiotics, prebiotics, synbiotics, and enzymes [
34]. The kind of microorganisms used in this fermentation is one of the major factors affecting the nutritional value of feed and feed ingredients.
Bacillus ssp. are the predominant fermentative microorganisms used to ferment feed. In previous studies,
Bacillus ssp. were used to ferment SBM from traditional fermented soy food [
22,
23], and aquatic animals, such as shrimp guts [
35] and grass carp intestine [
24], respectively. They were screened with soymilk, xylan, and starch agar plate methods. However, the ability of the target strains to degrade glycinin and β-conglycinin could not be determined indirectly with these agar plate tests. In the present study, glycinin and β-conglycinin were extracted from SBM and used as the sole substrates upon which to culture the candidate strains. The abilities of the candidate strains to degrade the antigenic proteins were compared directly by measuring the diameters of the transparent zones on the culture plates. Among these eight isolates, strain GYL6 showed higher proteinase, xylanase, and amylase activities than other seven strains on agar plates. The results of solid state fermentation with these eight strains confirmed that strain GYL6 was the most efficient strain. It eliminated most glycinin and β-conglycinin from the SBM, and significantly increased its content of crude protein and TCA-SP. These results were consistent with the results of agar plate test method. These findings show that plate tests are an effective and convenient method to screen suitable strains in future studies.
It is recommended that gyr B gene sequences be used, as well as 16S rRNA gene sequences, to identify bacterial strains, because the high genetic similarity of
Bacillus spp. make them difficult to discriminate. A previous study demonstrated that the gyrB gene has a faster rate of molecular evolution than 16S rRNA and more precisely identifies bacterial strains [
36]. In the current study, strain GYL6 was identified as
B. amyloliquefaciens using both 16S rRNA and gyrB sequences.
This study clearly demonstrates that 92.32% of glycinin and 85.05% of β-conglycinin were removed after fermentation with
B. amyloliquefaciens for 24 h. These results are similar to those reported by Zhang et al., who found that 84.77% of glycinin and 87.07% of β-conglycinin were degraded after fermentation with
B. subtilis BS12 for 24 h [
22]. By contrast, only 42% of glycinin and 70% of β-conglycinin were eliminated by another strain of
B. subtilis [
16]. Under optimum fermentation conditions (temperature 40 °C, time 48 h and 4% inoculum (
w/
v)), the glycinin in SBM fermented with
B.
sanfensis or
B.
stratosphericus decreased by only 62.0% or 41.1%, respectively [
35]. These differences in the degradation of the antigenic proteins in SBM indicates that the
B.
amyloliquefaciens isolated in the present study has a stronger capacity for protein hydrolysis. Recently, we found that
B.
amyloliquefaciens secretes many extracellular hydrolytic enzymes during its fermentation of SBM, including not only serine protease and acid protease, but also endoglucanase, acetylxylan esterase, and catalase, which broke down cell-wall polymers and facilitated the degradation of antigenic protein (unpublished data). The reduction of antigenic soybean proteins increases the digestibility of nutrients and improves the growth performance of animals [
9,
37]. Previous studies have demonstrated that FSM enhances intestinal barrier function [
38,
39], attenuates the intestinal mucosal immune response [
9], and reduces the incidence of diarrhea, which are related to the reduction in the antigenic proteins of SBM.
Several reports had demonstrated that fermentation with different
Bacillus spp. substantially increases the crude protein content of SBM [
17,
18,
40]. In the present study, crude protein increased significantly by 17.54% after fermentation with
B. amyloliquefaciens for 24 h. The increase in crude protein may be attributable to the loss of carbohydrates, which might be used as substrates in the growth and reproduction of bacteria. At the same time, the content of TCA-SP, which consists of small peptides with 2–20 residues, was approximately 11-fold higher in FSBM than in SBM. A previous study demonstrated that dipeptides and tripeptides are directly absorbed by the animal intestinal tract, and that amino acids in the form of small peptides are transported more rapidly than their constituent amino acids in free form [
22]. Our analysis of the in vitro digestibility suggested that the fermentation of SBM with
B. amyloliquefaciens enhanced its nutritional value and it was digested easily by digestive enzymes.
In present study, SBM fermented with
B. amyloliquefaciens significantly increased the total amino acids by 6.57% and changed its amino acids composition. The essential amino acids, including valine, methionine, phenylalanine, and histidine, were 10–17% higher in FSBM than in SBM. These results are partly consistent with the findings of Medeiros et al., who reported that the contents of lysine, valine, histidine, and phenylalanine increased by 15–19% in FSBM fermented with
B. amyloliquefaciens. Among these essential amino acids, methionine, the first limiting amino acid in the corn-soybean meal diet, significantly influences the energy metabolism, immune function [
41] and growth performance of animals [
42]. Valine is a functionally essential branched-chain amino acid and the fourth limiting amino acid, which also plays an important role in improving growth performance and enhancing the immunity of animals [
43]. Therefore, these increases in some essential amino acids could reduce the currently necessary supplementation of animal feed with synthetic amino acids. However, the amino acid composition of SBM is altered differently when it was fermented with different microorganisms. In the present study, we found that the lysine content remained unchanged, whereas methionine increased in FSBM. These results are identical to those reported by Chi and Cho [
18], but contrary to findings of Gao et al. [
35], who demonstrated that the content of methionine was more than two fold higher in FSBM than in SBM. Hassaana et al. reported that except for serine and histidine, all the amino acids in SBM increased significantly after fermentation with
Saccharomyces cerevisiae [
44], whereas in another study, the essential amino acid profile of SBM was unchanged after fermentation with
A. oryzae GB-107 [
45]. The discrepancies in these finding may be attributable to the proteinase profiles and secretion abilities of the microorganism used [
15]. These findings also indicate that the suitable microorganism for fermentation must be selected in accordance with the nutritional requirements of final fermented feed product.
Today, many researchers and companies use strains from different sources to directly ferment feed, without considering the potential risks of the isolated strains. Previous studies have reported that some
Bacillus spp., including
B. cereus,
B. coagulans, and
B. weihenstephanesis, produce both enterotoxins and emetic toxins, which are recognized as the main causes of food poisoning and other non-gastrointestinal infections [
46]. Therefore, it is important to comprehensively evaluate the safety of a bacterial strain before it is incorporated into animal feed [
26].In vitro and in vivo method are the main approach to evaluate the safety of probiotics. In vivo method includes detecting the known toxin gene, using cultured cell line to evaluate adhesion, invasion, and cytotoxicity of probiotics [
47,
48]. In vivo studies are the preferred approach to substantiate in vitro analysis of safety [
49]. However, no specific guidelines are currently in place for assessing probiotics, but acute oral toxicity testing is the fundamental assay for assessing the safety of novel microbial strains intended as feed additives or for use in feed production, and has been applied in many studies [
14,
50]. For an acute toxicity study, mice were chosen because they are considered the most sensitive laboratory animals [
47]. Based on the results of our preliminary experiment, acute oral toxicity testing was performed by intragastrically administering
B. amyloliquefaciens to mice at a maximal dose of 2.0 × 10
10 CFU/kg BW for 7 consecutive days in this study. At the end of this experiment, all the mice were healthy, with no noticeable abnormal behavior or changes in activities. There were no significant changes in the relative weight of the major organs of mice. Relative organ weight has long been recognized as an important indicator of chemical organ damage [
46,
51]. Hyperplastic changes, congestion, edema, and visceral atrophy also reflect the damage caused by poisonous substances [
51]. In the present study, there were no differences in the relative organ weights or morphological structures of the tissues after the administration of
B. amyloliquefaciens, indicating that the strain had no adverse effects on the health status of mice and can be considered safe for fermented feed production.






{kind=link}
{kind=link}
{kind=link}
{kind=link}